











 nova página do texto(beta)
nova página do texto(beta) Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO 
 Permalink
PermalinkIntroduction
Taxonomic and biostatigraphic research in the Mexican Late Tertiary sedimentary basins has shown a highly diversified mammal fauna. Among the best known and important local faunas are those from Rancho El Ocote in the San Miguel Allende basin, Guanajuato, and from Santa María in the Tecolotlán basin, Jalisco. The faunal complex known for those late Hemphillian localities includes the fol-lowing: equids, Nannippus aztecus, Neohipparion eurystyle, Astrohippus stockii and Dinohippus mexicanus; rhinoceros, Teleoceras fossiger; carnivores, Agriotherium schneideri, Borophagus secundus, Canis ferox, and Machairodus cf. M. coloradensis; camelids, Hemiauchenia vera, Megatylopus matthewi, and Alforjas sp.; rodents, Spermophilus sp., Ammospermophilus sp., Paenemarmota sp., Perognathus sp., ?Pliogeomys sp., Calomys sp., Baiomys sp., Prosigmodon sp. and Neotoma sp.; the xenarthran Megalonyx sp.; lagomorphs, Hypolagus mexicanus and Notolagus velox; and the proboscidean Rhynchotherium sp. (Miller and Carranza-Castañeda, 1984, 1998; Carranza-Castañeda, 2006; Jiménez-Hidalgo and Carranza-Castañeda, 2010; Carranza-Castañeda et al., 2013). These faunas constitute the basis for the late-Hemphillian biostratigraphy of central Mexico.
Carbon isotope values from dental enamel record the photosyn-thetic pathways of plants consumed by the animal in an early life stage when the molars were being formed (Koch, 2007). On the other hand, the mesowear method is based on the attrition (tooth-on-tooth contact) and abrasion (tooth-on-food contact) of the teeth (largely their cusps), regardless of the photosynthetic pathway used by the plant that gener-ates it (Kaiser and Solounias, 2003). This method assesses the dental wear caused by the animal’s diet during months to years of its life span, i.e., the long-term alteration of the enamel (Fortelius and Solounias, 2000). The abrasion produced by the intake of abiotic material (e.g. sand, dust, or soil) could be many times more severe than the effect of enamel attrition (Sanson et al., 2007).
Analysis of carbon and oxygen stable isotopes in fossil molars of the equids Astrohippus stocki, Dinohippus mexicanus, Nannipus aztecus, and Neohipparion eurystyle from Yepómera, Chihuahua, and Rancho El Ocote, Guanajuato, suggested that these species fed predominantly on C4 plants, such as grasses or other herbaceous plants (MacFadden, 2008). On the other hand, a mesowear analysis of molars from D. mexicanus and N. eurystyle from the Tecolotlán basin (Jalisco) indicated that N. eurystyle had specialized in feeding on abrasive material (such as grass or external grit), whereas D. mexicanus was a mixed feeder (Barrón-Ortiz and Guzmán, 2008). A combination of mesowear, micro-wear and stable isotope analyses (Bravo-Cuevas et al., 2015) suggested that A. stocki from Rancho El Ocote had a C3/C4 mixed diet; most C3 plants are shrubs and trees, with a few C3 grasses. Unfortunately, those authors did not provide the stratigraphic context of their specimens. Carbon and oxygen stable isotopes in molars of D. mexicanus and N. eurystyle from the Tecolotlán basin have not previously been analyzed. In the present study, enamel from the molar of an individual from each species from Tecolotlán basin was analyzed and the data were compared with those from the molar of an individual of each species from Rancho El Ocote. The objective was to detect differences and similarities in diet of the two species in comparison with the results recorded by MacFadden (2008).
δ13C and δ18O biogeochemical markers
Since 1980, the application of carbon and oxygen stable iso-topes analysis has been important in inferring diet and habitat of Cenozoic herbivorous mammals and birds (Barrick, 1998; Koch, 1998; MacFadden and Higgins, 2004; Angst et al., 2014). Carbon is incor-porated into animals by three plant photosynthetic pathways. The C3 pathway has δ13C from -22‰ to -35‰ and is found in trees and shrubs, as well as in some cold-zone grasses (Smith and Epstein, 1971; O’Leary, 1981; Medina et al., 1986). The C4 pathway has δ13C values from -10‰ to -12‰ and is typical for grasses, and also for some dry-zone trees and shrubs (Stowe and Teeri, 1978; Cerling, 1999; Cerling and Ehleringer, 2000). Finally, the Crassulacean Acid Metabolism (CAM) pathway has δ13C values from -35‰ to -12‰ and is found in succulent plants such as cacti and some orchids (Gröcker, 1997; Andrade et al., 2007).
Herbivores incorporate the carbon of the plants on which they feed into their organs and tissues, so they have carbon isotope values reflecting those of the plants they feed upon. In the case of tooth enamel, the tissue is enriched by 14.1‰ relative to the plant value (Cerling and Harris, 1999). Animals that feed on C3 plants have carbon isotopic values from -19‰ to -9‰, while herbivores that have consumed C4 plants have carbon isotopic values from -2‰ to +2‰. The C3/C4 mixed feeders show values from -9‰ to -2‰ (MacFadden and Cerling, 1996).
On the other hand, oxygen is incorporated into animals by inhala tion, from water in food, and mainly by ingested water. Such oxygen is in equilibrium with what is lost through CO2 exhalation, feces, urine and sweat (Koch et al., 1994; Sánchez, 2005). Given that ingested water comes from meteoric water, its composition is affected primarily by temperature, but also by altitude, latitude and the amount of rain fall in a zone (Dansgaard, 1964). The oxygen composition of dental enamel from mammals is used mainly for inferring the paleoclimatic conditions that prevailed in a locality in the past (Longinelli and Nuti, 1973; Kohn, 1996; Kohn et al., 1998; Schoeninger et al., 2000; Mahboubi et al., 2014).
Study localities
Rancho El Ocote is located at 21º05´28”N and 100º41´01”W in the San Miguel Allende basin, Guanajuato. The ashes found in the stratigraphic sequence were dated by fission tracks, and provided ages of 4.8 Ma for the lower sediments (the Rhino layer) formed during the late Hemphillian, and 4.7 Ma for the upper ash from the early Blancan (Kowallis et al., 1998). These ages provide the limits of the North American Land Mammal Ages (NALMA), late Hemphillian and early Blancan (Figures 1 and 2) (Flynn et al., 2005; Carranza-Castañeda, 2006; Cohen et al., 2013). Four equid species, D. mexicanus, N. eurystyle, A. stockii, and N. aztecus occur in the Rhino level, with lithology and sedimentation rates showing a more humid environment than those of the overlying Blanco level, where only D. mexicanus and N. neurystyle are found (Carranza-Castañeda, 2006).
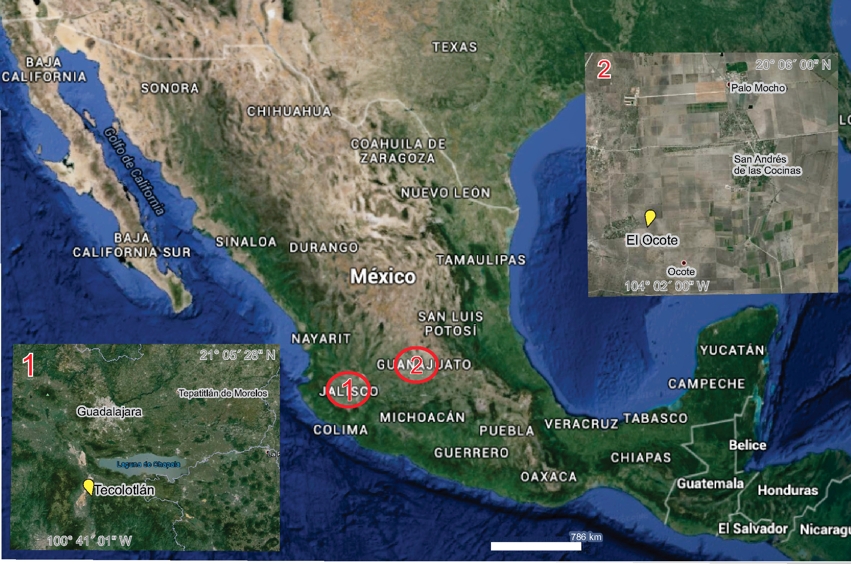
Figure 1 Late Hemphillian fossiliferous localities in the Tecolotlán basin (1) and Rancho El Ocote (2), Mexico.
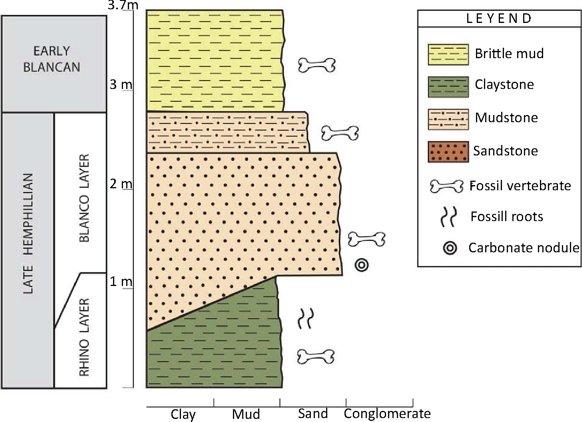
Figure 2 Stratigraphic column for Rancho El Ocote (GTO 2A). Investigated specimens came from the Rhino layer. Taken from Pacheco-Castro and Carranza-Castañeda (2015).
The molars from Rancho El Ocote were collected at the GTO-2A locality. Carranza-Castañeda et al. (2013) recorded that Dinohippus mexicanus and Neohipparion eurystyle were found in the Rhino level, and to Hh3 in the Mammalian Chronology proposed by Tedford et al. (2004). On the other hand, N. eurystyle was not found above the Rhino level. This implies that the level above the Rhino level represents the final portion of the late Hemphillian (Hh4) (Figure 2).
The Tecolotlán basin is 100 km southeast of Guadalajara, in the state of Jalisco (Figure 1; the late Hemphillian stratigraphic sequence is divided by lacustrine sands). The best faunal representation is from the Santa María locality and the studied molars were collected at this site (Figure 3). Lower ash has been dated by 40Ar/39Ar at 4.89 Ma (Kowallis et al., 1998). Carranza-Castañeda et al. (2013) have shown that the Santa María area belongs to Hh3. As an erosional discordance, the late Blancan-Irvingtonian, Buenaventura sequence contains gravel and sands, with an internal ash dated at 2.6 Ma (Kowallis et al., 1998). Santa María has a stratigraphic sequence in the Tecolotlán basin that terminates at the bottom of the lacustrine sands. Lithology and sedimentation rates at Santa María indicate the presence of a humid environment, containing remains of D. mexicanus, N. eurystyle, A. stockii and N. aztecus. The last two species are not found in the San José stratigraphic sequence that is overlying the lacustrine sands, with lithology and sedimentation rates typical for a dry environment (Carranza-Castañeda et al., 2013).

Figure 3 Statigraphic column for the Tecolotlán basin (Santa María). Investigated specimens came from the bone layer. B: Bone layer, K: Cretaceous. Taken from Carranza-Castañeda (2006).
Materials and methods
Sample extraction, preparation and statistical analysis of the results
A bulk sample was taken from isolated teeth from Rancho El Ocote: MPGJ-2056 (Dinohippus mexicanus), MPGJ-2026 (Neohipparion eurystyle) and Tecolotlán Basin: MPGJ-2956 (D. mexicanus), MPGJ-2959 (N. eurystyle), each of them representing a single individual. The samples were taken at the crown-root joint following the protocol of Feranec and MacFadden (2000) and belong to the Palaeontological Collection of the Centro de Geociencias of the Universidad Nacional Autónoma de México (UNAM), Campus Juriquilla, Querétaro.
Samples were processed in the Stable Isotope Laboratory at the Instituto de Geología, UNAM, with the method proposed by Koch et al. (1997). First, 20 mg of enamel was ground and sieved (125 μm mesh) to obtain a fine and uniform powder. Then 10 ml of hydrogen peroxide at 30 % was added to eliminate the organic matter. After 2 h, the samples were centrifuged and the hydrogen peroxide de-canted and washed again three times with water type I (grade HPLC 18.2 MΩ). Once the washing was finished, 5 ml of a buffer solution, Ca(CH3CO2)2-CH3COOH 1.0 M, pH 4.75, was added and allowed to rest for 9 h. The buffer solution was decanted and the samples were washed again three times with water type I. Finally, to eliminate any remaining water, ethanol was added, and the solution was left for 20 h in an oven at 90 oC. Isotopic ratios were determined with a Finnigan MAT 253 mass spectrometer with a dual inlet system and auxiliary Gas Bench equipment with a GC Pal autosampler with a temperature-controlled aluminum plate adjoined to the mass spectrometer (Révész and Landwehr, 2002). Results were reported as δ18OVPDB and δ13CVPDB, and they were normalized using NBS-19, NBS-18 and LSVEC to the Vienna Pee Dee Belemnite (VPDB) scale in accordance with the cor-rections described by Coplen (1988), Werner and Brand (2001) and Coplen et al. (2006). For this technique, the standard deviation was 0.2‰ for oxygen and carbon.
Results
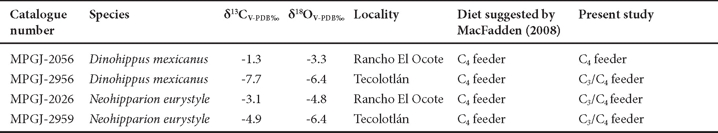
Carbon and oxygen isotopic values for D. mexicanus from Rancho El Ocote are more positive than those from Tecolotlán (Table 1, Figure 4). The 13C values for specimens of N. eurystyle from the two sites are similar to each other, although somewhat higher from Rancho El Ocote than from Tecolotlán (Table 1, Figure 4).

Discussion
Diet
The carbon isotope value (δ13C: -1.3‰) for Dinohippus mexicanus (MPGJ-2056) from Rancho El Ocote implies that this individual was a specialist feeding on C4 plants such as grasses or other herbs (Table 1), also observed by MacFadden (2008) for Rancho El Ocote and Yepómera locations. In contrast, individuals found in Florida and Texas were mixed C3/C4 feeders (MacFadden, 2008), but consumed an important amount of C3 plants, such as C3 grasses, tree leaves and shrubs. On this basis, MacFadden (2008) assumed that, during the late Hemphillian, C4 plants were abundant in Mexican ecosystems, whereas C3 plants were abundant further north. However, the present results with specimen MGPJ-2956 from Santa María (Tecolotlán basin) differ in that they show a δ13C value that characterize it as a C3/C4 mixed feeder, but with an important component of C3 plants (Table 1).
Mesowear studies on specimens of D. mexicanus from the Tecolotlán basin (Barrón-Ortiz and Guzmán, 2008) found both browsers and mixed feeders.
The present isotope analysis has suggested that specimen MPGJ-2956, pertaining to D. mexicanus from Tecolotlán, may have fed on trees, shrubs and C3 grasses, a diet similar to that proposed by Barrón-Ortiz and Guzmán (2008) for other Tecolotlán specimens. MPGJ-2956 was collected in the lower stratigraphic sequence of the Tecolotlán basin (Carranza-Castañeda, 2006), whereas Barrón-Ortiz and Guzmán (2008) assumed that their specimens likely came from the upper part of the sequence. Their lack of stratigraphic data precludes direct comparison of the two studies; it is uncertain whether the indi-viduals represented by the different specimens were contemporaneous and whether the diet of this species remained constant through time. In Costa Rica, D. mexicanus seems to have had a generalist, not a special-ized, diet based on morphological ground (Laurito and Valerio, 2010), similar to that suggested by the isotopes in specimens from Texas, Yepómera and Rancho El Ocote (MacFadden, 2008).
In the case of N. eurystyle, carbon isotope values for both MPGJ-2026 from Rancho El Ocote and MPGJ-2959 from the Tecolotlán basin show that these animals were C3/C4 mixed feeders, with in the case of MPGJ-2026, an emphasis on the intake of C4 plants (Table 1). This pattern is different from what was found by MacFadden (2008) with specimens from Rancho El Ocote and Yepómera, which fed exclusively on C4 plants. Mesowear analyses (Barrón-Ortiz and Guzmán, 2008) suggested that N. eurystyle from the Tecolotlán basin was a grazer, but the lack of stratigraphic control makes it difficult to ascertain whether the diet variation between individuals was due to different locations from the same time period, or whether the differences were due to environmental change.
Since the individuals of N. eurystyle analyzed here appear to have been C3/C4 mixed feeders like some individuals in Florida (MacFadden, 2008), both N. eurystyle and D. mexicanus from the Mexican late Hemphillian appear to have been generalists in their diet.
This plasticity in equid diets may be explained by different fac-tors. For example, in desert areas in the southern USA and northern Mexico, C3 plants are more abundant in winter, the rainy season, whereas C4 plants are more abundant in summer (Ehleringer and Monson, 1993). In temperate areas in general, the two kinds of plants co-inhabit year-around, but in microhabitats that differ in tempera-ture and humidity. Isotope analysis has shown that present-day feral horses in southern New Mexico feed on leaves and grasses that differ in proportion according to seasonal availability (Hansen, 1976; Smith et al., 1998). Furthermore, factors such as soil salinity, light intensity and soil nutrients may influence the carbon isotope composition of plants (Bocherens, 2003) and hence the composition of the dental enamel of herbivores.
Competition with other herbivore species may in part explain the wide feeding spectra shown by these equids; Late-Pleistocene bison and horses at Rancho La Brea, California, fed mainly on C3 plants, but bison more frequently preferred C4 plants (Feranec et al., 2009), suggesting the presence of resource partitioning. The small sample size in the present study precludes further speculation.
Habitat
The carbon and oxygen isotope values of both specimens from Rancho El Ocote suggest a preference for open vegetation zones (Figure 4). In the case of the specimens from the Tecolotlán basin, N. eurystyle lived in grassland or savannas, whereas D. mexicanus preferred areas with major tree cover. In Florida, N. eurystyle seems to have inhabited open zones, whereas D. mexicanus could live in either open regions or areas with some tree cover (MacFadden et al.,1999a; MacFadden, 2008). Hence, D. mexicanus may have been more flexible in its habitat and feeding habits according to local conditions.
Although D. mexicanus had a mixed feeding habit associated with environments of mixed vegetation and high humidity (Laurito and Valerio, 2010), some individuals were grazers living in herbaceous grassland and arid areas. There is a latitudinal gradient: above 35 o-40 oN, C3 plants such as grasses, shrubs and trees are abundant, with diminishing C4 grasses (MacFadden et al., 1999b); this is reflected in the differences in diet between specimens from the USA and those from Mexico (MacFadden, 2008).
The results of mesowear analyses of D. mexicanus and N. eurystyle from the Tecolotlán basin suggest that this basin developed a heterogeneous vegetation of trees and grasses fostered by a seasonal climate (Barrón-Ortiz and Guzmán, 2008); dry seasons were favorable for C4 plant growth, and wet seasons were favorable for more abundant C3 plants. In the Santa María locality, lacustrine sediments and paleochannels indicate the presence of water bodies such as rivers, lakes and water channels (Carranza-Castañeda, 2006). Fossil freshwater gastropods, teeth of Crocodylus sp., all aquatic animals, and remains of the rhinoceros Teleoceras fossiger, have been found there (Carranza-Castañeda et al., 2005). Although T. fossinger has been defined as a browser, isotopic analyses of specimens from Florida showed its diet to be more generalist (MacFadden, 1998), so its presence would not necessarily indicate the existence of C3 plants in the locality. Nevertheless, the wet environment would have been favorable to C3 plants (Ehleringer and Monson, 1993). Abundance of C3 plants at Santa María did not depend on the presence of a latitudinal gradient, but on specific conditions at each locality.
The conclusions above are supported by the oxygen isotope values: D. mexicanus from Santa María shows a δ18O value of -6.4‰, whereas from Rancho El Ocote it is -3.3‰. For N. eurystyle from Santa María it is -6.4‰, whereas from Rancho El Ocote it is -4.8‰. With increasing amounts of rain, oxygen isotope values become more negative (Dansgaard , 1964). Because Santa María and Rancho El Ocote are at different altitudes and latitudes, differences in δ18O values are to be expected. Additionally, the location of Santa María on the foothills of the Sierra Madre Occidental implies that it received more precipitation, creating more humidity favorable for the establishment of C3 plants, as reflected by the lacustrine sediments (Carranza-Castañeda, 2006). Another factor, however, could explain the different oxygen isotope values. Both D. mexicanus and N. eurystyle may have migrated from place to place in search of food (MacFadden, 2008). They drank water from different feeding areas, and hence the isotopic composition could differ, as found for bison in the eastern Great Plains (Widga et al., 2010).
Carbon isotope analyses on the two other equid species found at Rancho El Ocote, A. stocki and N. aztecus, showed that some of these individuals were C3/C4 mixed feeders, suggesting that this area was drier than at the Florida and Texas localities, and favored the presence of C4 plants (MacFadden, 2008). However, no inference could be made regarding whether during the late Hemphillian grassland with some trees or shrubs existed.
On the basis of the diversity of fossil herbivorous species found at Rancho El Ocote or in the Tecolotlán basin, other species should be included in subsequent studies in order to draw more precise infer-ences regarding the vegetation type and environmental conditions that existed at each locality during the late Hemphillian.
Conclusions
At Rancho El Ocote, the diet of the Dinohippus mexicanus speci-men was primarily based on C4 plants, and this animal preferred open zones, such as grasslands or savannas. In contrast, the Neohipparion eurystyle was a C3/C4 mixer feeder with an emphasis on C4 plants, living in open zones.
In the Tecolotlán basin, the Dinohippus mexicanus and Neohipparion eurystyle were C3/C4 mixed feeders and lived in areas with some tree coverage; this suggests that conditions here were more favorable for C3 plants than at Rancho El Ocote.
These preliminary conclusions require substantiation by analyses of more specimens and from a wider range of species.

