










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkRevista Chapingo. Serie horticultura
On-line version ISSN 2007-4034Print version ISSN 1027-152X
Rev. Chapingo Ser.Hortic vol.14 n.1 Chapingo Jan./Apr. 2008
Parámetros genéticos en la variedad CHF1 Chapingo de tomate de cáscara (Physalis ixocarpa Brot.)
Genetic parameters of the husk tomato ( Physalis ixocarpa Brot.) CHF1 cultivar
A. Peña–Lomelí1*, J. D. Molina–Galán2, J. Sahagún–Castellanos1, J. Ortiz–Cereceres2, F. Márquez–Sánchez1, T. Cervantes–Santana2 y J. F. Santiaguillo–Hernández1
1 Departamento de Fitotecnia, Universidad Autónoma Chapingo. Km. 38.5 Carretera México–Texcoco. Chapingo, Estado de México, C. P. 56230. México. Correo–e: aplomeli@correo.chapingo.mx (*Autor responsable).
2 Programa de Genética, IREGEP, Colegio de Postgraduados. Km. 36.5 Carretera México–Texcoco. Montecillo, Estado de México, C. P. 56230. México.
Recibido: 8 de noviembre, 2006
Aceptado: 14 de agosto, 2007
Resumen
El objetivo de esta investigación fue estimar los parámetros genéticos de la variedad CHF1–Chapingo de tomate de cáscara (Physalis ixocarpa Brot.). Se evaluaron 200 familias de medios hermanos maternos en tres ambientes, con un diseño experimental de bloques al azar con dos repeticiones y 22 plantas por unidad experimental. La heredabilidad en sentido estricto de los 13 caracteres estudiados fluctuó entre 37.1 y 57.7% y el coeficiente de variación genético aditivo entre 8.2 y 24.4%; para rendimiento total estos valores fueron 37.5 y 10.4%, respectivamente. El número y peso de frutos en el primer corte, y frutos totales por planta fueron los principales componentes del rendimiento total. Se encontraron correlaciones genéticas aditivas con valores absolutos altos del rendimiento total con: rendimiento en el primer corte (0.67), frutos por planta en éste (0.50), frutos totales por planta (0.58), número de frutos amarrados (0.38) y altura a la primera bifurcación del tallo (–0.49).
Palabras clave: tomatillo, varianza aditiva, heredabilidad, correlaciones genéticas.
Abstract
The objective of this research was to estimate the genetic parameters of the husk tomato (Physalis ixocarpa Brot.) CHF1–Chapingo cultivar. Two hundred maternal half sib families were evaluated under three environments, in a complete randomized block design with two replications, and 22 plants by experimental plot. The narrow sense heritability of the 13 studied traits varied between 37.1 and 57.8%; the genetic additive variation coefficient presented values between 8.2 and 24.4%; for the total yield, values were 37.5 and 10.4%. The fruit number and weight at the first harvest, and the total plant fruit number were the most important yield components. High additive genetic correlation absolute values were found for the total yield with: the first harvest yield (0.67), fruits per plant in the first harvest (0.50), total fruits per plant (0.58), fruit set number (0.38), and first stem bifurcation height (–0.49).
Key words: tomatillo, additive variance, heritability, genetics correlations, selection.
INTRODUCCIÓN
El tomate de cáscara (Physalis ixocarpa Brot.) –también llamado tomate, tomate verde, de hoja, de fresadilla, de bolsa y tomatillo– era conocido por los Mayas y Aztecas desde épocas prehispánicas y México es su centro de origen y domesticación (Peña y Márquez, 1990; Santiaguillo et al., 1994). Entre las hortalizas, el tomate de cáscara ocupa el cuarto lugar en superficie cultivada en México (SIACON–SAGARPA, 2005), donde existe gran variabilidad genética que ha sido agrupada en ocho razas: Silvestre, Milpero, Arandas, Tamazula, Manzano, Rendidora, Salamanca y Puebla (Ayala et al., 1992; Peña et al., 1992), distribuidas prácticamente en todo el país en altitudes que van desde los 8 hasta los 3,350 m (Santiaguillo et al., 1994). Las razas más importantes desde el punto de vista comercial son Rendidora, Salamanca y Tamazula (Peña y Márquez, 1990).
En una especie como el tomate de cáscara (alógama obligada por ser autoincompatible) donde no se ha realizado mejoramiento genético en gran escala, la selección masal es en primera instancia un buen método para obtener avances significativos (Pérez et al., 1996). Sin embargo, la selección recurrente exitosa debe cambiar la estructura de la variabilidad genética de la población, por lo que es pertinente estimar periódicamente los parámetros genéticos de las poblaciones bajo mejoramiento con el fin de, en su caso, diseñar estrategias alternativas para mantener o aumentar el avance genético (Hallauer y Miranda, 1981). En este contexto, el objetivo de la presente investigación fue estimar la varianza genética aditiva y la heredabilidad de diferentes caracteres y las correlaciones genéticas entre éstos en la variedad CHF1 Chapingo, obtenida mediante seis ciclos de selección a partir de la variedad Rendidora (Peña, 2005).
La varianza genética aditiva es la determinante principal de las propiedades genéticas observables en una población (heredabilidad y correlación genética aditiva) y de la respuesta a la selección (Falconer, 1986). La estimación de los componentes de varianza genética se puede realizar mediante los diseños de apareamiento de Carolina del Norte (Comstock y Robinson, 1952), el estudio de generaciones descendientes del cruzamiento entre dos líneas y sus retrocruzas (Mather y Jinks, 1971) y los diseños dialélicos (Christie y Shattuck, 1992). La heredabilidad es una propiedad de cada carácter y población, y la utilidad de su estimación radica en su sentido predictivo de la respuesta a la selección (Nyquist, 1991). La correlación genética aditiva, cuya causa principal es la pleiotropía, indica el grado en que dos caracteres están influidos por los mismos genes o por genes estrechamente ligados que determinan caracteres diferentes; es importante en el contexto de la selección, ya que explica el cambio de un carácter en función del cambio de otro, lo que permite hacer predicciones al respecto e incluso selección indirecta (Falconer, 1986).
Con relación a los parámetros genéticos de las poblaciones, se han hecho estimaciones de componentes de varianza genética, heredabilidad y correlaciones genéticas en muchas especies; sin embargo, en tomate de cáscara, objeto de estudio del presente trabajo, se cuenta con pocas investigaciones relacionadas con el tema (Pérez–Grajales et al., 1996; Moreno et al., 2002; Peña et al., 2004). Esta situación justifica realizar trabajos enfocados al conocimiento de los parámetros genéticos de poblaciones importantes, ya que además de aportar información básica sobre la especie, serán el punto de partida para la definición de mejores estrategias de mejoramiento genético.
MATERIALES Y MÉTODOS
La población estudiada fue la variedad CHF1–Chapingo de tomate de cáscara, obtenida de la variedad Rendidora mediante cinco ciclos de selección masal y uno de selección familial de medios hermanos maternos, primera registrada en México (Peña, 2005); ésta fue establecida por trasplante en Chapingo, Estado de México, en condiciones de riego durante el ciclo primavera–verano de 1994, a partir de ésta fueron derivadas, mediante polinización libre, 200 familias de medios hermanos maternos (FMHM) objeto de estudio de la presente investigación. La evaluación de familias se realizó en condiciones de riego en el ciclo primavera–verano en tres ambientes: Chapingo 1995 (CH95), Tecamac 1995 (TE95) y Chapingo 1996 (CH96), con un diseño experimental de bloques al azar con dos repeticiones. La unidad experimental estuvo constituida por 22 plantas distanciadas a 0.3 m en un surco de un metro de ancho. Los experimentos se establecieron por trasplante (en las fechas 06/04/95, 11/ 04/95 y 28/03/96, para CH95, TE95 y CH96, respectivamente).
Los caracteres estudiados fueron: altura a la primera bifurcación del tallo (APB) en cm; altura total de planta (ATP) en cm; longitud (cm) de la rama principal (LRP), medida desde la primera bifurcación hasta el fin de la rama; número de frutos amarrados (NFA), considerado como el número de flores que desarrollaron el cáliz y tiraron la corola. Estos cuatro caracteres fueron medidos en 20 plantas por unidad experimental al momento de realizar la segunda escarda, 40 días después del trasplante, y están expresados en promedio por planta. En cada uno de los tres experimentos se realizaron dos cortes: el primero a los 71, 73 y 76 días después del trasplante y el segundo a los 18, 20 y 21 después del primero, respectivamente para CH95, TE95 y CH96. Los caracteres cuantificados en cada cosecha fueron: peso promedio (g) de fruto en los cortes uno (PF1) y dos (PF2) y promedio de ambos cortes (PFP), las mediciones se obtuvieron de una muestra de un kilogramo de fruto por unidad experimental; rendimiento de fruto en los cortes uno (RC1) y dos (RC2) y total por planta (RTP), expresados en gramos por planta; y número de frutos por planta en los cortes uno (FP1) y dos (FP2) y totales por planta (FTP), cuyo cálculo se hizo dividiendo el rendimiento por planta entre el peso promedio por fruto correspondiente.
Para la estimación de los componentes de varianza genética se utilizó la estructura de FMHM, que según Márquez–Sánchez y Sahagún–Castellanos (1994) es más precisa en las estimaciones obtenidas que el diseño I de Carolina del Norte. Sin embargo, sólo fue posible estimar la varianza genética aditiva ( σ2A ) por no contar con material genéticamente uniforme (una línea, un híbrido o un clon) que introducido en las evaluaciones permitiera estimar la varianza de dominancia. El análisis de varianza correspondiente al modelo aleatorio que representa la evaluación de f familias (F) en b ambientes (B) y r repeticiones anidadas en ambiente (R/B) para datos promedio por parcela, se presenta en el Cuadro 1 (Knapp et al., 1985; Nyquist, 1991; Molina, 1992; Sahagún, 1993).
Las estimaciones se realizaron suponiendo herencia diploide, dos alelos por locus, equilibrio Hardy–Weinberg de la población, equilibrio de ligamiento y ausencia de epistasis. Los parámetros genéticos se estimaron con base en la estructura del Cuadro 1, del modo siguiente: a) Varianza aditiva, ; b) coeficiente de variación genética aditiva CVA = [σA/
; b) coeficiente de variación genética aditiva CVA = [σA/ ]x100, donde σA es la desviación estándar aditiva y es la media fenotípica del carácter; c) heredabilidad en sentido estricto,
]x100, donde σA es la desviación estándar aditiva y es la media fenotípica del carácter; c) heredabilidad en sentido estricto, ; d) 95% intervalo de confianza para h2 (ICh2), con límite
; d) 95% intervalo de confianza para h2 (ICh2), con límite
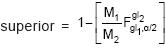 , donde F es el valor de la distribución de F, P≤0.05 y gl1 y gl2 los grados de libertad de familias y de la interacción de BxF, respectivamente. Se calculó también la amplitud del ICh2 (AMIC = límite superior menos límite inferior del intervalo) y la tasa de la heredabilidad (TASA = AMIC /
, donde F es el valor de la distribución de F, P≤0.05 y gl1 y gl2 los grados de libertad de familias y de la interacción de BxF, respectivamente. Se calculó también la amplitud del ICh2 (AMIC = límite superior menos límite inferior del intervalo) y la tasa de la heredabilidad (TASA = AMIC /  ).
).
Con base en la estructura del análisis de varianza (Cuadro 1) se calcularon los componentes de covarianza pares de caracteres. Sean X y Y caracteres cualquiera, la covarianza entre ambos es de la forma: COVF = (PCM1–PCM2)/ar; en tanto que la covarianza aditiva es: COVAxy = 4 COVF. Con esta covarianza se puede estimar la correlación genética aditiva [rGA = COVAxy/(σAxσAy) ] entre cada dos de los caracteres evaluados (Falconer, 1986).
RESULTADOS Y DISCUSIÓN
Medias, coeficientes de variación y parámetros genéticos
El peso por fruto en el corte uno (PF1) fue mayor que en el corte dos (PF2), ocurriendo lo mismo con el número de frutos (FP1 > FP2) y con el rendimiento (RC1 > RC2); el rendimiento total por planta fue de 1,296 g (Cuadro 2), que a la densidad de población utilizada (33,333 plantas·ha–1) significa un rendimiento de 43.2 t·ha–1. La mayor importancia del primer corte en el rendimiento puede deberse a que este momento coincide con el mayor desarrollo del área foliar, en tanto que en el segundo corte se presentan decrementos en ésta por una mayor tasa de producción de etileno (Mulato et al., 1987; Cartujano et al., 1987; Cantwell et al., 1992).
Los coeficientes de variación en general fueron aceptables (Cuadro 2) e incluso se podrían considerar como bajos en comparación con otros estudios sobre evaluación de materiales de tomate de cáscara (López et al., 1994). El más bajo (12.1%) correspondió al carácter longitud de la rama principal y el más alto (34.6) al rendimiento en el corte dos.
De los 13 caracteres estudiados, los estimadores de σ2A (Cuadro 2) para número de frutos por planta en el corte uno (FP1 = 24.6) y rendimiento en el corte uno (RC1 = 19 959.7) son los únicos comparables con datos preexistentes en variedades de la raza Rendidora. Pérez–Grajales et al. (1996) sólo reportan estos dos caracteres, con varianzas aditivas de 23.1 y 22.038 en el material SBI200–93 y de 67.8 y 41.934 para la variedad Rendidora. La diferencia entre la variedad Rendidora y la CHF1 Chapingo es el doble de la  en la primera, lo cual indica que el proceso de mejoramiento redujo este parámetro, esperado (Falconer, 1986) cuando se supone un modelo aditivo y frecuencias génicas de los alelos favorables de al menos 1/2. Los valores de en número de frutos y rendimiento en el corte uno son similares para CHF1–Chapingo y SBI200–93, debido a que ambos materiales han sido derivados a partir de Rendidora y sólo difieren en que en la segunda variedad y el último ciclo de selección se realizó al utilizar como criterio de selección la concentración de frutos y la precocidad de cosecha (Pérez Grajales et al., 1994), en tanto que en CHF1 Chapingo, el criterio de selección fue rendimiento.
en la primera, lo cual indica que el proceso de mejoramiento redujo este parámetro, esperado (Falconer, 1986) cuando se supone un modelo aditivo y frecuencias génicas de los alelos favorables de al menos 1/2. Los valores de en número de frutos y rendimiento en el corte uno son similares para CHF1–Chapingo y SBI200–93, debido a que ambos materiales han sido derivados a partir de Rendidora y sólo difieren en que en la segunda variedad y el último ciclo de selección se realizó al utilizar como criterio de selección la concentración de frutos y la precocidad de cosecha (Pérez Grajales et al., 1994), en tanto que en CHF1 Chapingo, el criterio de selección fue rendimiento.
Si bien las estimaciones de varianza aditiva tienen sentido cuando son comparadas con las de otras poblaciones, el uso de sus valores absolutos puede inducir a interpretaciones erróneas; en todo caso, es preferible comparar sus coeficientes de variabilidad genética aditiva (CVA), sobre todo cuando se desea comparar las varianzas de diferentes caracteres (Molina, 1992). Los CVA obtenidos en el presente trabajo (Cuadro 2) indican que existe menor variabilidad genética aditiva en ATP, LRP y RTP que en NFA, FP1, FP2 y RC2, lo cual significa que es más fácil obtener avances por selección en el segundo grupo de caracteres, ya que en el primero la propia selección ha reducido la varianza aditiva y el CVA, ya que la altura de planta y longitud de rama están asociadas al hábito de crecimiento semirrecto, carácter utilizado como primer criterio de selección, lo que ha conducido a eliminar plantas erectas de ramas largas que se desgajan en la etapa de producción plena. Por otra parte, el CVA fue menor en RC1 que en RC2, debido posiblemente a que la selección masal practicada en la población se ha realizado con base en el primer corte solamente, en donde se considera el número y peso total de frutos, situación que ha propiciado la eliminación de las plantas tardías al primer corte y ha dejado de considerar la variabilidad contenida en ellas. En los caracteres número de frutos por planta (FP1, FP2 y FTP) y peso por fruto (PF1, PF2 y PFP), con menor CVA en FTP y PFP, la explicación puede estar en el hecho de que se han seleccionado frutos compactos y numerosos en un corte total. Con relación a la selección, en poblaciones de maíz con valores de CVA menores de 12% no se ha logrado una alta respuesta a la selección masal, aunque ésta ha reducido el CVA (Estrada, 1977; Vargas et al., 1982; Sahagún et al., 1991), lo anterior sugiere que para el rendimiento total de tomate de cáscara (CVA = 10.4%) es más adecuado realizar selección con base en familias de medios hermanos o de hermanos completos, con las cuales se tendría mejor control ambiental y mayor posibilidad de explotar más intensamente la varianza aditiva (Márquez, 1985).
La estimación de heredabilidad de los 13 caracteres estudiados resultó alta, entre 37.1 y 57.8%, los menores valores ocurrieron en RC2, RTP y PF2 y los mayores en FP1, NFA y APB. Esto indica que aún es posible obtener avances significativos en el mejoramiento por selección y derivar mejores variedades a partir de la CHF1–Chapingo. Sin embargo, es pertinente comentar que tanto σ2A como h2 pudieran estar un poco sobrestimadas, ya que la evaluación de familias de medios hermanos obtenidas de un lote de polinización libre sobrestima la σF2 por contener alguna proporción de hermanos completos (Nyquist, 1991).
Por otro lado, la amplitud de los intervalos de confianza de las heredabilidades en general fue pequeña (Cuadro 2), se obtuvo la menor en el carácter FP1 y la mayor en RC2, mismos que presentaron la menor y mayor tasa de la heredabilidad, respectivamente. Lo anterior significa que la precisión en las estimaciones fue buena (Knapp et al., 1987); en el mejor de los casos, FP1, donde la estimación puntual de h2 fue 57.8%, el valor verdadero de este parámetro se encuentra entre 46.5 y 67.0% (P≤0.05) y para el peor, RC2, la estimación puntual de h2 fue 37.1%, el valor verdadero estará entre 20.4 y 50.9% (P≤0.05). La heredabilidad reportada por Pérez–Grajales et al. (1996) en el material SBI200–93 del número y peso de frutos por planta en el corte uno fue de 23 y 26%, respectivamente, valores inferiores a los obtenidos en el presente trabajo (FP1 = 57.8% y RC1 = 52.8%) incluso menores a los límites inferiores de los intervalos de confianza correspondientes. Esta situación pudiera explicarse en función del tamaño de muestra utilizado (Márquez–Sánchez y Hallauer, 1970; Hallauer y Miranda, 1981), que para familias de medios hermanos en sorgo debe ser de por lo menos 190 familias (Knapp et al., 1987). En el caso de Pérez–Grajales et al. (1996) se evaluaron sólo 80 FMHM en un solo ambiente, en tanto que en el presente trabajo se evaluaron 200 en tres ambientes; situación que hace más precisa la estimación; sin embargo, como lo señalan los autores citados, es necesario determinar el tamaño de muestra óptimo en tomate de cáscara. Otra posible explicación de las diferencias en las estimaciones es el número de ambientes de evaluación utilizados en cada caso, que típicamente deben ser varios años y localidades para que las estimaciones de los parámetros genéticos sean más confiables (Comstock y Robinson, 1951; Sahagún, 1994).
Las estimaciones de h2 indican que es posible seguir obteniendo avances con la selección en los 13 caracteres evaluados; sin embargo, el coeficiente de variación genética aditiva (CVA), que en los 13 caracteres fluctuó entre 8.2 y 24.4%, para el rendimiento total fue relativamente bajo (10.4%). De hecho, no hay consistencia entre el CVA y , ya que caracteres con prácticamente la misma (RC2 y RTP, por ejemplo) presentan CVA muy diferentes, y viceversa (FP1 y FP2, por ejemplo, con igual CVA y diferente ). Esto puede ser explicado en función de la forma en que se estima cada parámetro, pues mientras que el CVA contiene sólo la , la involucra tanto a como  , lo que indica que no tiene porqué esperarse una correlación perfecta entre CVA y .
, lo que indica que no tiene porqué esperarse una correlación perfecta entre CVA y .
Correlaciones fenotípicas y genéticas aditivas
La correlación fenotípica del rendimiento total con todos los caracteres fue alta, positiva y significativa, excepto con APB que fue negativa y con NFA que fue baja (Cuadro 3). Las correlaciones genéticas aditivas del mismo carácter (RTP) fueron altas y significativas con RC2, RC1, FTP, FP1, NFA y APB, aunque negativa en el último carácter, lo cual indica que dichos caracteres tienen genes pleiotrópicos comunes. Por otra parte, se observa que no hay correspondencia entre las correlaciones fenotípicas y genéticas aditivas, tanto en magnitud como en signo y significancia, posiblemente debido al efecto ambiental involucrado en las correlaciones fenotípicas (Vargas et al., 1982; Holthaus y Lamkey, 1995; Betrán y Hallauer, 1996). La utilidad de las correlaciones estriba en la posibilidad de estimar el efecto que tiene la selección con base en un carácter sobre otros caracteres, además de permitir realizar selección indirecta (Falconer, 1986); sin embargo, para lo segundo debe cumplirse que la correlación genética aditiva (rGA) sea substancial y que la heredabilidad del carácter secundario sea mayor que la del primario (Hallauer y Miranda, 1981). En el presente trabajo se encontró que la rGA entre el rendimiento total y el del corte uno fue más importante que con el del corte dos, de hecho que con todos los otros caracteres, lo que sugiere que FP1, RC1 y FTP, son los componentes más importantes del rendimiento, ya que sus correlaciones con éste fueron casi el doble de las correspondientes en el corte dos (FP2 y RC2). En el contexto de selección indirecta, es destacable la correlación positiva de RTP con NFA, RC1 y FP1, y negativa con APB, ya que esto puede significar que es posible hacer selección temprana con base en mayor NFA, FP1 y RC1 y menor APB, pues estos caracteres presentaron una mayor que la de RTP (Cuadro 2); desde luego, el corte uno, considerando tanto número como peso de frutos (FP1 y RC1), debiera ser el principal criterio de selección, tanto por sus parámetros genéticos como por su importancia per se.
Las correlaciones genéticas aditivas de APB y ATP con los principales componentes del rendimiento total (NFA, FP1, FP2, FTP, RC1 y RC2) y con este mismo, fueron negativas y significativas (excepto en ATP con NFA y RC1 donde no hubo significancia). Esto indica que las plantas de porte bajo (menor APB y ATP) son más rendidoras que las de porte alto, resultado consistente con lo reportado por Peña et al. (1991) en el sentido de que la variedad Rendidora (de hábito de crecimiento rastrero) superó significativamente a la Salamanca (de crecimiento erecto). Las correlaciones de la LRP con los componentes directos del rendimiento no fueron significativas (excepto con FP1); aunque, al igual que ATP, sí lo fueron con el peso por fruto (PF1, PF2 y PFP). Esto indica que a mayor LRP, correlacionada positiva y significativamente con ATP, se obtienen frutos más grandes, lo cual es razonable si se considera que la mayor longitud de rama se puede deber a una mayor longitud de entrenudos (lo que habría que comprobarse) y por ende la planta presenta menos nudos y menor número de frutos (se produce una flor por nudo), disponiendo éstos de más fotoasimilados para su crecimiento. Al respecto, la variedad Salamanca es más tardía que la Rendidora y produce frutos más grandes, aunque es de menor rendimiento total (Peña et al., 1991).
CONCLUSIONES
El proceso de selección ha reducido la variabilidad genética aditiva en la variedad CHF1–Chapingo, principalmente en el rendimiento total, altura de planta y longitud de rama. La heredabilidad estimada de los 13 caracteres estudiados fluctuó entre 37.1 y 57.8%, por lo que aún es posible obtener avances con el mejoramiento genético por selección. El coeficiente de variación aditiva osciló entre 8.2 y 24.4%, para rendimiento total fue de 10.4%. El número y peso de frutos en el primer corte y frutos totales por planta fueron los principales componentes del rendimiento total, ya que sus correlaciones genéticas aditivas (rGA) con éste fueron las más altas. Las rGA entre el rendimiento total y el rendimiento en el primer corte (0.67), frutos por planta en éste (0.50), número de frutos amarrados (0.38) y altura a la primera bifurcación (–0.49) fueron altas, por lo que es posible hacer selección temprana con base en los dos últimos caracteres y usar el corte uno como criterio principal. Las plantas de porte bajo son más rendidoras que las de porte alto, ya que las rGA de altura a la primera bifurcación y altura total de planta con frutos amarrados, cosechados y rendimiento fueron negativas.
LITERATURA CITADA
AYALA P., J. P. ; PEÑA L., A.; MULATO B., J. 1992. Caracterización de germoplasma de tomate de cáscara (Physalis ixocarpa Brot.) en Chapingo, México. Revista Chapingo 79/80: 128–137. [ Links ]
BETRÁN, F. J.; HALLAUER, A. R. 1996. Characterization of interpopulation genetic variability in three hybrid maize populations. Journal of Heredity 87: 319–328. [ Links ]
CANTWELL, M.; FLORES–MINUTTI, J.; TREJO–GONZÁLEZ, A. 1992. Developmental changes and postharvest physiology of tomatillo fruits (Physalis ixocarpa Brot.). Scientia Horticulturae 50: 59–70. [ Links ]
CARTUJANO E., F.; FERNÁNDEZ O., V. M.; JANKIEWICS, L. S. 1987. Tomate de cáscara variedad rendidora: desarrollo y fenología. Revista Chapingo 56/57: 48–50. [ Links ]
CHRISTIE, B. R.; SHATTUCK, V. I. 1992. The diallel cross: Design, analysis, and use for plant breeding. Plant Breeding Reviews 9: 9–35. [ Links ]
COMSTOCK, R. E.; ROBINSON, H. F. 1951. Consistency of estimates of variance components. Biometrics 7(1): 75–82. [ Links ]
COMSTOCK, R. E.; ROBINSON, H. F. 1952. Estimation of average dominance of genes. In: Heterosis, J. W. GOWEN (ed.). Iowa State University Press, Ames. pp: 494–516. [ Links ]
ESTRADA M., A. 1977. Selección masal y modificada mazorca por surco en dos variedades de maíz de la raza Zapalote Chico. Tesis de Maestría en Ciencias. Colegio Superior de Agricultura Tropical. Cárdenas, Tabasco, México. 164 p. [ Links ]
FALCONER, D. S. 1986. Introducción a la Genética Cuantitativa. Trad. F. Márquez S. Editorial CECSA. 2a. Edición. México. 383 p. [ Links ]
HALLAUER, A. R.; MIRANDA F., J. B. 1981. Quantitative Genetics in Maize Breeding. First Edition. Iowa State University Press, Ames, Iowa. 468 p. [ Links ]
HOLTHAUS, J. F.; LAMKEY, K. R. 1995. Population means and genetic variances in selected and unselected Iowa Stiff Stalk Synthetic maize populations. Crop Sci. 35: 1581–1589. [ Links ]
KNAPP, S. J.; STROUP, W. W.; ROSS, W. N. 1985. Exact confidence intervals for heritability on a progeny mean basis. Crop Sci. 25: 192–194. [ Links ]
KNAPP, S. J.; ROSS, W. N.; STROUP, W. W. 1987. Precision of genetic variance and heritability estimates from sorghum populations. Crop Sci. 27: 265–268. [ Links ]
LÓPEZ M., R.; SANTIAGUILLO HERNÁNDEZ, J. F.; PEÑA LOMELÍ, A.; CUEVAS SÁNCHEZ, J. A.; SAHAGÚN CASTELLANOS, J. 1994. Evaluación de 60 colectas de tomate de cáscara (Physalis ixocarpa Brot.) en Chapingo, México. Revista Chapingo Serie Horticultura 1(2): 131–134. [ Links ]
MÁRQUEZ SÁNCHEZ, F. 1985. Genotecnia Vegetal: Métodos–Teoría–Resultados. Tomo I. Ed. AGT. 1a. Edición. México, D. F. 359 p. [ Links ]
MÁRQUEZ SÁNCHEZ, F.; HALLAUER, A. R. 1970. Influence of sample size on the estimation of genetic variances in a synthetic variety of maize. I: grain yield. Crop Sci. 10: 357–361. [ Links ]
MÁRQUEZ–SÁNCHEZ, F.; SAHAGÚN CASTELLANOS, J. 1994. Estimation of genetic variances with maternal half–sib families. Maydica 39: 197–201. [ Links ]
MATHER, K.; JINKS, J. L. 1971. Biometrical Genetics: The study of continuous variation. Chapman and Hall Ltd. 2a Edición. Great Britain. 377 p. [ Links ]
MOLINA G., J. D. 1992. Introducción a la Genética de Poblaciones y Cuantitativa (algunas implicaciones en genotecnia). AGT Editores. Primera edición. México, D. F. 349 p. [ Links ]
MORENO M., M.; PEÑA LOMELÍ, A.; SAHAGÚN CASTELLANOS, J.; RODRÍGUEZ PÉREZ, J. E.; MORA AGUILAR, R. 2002. Varianza aditiva, heredabilidad y correlaciones en la variedad M1–Fitotecnia de tomate de cáscara (Physalis ixocarpa Brot.). Revista Fitotecnia Mexicana 25(3): 231–237. [ Links ]
MULATO B., J.; FERNÁNDEZ O, V. M.; JANKIEWICS, L. S. 1987. Tomate de cáscara: desarrollo y fenología. Revista Chapingo 56/ 57: 44–47. [ Links ]
NYQUIST, W. E. 1991. Estimation of heritability and prediction of selection response in plant populations. Critical Reviews in Plant Science 10(3): 235–322. [ Links ]
PEÑA LOMELÍ, A.; MÁRQUEZ SÁNCHEZ, F. 1990. Mejoramiento genético de tomate de cáscara (Physalis ixocarpa Brot.). Revista Chapingo 71/72: 85–88. [ Links ]
PEÑA LOMELÍ, A.; RAMÍREZ P., F.; CRUZ G., R. A. 1991. Edad al trasplante en tomate de cáscara (Physalis ixocarpa Brot.) en Chapingo, México. Revista Chapingo 73/74: 57–60. [ Links ]

