











 nueva página del texto (beta)
nueva página del texto (beta) Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkINTRODUCTION
Deccan continental flood basalts are one of the largest magmatic provinces, occupying an area of about 500000 km2 in peninsular India (Jay and Widdowson, 2008), and straddling the Cretaceous-Palaeogene (K-Pg) boundary (Courtillot et al., 1986; Duncan and Pyle, 1988; Chenet et al., 2007; Keller et al., 2009a-b-c, 2010a, b, 2012; Malarkodi et al., 2010; Khosla and Verma, 2015; Schoene et al., 2015). The age and duration of the Deccan activity have come under intense scrutiny. In the decade of 1980, Courtillot et al. (1986) and Duncan and Pyle (1988) suggested that duration of volcanic eruptions spanning less than 1 m.y. in magnetic polarity chron 29r. More recently, radiometric dating confirmed the chron C29r age for the bulk of the eruptions followed by another pulse near the base of C29n (64.7± 0.6: Ma, Chenet et al., 2007) and an initial smaller eruption at 67.4 Ma near the base of C30n. This indicates that Deccan eruptions occurred intermittently over an interval of at least 3 m.y., although some earlier studies suggest an even longer duration spanning 4 to 5 m.y. (Courtillot, 1990; Pande et al., 2004) or even 7 to 8 m.y. (Sheth et al., 2001).
Current palaeomagnetic, radiometric, biostratigraphic, and chemostratigraphic data suggest that the main phase of Deccan volcanism, which accounts for approximately 80% of the 3500 meter thick continental basalts, erupted within a short period (less than 1 m.y.) or even less (a couple of hundred thousand years) mostly near the end of C29r, which spans the K-Pg boundary (Chenet et al., 2007; Keller et al., 2008, 2009a, b, c, 2010a, b, 2011, 2012; Gertsch et al., 2011; Khosla and Verma, 2015; Schoene et al., 2015). Volcanic eruptions may have played a major role in mass extinctions at the K-Pg boundary. According to Schulte et al. (2003, 2010) Chicxulub impact had generated a tsunami, with impact spherules in the latest Maastrichtian sediments in Mexico and Texas. They regarded Deccan volcanism as much older in age and have not played any role in K-Pg mass extinction. Schulte et al. (2010) also considered Chicxulub impact as the solitary cause for the mass extinction at K-Pg boundary and they also assumed that Chicxulub and the iridium anomaly at the Cretaceous-Palaeogene are heritably related and are of similar age. Keller et al. (2010a, b) refuted the above listed assumptions as proposed by Schulte et al. (2010). Keller et al. (2010a, b) and Keller (2014) further stated that there is no record of iridium anomaly in alliance with Chicxulub ejecta, and impact spherules have never been recognized in the iridium enriched K-Pg clay layer in Mexico. Recent work on Deccan volcanism clearly indicates that the main mass extinction is linked with the major phase of Deccan eruptions in C29r (Keller et al., 2009c, 2010a, b, 2012; Keller, 2014).
Chenet et al. (2007) suggested that the Deccan volcanism was at its acme at 65 Ma and initial eruptions took place in the Nasik and Narmada (north-western) region (Widdowson et al., 2000). Later volcanism occurred on the southern sector (Beane et al., 1986; Mitchell and Widdowson, 1991), and the final phase of eruption occurred in the southern part of the Deccan Province near Belgaum (Jay and Widdowson, 2008). However, it is only recently that decisive evidence in this regard has emerged (Keller et al., 2008, 2009a, b, c, 2010a, b, 2011).
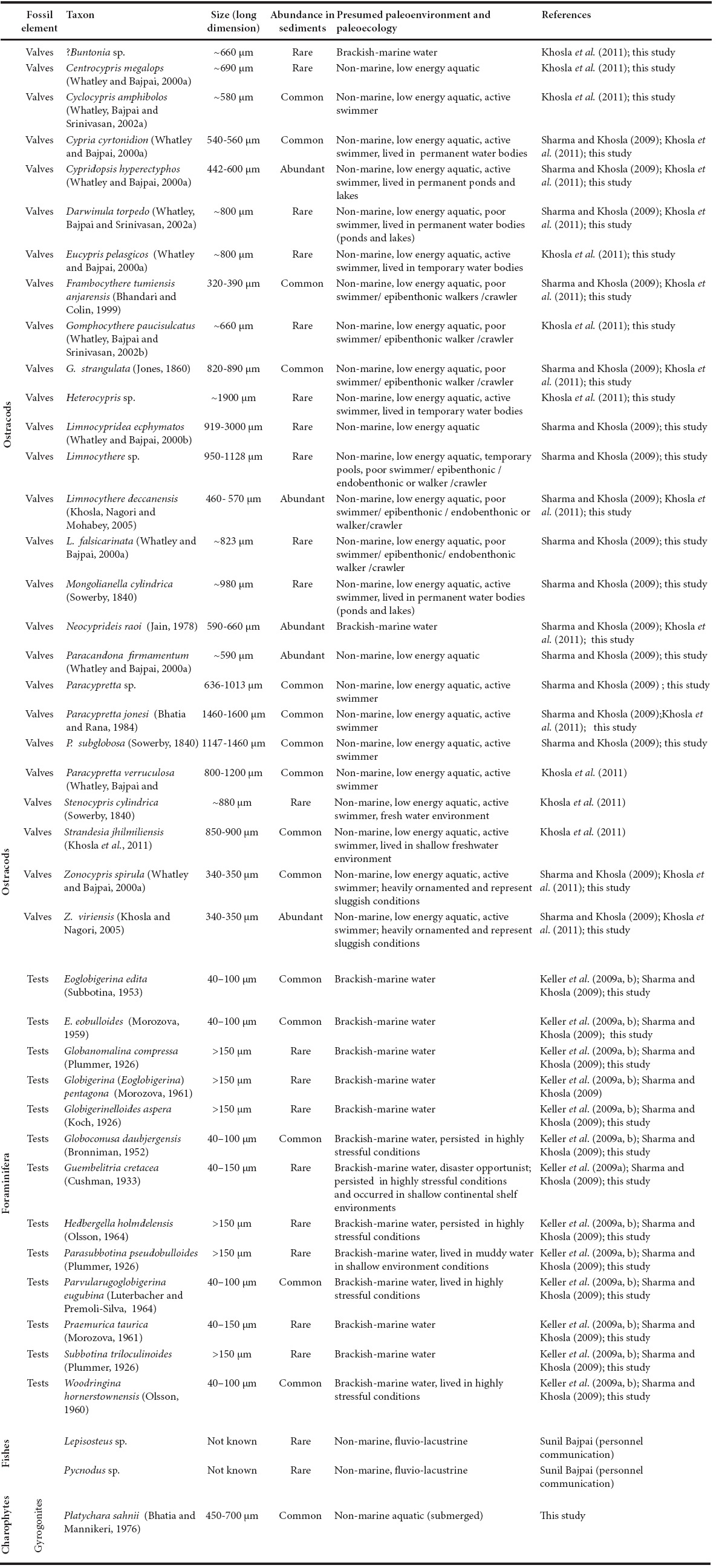
The infraand intertrappean sediments associated with the Deccan Traps have been widely studied for more than two decades leading to a better understanding of the biotic diversity (Figure 1; Khosla, 1994, 2001; Khosla and Sahni, 1995, 2000, 2003; Khosla et al., 2004, 2009; Prasad et al., 2007a, b, 2010; Keller et al., 2009a-b-c, 2012) and the palaeobiogeographic relationships of the Indian subcontinent during its northward passage (Loyal et al., 1996; Prasad et al., 2010; Verma et al., 2012; Khosla, 2014; Khosla and Verma, 2015; Fernandez and Khosla, 2015; Khosla et al., 2015). Further, these unique deposits have permitted examination of the effects of volcanism on the biota (Bajpai and Prasad, 2000). The Deccan intertrappean outcrops have yielded diverse fauna and flora represented by vertebrates (fishes, anurans, lizards, snakes, turtles, crocodiles, dinosaurs and mammals), mollusks, ostracods, and plants (megaplants, palynofossils and charophytes; Khosla and Sahni, 2003). At present, the majority of intertrappean beds fringing the main Deccan basaltic province have been equated with the Ambenali and Poladpur formations of the classic Western Ghats sections (Widdowson et al., 2000). These beds have produced dinosaur remains including eggshells, together with diagnostic Late Maastrichtian ostracod assemblages (Sahni and Khosla, 1994a, b; Whatley and Bajpai, 2000a, b; Whatley et al., 2002a, b; Vianey-Liaud et al., 2003; Whatley and Bajpai 2005, 2006; Khosla et al., 2005; Khosla and Nagori 2007) and a palynofloral assemblage consisting of Aquilapollenites-Gabonisporites-Ariadnaesporites (Kar and Srinivasan, 1998). A few intertrappean outcrops, such as Papro in the Lalitpur District, Uttar Pradesh (Singh and Kar, 2002; Sharma et al., 2008), Ninama in Gujarat (Samant et al., 2014) and Rajahmundry, Andhra Pradesh, have been assigned to the early Paleocene (Keller et al., 2008, 2009a, b, 2011; Malarkodi et al., 2010) based on palynofossils, ostracods and foraminifera. It is interesting to note that the deep exploratory oil wells of the Oil and Natural Gas Commission (ONGC) at Narsapur have yielded Maastrichtian planktonic foraminiferal assemblages at depths exceeding 3 km (Govindan, 1981; Keller et al., 2008, 2011), together with an Aquilapollenites pollen assemblage (Kar et al., 1998). The documentation of a planktonic foraminiferal assemblage from Jhilmili intertrappean beds (Figures 2a, 2b) assigned to the Early Danian (P1a) (Keller et al., 2009a-b-c; Sharma and Khosla, 2009) just north of Chhindwara town and in the heart of peninsular India, has interesting implications for constraining the age limits of basaltic flows. The occurrence of freshwater taxa such as vertebrates, mollusks and algae in association with brackish water ostracods (Kar and Srinivasan, 1998; Khosla and Nagori, 2007; Keller et al., 2009a-b-c, 2010a, 2011; Samant and Mohabey, 2009; Sharma and Khosla, 2009) in the nearby Mohgaon Kalan and Singpur localities has also raised questions about the depositional environment of the intertrappean beds of this region. The fauna and floral assemblages recovered from the Jhilmili intertrappeans are listed in Table 1.

Figure 1 Distribution of Cretaceous-Palaeogene (K-Pg) Deccan volcanics, showing the major infratrappean and intertrappean fossiliferous localities. The localities are marked by fresh water, brackish water and marine environments. Marine incursions are along the Narmada-Tapti rift and possibly also along the Godavari graben.

Figure 2 Location (a) and lithostratigraphy (b) of the Jhilmili intertrappean section, District Chhindwara, Madhya Pradesh, peninsular India.
Table 1 List of fauna and flora recovered from the Jhilmili intertrappean section, Chhindwara District, Madhya Pradesh, Central India.
The main issue that is pertinent to the current discussion is how planktonic foraminifera could be found so far inland (Keller et al., 2009a, b). Sahni (1983) proposed the presence of a marine seaway "Trans Deccan Strait" based on the occurrence of an admixture of freshwater, brackish water and marine forms in intertrappean beds of Asifabad and Nagpur and the Lameta Formation of Pisdura and Jabalpur. The record of planktonic foraminifera at Jhilmili revive interest in the role that the two major structural lineaments, namely the Narmada and Godavari, played in the anomalous admixture of the biota's representing diverse facies (Figure 1). The main objective of this paper is to analyze the palaeoenvironments, palaeoecology and palaeobiogeographical implications of the recovered micro biota from Jhilmili intertrappean beds.
GEOLOGICAL SETTING AND LITHOLOGY
The intertrappean section of Jhilmili (Lat. 22° 02' 44" N; Long. 79° 09' 34' E; Figures 2a, 2b and 3a, 3b) is situated about 5 km NW of the well known village of Mohgaon Kalan on the Seoni-Chhindwara road in the Chhindwara District, Madhya Pradesh (central India). This intertrappean section was discovered and sampled by A. Khosla and A. Sahni in 1999. Subsequently this intertrappean outcrop was studied by a multidisciplinary and multi-institutional international team (Keller et al., 2009a, b). The exposed intertrappean deposit is 14 m thick and bounded by Deccan lava flows at the base and top (Figure 2b). The basal unit of this intertrappean section is 6 m thick and consists of red clayey siltstone with carbonate nodules and root traces. The upper unit spans 6.5 m and consists of red and green shales. Both units have been interpreted as paleosols (Keller et al., 2009a, b). In between these two paleosols is a 60 cm thick unit that consists of yellow to pink, ostracodrich shale and calcareous limestone with planktonic foraminifera. The alternating yellow to pink clay and marly limestone mark the lower part of this short interval. Ostracods and charophytes are common in unit 3 (Figure 2b, Figure 4, samples JH17, JH19).

Figure 3 a) Field photograph of the Jhilmili intertrappean beds showing a narrow interval of lacustrine to brackish marine unit 3 (60 cm thick), which has yielded ostracods and planktonic foraminifera. b) Enlarged, close up view of the fossiliferous light yellow clays with hard claystone bands. Scale (Marker pen) = 15 cm.

Figure 4 Relative abundance of Jhilmili ostracods (rare, 10 carapaces; common, 10-20 carapaces; abundant, 20 carapaces), charophytes and planktonic foraminifera in unit 3 of the Jhilmili intertrappean sediments. Ostracod assemblages mark predominantly freshwater environments, except for Neocyprideis raoi (Jain, 1978). Modified after Keller et al. (2009b).
MATERIALS AND METHODS
To date five palaeontological field expeditions were carried out and nearly 500 kg of sediments collected, processed and analyzed in the palaeontological laboratories of Indian Institute of Technology (IIT), Roorkee and the Panjab University, Chandigarh. Most samples were collected from the 60 cm thick middle unit 3 composed of yellow to pink ostracod-rich shale and calcareous limestone, which also yielded planktonic foraminifera. This intertrappean unit (Figures 2 and 3) preserves a relatively diverse micro biota (Sharma and Khosla, 2009), that includes ostracods, foraminifera, fishes and charophytes (Table 1). For the recovery of microfossils standard sample processing methods were applied (Keller et al., 1995). The sediment was crushed into small pea-sized pieces and immersed in water for 24 hours to allow complete disaggregation into mud slurry. The sample was then washed through various sets of sieves and the residue was oven dried. The dried residue was scanned under a stereoscopic binocular microscope for microfossils. SEM photomicrographs of planktonic foraminifera and ostracods were taken at Indian Institute of Technology, Roorkee and Panjab University, Chandigarh, by means of a JEOL JSM 6400 SEM (Figures 5a 5r). The described specimens are housed in the Department of Earth Sciences, IIT Roorkee and Vertebrate Paleontological Laboratory of the Department of Geology, Panjab University, India.

Figure 5 a) Neocyprideis raoi (Jain, 1978), carapace, right lateral view; b) Neocyprideis raoi (Jain, 1978), carapace, dorsal view; c) Paracypretta subglobosa (Sowerby, 1840), carapace right lateral view; d) Paracypretta jonesi (Bhatia and Rana, 1984), carapace, dorsal view; e) Zonocypris spirula (Whatley and Bajpai, 2000a), carapace, dorsal view; f) Cypria cyrtonidion (Whatley and Bajpai, 2000a) carapace, left lateral view; g) Mongolianella cylindrica (Sowerby, 1840) carapace, right lateral view; h) Limnocypridea ecphymatos (Whatley and Bajpai, 2000b) carapace, left lateral view; i) Gomphocythere strangulata (Jones, 1860), carapace, dorsal view; j) Frambocythere tumiensis anjarensis (Bhandari and Colin, 1999), carapace, dorsal view; k) Frambocythere tumiensis anjarensis (Bhandari and Colin, 1999), carapace, dorsal view; l) Limnocythere deccanensis (Khosla et al., 2005), carapace, left lateral view; m) Subbotina triloculinoides (Plummer, 1926); n) Guembelitria cretacea (Cushman, 1933); o) Hedbergella cf. holmdelensis (Olsson, 1964); p) Parasubbotina pseudobulloides (Plummer, 1926); q) Globanomalina compressa (Plummer, 1926); r) Globigerina (Eoglobigerina) pentagona (Morozova, 1961). Scale bar equals 100 μm for a; 800 μm for b; 250 μm for c; 200 μm for d; 60 μm for e; 200 μm for f; 400 μm for g; 200 μm for h; 200 μm for i; 100 μm for j; 200 μm for k; 60 μm for l; 100 μm for m, n, o, p, q, r. (Figures a to l modified after Sharma and Khosla (2009) with permission from the Journal of the Palaeontological Society of India and m to r after Keller et al. (2009a) with permission from Journal of the Foraminiferal Research).
PALAEOENVIRONMENTAL AND PALAEOECOLOGICAL IMPLICATIONS OF THE FOSSIL FINDS
Palaeoenvironmental interpretations are based on lithological variations and microfossil assemblages. Six lithological units have been identified within the Jhilmili intertrappean section (Figure 2b). Unit 1 is the basalt, and the lower part of unit 2 is a paleosol; lithologically, it consists of purple siltstone and contains clasts of basalt, which have been derived from the lower part of the basalts. Other palaeosol features include slickenslide structures and rotated quartz grains (Keller et al., 2009a, b). The upper part of unit 2 (JH11) also contains red clayey siltstone with carbonate nodules, manganese streaks and slickensides, which are undoubtedly deposited in terrestrial, palustrine and flood plain environments (Keller et al., 2009a, b). Paleosols (JH14) are well recorded which might be due to the weathering of basalts resulting in a thin purplecoloured paleosol. Unit 2 contains the clay mineral smectite, which has been derived from basalts and indicates semiarid to sub-humid climatic conditions (Keller et al., 2009a, b; Pal et al., 2013). The middle unit 3 (Figure 2b, Figure 4, JH16, JH19, JH22, JH23) of the Jhilmili intertrappean section is 60 cm thick and contains fresh water ostracods and brackish to fresh water charophytes (JH17, JH19), and foraminifera of marine water in nature (Figure 2b, JH16, JH18, JH20-22). The number of ostracod taxa recovered from screenwashed residue (i.e. rare <10, common 10-20 and abundant > 20, Figure 4) suggests that six out of twelve species are common to abundant in, at least, some intervals. Other taxa are rare and intermittently present (Keller et al., 2009a, b). The abundance of various ostracod taxa is shown in Figure 4. Most of the ostracods have been richly recovered from samples JH24 and JH25 (Figures 2b and 4). Of all the ostracod species, Limnocythere deccanensis dominates the ostracod assemblages (80-90 %), whereas Zonocypris viriensis dominates in samples JH17 and JH18 (Keller et al., 2009a, b). Dry and arid climatic conditions have also been recorded in unit 3 because of the presence of clay mineral assemblages such as palygorskyte, and red clayey siltstone with scanty rhizo concretionary structures (Keller et al., 2009a-b-c; Pal et al., 2013). The upper part of unit 3 (JH 24 and JH25) consists of laminated claystone with fresh water and brackish ostracods, charophytes and planktonic foraminifera. This mixed assemblage indicates a shallow marine to brackish water environment. The topmost part of unit 3 (JH26) is characterized by packstone with no foraminifera but containing ostracod shells fragments and charophytes, which mark high energy wave action (Keller et al., 2009a, b). Unit 4 (Figure 2b) is interpreted as a paleosol horizon, and is characterized by red clayey siltstone with abundant Microcodium, whereas unit 5 is a greenish grey siltstone with rare, fine sand layers, which indicates the return of terrestrial deposition (Keller et al., 2009a, b). In units 4 and 5, poorly crystallized illite is present which may have been derived from the degradation of smectite. Both units indicate arid climatic conditions with alternating wet and dry cycles (Keller et al., 2009a, b). Very negative stable isotope (δ13C and δ18O) values have been recorded. The δ13C values (-4.3 %, JH17-JH19) of the foraminifera-bearing clasts reveal fresh to brackish water environments. The heavier δ13C values (-3.3-2.7 %, JH24-JH26) of the ostracod and foraminifera-bearing sediments point to brackish water environments. The δ18O values (-6.2 to -9.5 %) also indicate fresh to brackish water environments (Keller et al., 2009a, b).
Palaeoecologically, the Jhilmili freshwater ostracod assemblage (Figures 5a 5l) represents an admixture of drifters or benthic crawlers/diggers and swimmers. Limnocythere deccanensis and Zonocypris viriensis dominate the recovered assemblage of 26 species from the Jhilmili intertrappean beds (Table 1). Most of the taxa are nonor poor swimmers, such as the cytheraceans (Limnocythere, Gomphocythere, Frambocythere) and darwinulaceans (Darwinula). According to McKenzie (1971), some species of Limnocythere are endobenthonic and spent part of the day within the sediment while the majority of the family members occupied permanent waters and some species of Limnocythere lived in temporary pools. The other two species Gomphocythere and Frambocythere have been considered as epib enthonic crawlers (McKenzie, 1971). Darwinula occurs in streams but prefers to live in permanent water bodies such as lakes and ponds (Whatley and Bajpai, 2005). On the other hand, taxa such as Paracypretta, Zonocypris, Cypridopsis, Cypria and Mongolianella are active swimmers (Table 1). The genus Cypridopsis preferred to live in permanent water bodies like ponds and lakes (Whatley and Bajpai, 2005; Khosla et al., 2011). The genus Strandesia is mainly found in shallow, freshwater environments (Khosla et al., 2011). The extinct genus Mongolianella, with its close morphological similarity (i.e. long, cylindrical shape) to modern Herpetocypris, was probably also a good swimmer (Whatley and Bajpai, 2005). The taxa like Mongolianella and Cypridopsis preferred to dwell in more permanent waters that would allow retreat to the deeper parts of the water body (Whatley and Bajpai, 2005).
Genera like Heterocypris, Eucypris and Strandesia are indicative of temporary water bodies. All of these taxa listed above suggest the presence of a permanent freshwater lake/pond. Further, a clear augmentation in the alkalinity of the environments is indicated by greater calcification of the heavily ornamented ostracods (e.g. in Frambocythere tumiensis anjarensis Bhandari and Colin, 1999; Gomphocythere strangulata Jones, 1860; Paracypretta subglobosa Sowerby, 1840; Paracypretta jonesi Bhatia and Rana, 1984; Zonocypris spirula Whatley and Bajpai 2000a; Zonocypris viriensis Khosla and Nagori, 2005; Limnocypridea ecphymatos Whatley and Bajpai, 2000b; Limnocythere sp. and Paracandona firmamentum Whatley and Bajpai, 2000a). Heavily ornamented ostracods particularly Zonocypris indicates sluggish conditions (Khosla et al., 2011).
Presence of a brackish water ostracod species (Neocyprideis raoi) in sample numbers JH21 to JH26 along with rare planktonic foraminifera points to intervals of a brackish-marine environment. Neocyprideis are mostly represented by moult stages in the present collection and lived with other fresh water ostracod assemblages in the lake at times of low mesohaline salinities (Sharma and Khosla, 2009; Khosla et al., 2011). The brackish water species Neocyprideis raoi was previously recorded from the intertrappean beds of the Rajahmundry area near the southeast coast (Jain, 1978). The occurrence of Neocyprideis in a brackish water environment was indicated by Keij (1957), Morkhoven (1963), Keen (1977), and Neale (1988). Another exclusively shallow marine/brackish water ostracod taxon Buntonia sp. has been recovered from sample JH21 from the Jhilmili intertrappean beds (Khosla et al., 2011). Furthermore, presence of holostean fish taxa such as Pycnodus sp. and Lepisosteus sp. also indicates fluvio-lacustrine environments of deposition for the Jhilmili intertrappean sediments (Sunil Bajpai, pers. comm.).
Foraminiferal as well as microfacies data (Keller et al., 2009a, b) point towards temporary or short term marine incursions into freshwater environments ensuing in shallow marine to brackish water conditions. Seasonal currents could have transported planktonic foraminifera from a nearby marine seaway. Absence of benthic foraminifera attests that marine conditions did not continue long enough at Jhilmili for benthics to invade the environments (Keller et al., 2009a, b). Most of the foraminifera recovered from the maceration techniques are not identifiable because many of them were mechanically compressed and deformed because of tectonic stress and pressure (Keller et al., 2002; Punekar et al., 2014). As a result they were diagenetically altered and have calcite overgrowths and are characterized by a recrystallized test and are difficult to identify. However, the presence of a cancellate spinose wall texture in numerous specimens corroborates the evidence that they are planktonic in nature (Figure 6, Keller et al., 2009a).

Figure 6 Planktonic foraminifera showing the cancellate spinose wall texture with calcite overgrowth of chambers; a) and b) Subbotina triloculinoides (Plummer, 1926). Photographs from Keller et al. (2009a).
The planktonic foraminiferal assemblage from Jhilmili intertrappeans (Table 1, 5m-5r) is not as diverse as that of the Rajahmundry intertrappean beds (southeastern coast of India). However, the presence of Guembelitria cretacea (Figure 5n, disaster opportunist) in the Jhilmili intertrappean beds, known universally in earliest Danian zones (P0 and Pla), indicates they continued to live in most stressful conditions especially during the late Maastrichtian along with other species (Abramovich and Keller, 2002; Pardo and Keller, 2008). Guembelitria cretacea occurred in shallow continental shelf environments and in areas of intense volcanism (Keller and Pardo, 2004). Guembelitria cretacea and Hedbergella sp. are considered as ecological generalists of varying environmental conditions and linked with warm humid climates, low salinity, temperature and low oxygen conditions (Boersma and Premoli-Silva, 1988; Keller, 1993; Malarkodi et al., 2010).
Apart from Guembelitria and Hedbergella, other earliest Danian species, such as Parvularugoglobigerina eugubina, P. extensa, Globoconusa daubjergensis and Woodringina hornerstownensis, also persisted in highly stressful conditions and turned over in great numbers or disappeared when circumstances changed after zone Pla (Malarkodi et al., 2010). The species Parasubbotina pseudobulloides (Figure 5p) contains a typical broad lip that covers the aperture and this may be due to the presence of muddy water in shallow environment conditions (Keller et al., 2009a, b).
The ostracod fauna is in common agreement with faunal survivorship in fresh water fluvial systems across the K-Pg boundary in the Deccan volcanic province. Ostracods are not crucial for age dating; in fact, as discussed above, the ostracod assemblages from Jhilmili in the uppermost Maastrichtian and lowermost Danian are similar, and their age is inferred by planktonic foraminifera. However, a few brackish water ostracods were dated as early Danian (P1a) (Sharma and Khosla, 2009; Keller et al., 2009a, b; Khosla et al., 2011). Taxonomically analogous ostracod assemblages have been described from Upper Cretaceous intertrappean sections across peninsular India (Whatley and Bajpai, 2005, 2006). The nonmarine ostracod assemblage recorded earlier from the intertrappean beds in Papro village, Lalitpur District, Uttar Pradesh, has been dated as Paleocene in age because the presence of pollen marker species (Lakiapollis ovatus, Dandotiaspora pseudoarticulata, D. dilata and Spinizonocolpites echinatus) also points to similarities in the taxonomic composition of ostracod faunas, though there are differences in the relative profusion of the diverse taxa (Singh and Kar, 2002; Sharma et al., 2008). A possible explanation is that the intertrappean freshwater ostracods were not considerably affected, at least qualitatively, by Deccan volcanic eruptions. Khosla et al. (2005, 2011) observed a strong resemblance of nonmarine ostracod taxa between the Upper Cretaceous (Maastrichtian) Lameta Formation, which lies below the Deccan volcanics and the Deccan intertrappean beds. This condition is evocative of other intertrappean freshwater organisms for instance molluscs (Hartman et al., 2007). The molluscan assemblage at the Papro intertrappeans consists of four taxa: Lymnaea, Physa and two species of Viviparus. These taxa can be compared to equally sized molluscs from Upper Cretaceous intertrappean beds of central India. The present ostracod data from Jhilmili (Madhya Pradesh) and Papro (Uttar Pradesh) support suggestions already made by Hartman et al. (2007) and Sharma and Khosla (2009). The fact that these taxa were able to repopulate during periods between basaltic flows in peninsular India imply that refugia were situated comparatively nearby. The intertrappean beds of Bombay region, has also yielded palynoassemblage of early Paleocene age and has been interpreted as showing no major floral declines (Cripps et al., 2005).
DISCUSSION
The seasonal offshore currents considered as short lived marine transgressive phase could have been the probable reason for bringing brackish water ostracods and planktonic foraminifera into the intertrappean beds of Jhilmilli, central India (Figure 1). This further develops provisional estuarine conditions leading to deposition of a marine biota (Keller et al., 2009a, b). Further evidence of marine transgression along the Narmada rift zone in stratigraphically older Bagh Beds in districts Dhar and Jhabua during late Cenomanian-early Turonian time was based on the presence of marine invertebrates, for example planktonic foraminiferas, ostracods, gastropods, ammonoids, bivalves, brachiopods, nannofossils and algae (Dassarma and Sinha, 1975; Jafar, 1982; Chiplonkar et al., 1977; Nayak, 2000; Keller et al., 2009a, b; Khosla and Verma, 2015). However, on the basis of sedimentological data of the Upper Cretaceous Lameta Formation it was proposed marine conditions in central India (Singh, 1981; Shukla and Srivastava, 2008). Other studies (Brookfield and Sahni, 1987; Tandon et al., 1995; Khosla, 2014) opposed this view by proposing a fresh water origin for these deposits.
Palaeobiogeographical implications
The ostracod fauna from the Indian infraand intertrappeans, including the present discussed locality Jhilmili, are highly endemic and many Indian taxa seem to have dispersed out from India, such as Mongolianella, Limnocythere, Cyclocypris and Eucypris to many parts of the Maastrichtian world (Whatley and Bajpai, 2006). Of the 26 taxa recorded so far from the Jhilmili intertrappean beds (Table 1), Paracypretta and Gomphocythere are exclusively endemic to peninsular India (Whatley and Bajpai, 2006). The genus Frambocythere is widely recorded from the Indian intertrappeans, but Frambocythere tumiensis was first recorded (outside India) from the Late Cretaceous of Spain (Helmdach, 1978). Other species of Frambocythere have been recorded from the Montian of Belgium (Tambareau, 1984) and Albian of Africa (Colin, 1993). Other genera such as limnocytherids Gomphocythere, Limnocythere and cyprids like Paracandona and Cypria, probably originated and evolved out from India (Whatley and Bajpai, 2006). The brackish water genus Neocyprideis has been earlier recorded from the Rajahmundry intertrappeans (Jain, 1978) and outside India the species has been described from assemblage IV of the upper Eocene beds of the Hampshire Basin, England (Keen, 1977). According to Whatley and Bajpai (2006) and Whatley (2012) the Upper Cretaceous ostracod species have nearly 98% endemism in India but genera like Limnocypridea, which has been widely recorded from the Maastrichtian of China and Mongolia, somehow were able to overcome competition and colonise India (Whatley and Bajpai, 2006). Thus as a whole, the Indian ostracod fauna has its own dominants like Mongolianella, Cypridopsis and Gomphocythere, and important endemics, such as Paracypretta (Whatley and Bajpai, 2006). The high species diversity of the Indian Maastrichtian freshwater ostracoda fauna provides evidence that around 88 Ma ago India was separated from Madagascar (Storey et al., 1995) and enjoyed protracted isolation during its northward drift through the Indian Ocean for 40 m.y. before colliding with the Asian landmass circa 50 Ma, carrying with it Gondwanan forms to Asia (Karanth, 2006). Therefore, the fauna from the Jhilmili intertrappeans and other localities of peninsular India supports the "Out of India" hypothesis (Whatley and Bajpai, 2006).
Revised age for Deccan intertrappeans and biostratigraphic implications for Jhilmili intertrappeans
The delineation of the K-Pg boundary and determining the age of Deccan traps and interbedded intertrappean sediments in central India was not possible previously because age diagnostic fossils were not known. For the last decade intertrappean sediments yielded a wealth of fossil data (Sahni and Khosla, 1994a, b; Loyal et al., 1996; Bajpai and Prasad, 2000; Khosla, 2001; Khosla and Sahni, 2000, 2003; Khosla et al., 2004, 2009; Whatley et al., 2002a, b; Whatley and Bajpai, 2005, 2006; Prasad et al., 2007a, b, 2010; Samant and Mohabey, 2009; Keller et al., 2008, 2009a-b-c, 2010a, b, 2011; Malarkodi et al., 2010; Bajpai et al., 2013). Biotic elements, i.e. fishes, frogs, crocodiles, turtles, dinosaurs, mammals, ostracods, charophytes and molluscs, point to a mainly freshwater, lacustrine depositional setting developed under semi-arid conditions for the intertrappean sediments of the District Dindori (Khosla et al., 2004; Prasad et al., 2007a, b, 2010), near Jabalpur (Mathur and Sharma, 1990). The intertrappean assemblages of Nagpur, Naskal, Mamoni, Kachchh and Gurmatkal (Bajpai and Prasad, 2000; Whatley and Bajpai 2000a, b; Whatley et al., 2002a, b; Vianey-Liaud et al., 2003) are also dominated by alike freshwater fauna and flora. The intertrappean beds show a diversity of lithotypes ranging from clays, claystones, marls, mudstones, siltstones and calcretes to limestone, which are strongly indicative of fluctuating lake levels (Khosla and Sahni, 2003). The freshwater biota is stratigraphically long ranging and is not useful in constraining the K-Pg boundary.
Early workers (Bande and Prakash, 1982; Bande et al., 1988) assigned a Paleocene-Eocene age to the Deccan intertrappean sediments of the Nagpur-Chhindwara region, interpreting a seashore palaeoenvironment based on marine algae (Solenopora and Peyssonnelia) and fossil palm fruits for example Cocos, Sonneratia and Nypa. The record of other fossils, for instance dinosaur eggshells, teeth and bones, fishes, frogs, turtles, crocodiles, ostracods and charophytes from the intertrappean localities of peninsular India other than Jhilmili, and the absence of Paleocene strata and fossils suggested a Maastrichtian age (Prasad and Cappetta, 1993; Loyal et al., 1996; Kar and Srinivasan, 1998; Bajpai and Prasad, 2000; Khosla and Sahni, 2000, 2003; Khosla et al., 2004, 2009; Whatley and Bajpai, 2005, 2006; Prasad et al., 2007a, b, 2010; Khosla, 2014; Khosla and Verma, 2015; Fernandez and Khosla, 2015; Khosla et al., 2015). The classic intertrappean beds exposed in the Chhindwara region especially in the Mohgaon Kalan area (Sahni and Rode, 1937; Kar and Srinivasan, 1998; Whatley et al., 2002b; Khosla and Nagori, 2007) are located about 5 km southeast of the Jhilmili section. The Mohgaon Kalan locality has a rich and diversified megaflora including palms and conifers suggesting estuarine and freshwater environments (Prakash, 1960; Kapgate, 2005). Apart from the flora, other Maastrichtian fauna include ostracods, charophytes, gastropods, dinosaur eggshells and pollen assemblages (Azolla cretacea, Aquilapollenites bengalensis and Gabonisporites vigourouxii (Kar and Srinivasan, 1998; Whatley et al., 2002a, b; Kapgate, 2005; Khosla and Nagori, 2007). Other intertrappean locality is exposed in the close vicinity of the Jhilmili area, in Singpur, which like Mohgaon Kalan contains a Maastrichtian palynoflora (Samant et al., 2008). Lithologically, Jhilmili intertrappean beds are virtually different from those of Mohgaon Kalan and Singpur localities, and do not contain any megaflora and palynofloral remains, as are present in the above mentioned localities. Therefore the intertrappean beds at Jhilmili show different age and palaeoenvironments. Based on the planktonic foraminiferal assemblage (Parvularugoglobigerina eugubina, P. extensa, Woodringina hornerstownensis, Eoglobigerina edita, E. eobulloides and Globoconusa daubjergensis, and rare Parasubbotina pseudobulloides, Praemurica taurica, Subbotina triloculinoides, Globanomalina compressa and Globigerina (E.) pentagona) an early Danian zone P1a age has been assigned to the intertrappeans of Jhilmili (Keller et al., 2009a, b). The occurrence of P. eugubina in the Jhilmilli intertrappeans shows that the deposition of the foraminifera bearing horizon took place about 100-150 ka later than the K-Pg boundary (Keller et al., 2009a, b). The upper part of the intertrappean sediments consisting of green and red shales and characterized by paleosols and deposited in a palustrine environment were considered the upper part of zone P1a (2) whereas the lower part of the intertrappean sediments which are 6 m thick and lithologically consists of paleosols (red shale), were possibly deposited during the early Danian P0-P1a (1) interval (Keller et al., 2009a, b). These intertrappean beds are correlative to the shallow marine intertrappean beds exposed in the Rajahmundry quarries, which also has yielded early Danian zone planktonic foraminiferal assemblages between the lower and upper basalt traps of C29r and C29n magnetic polarity zones (Keller et al., 2008). Keller et al. (2009a, b) reported magnetostratigraphic data for Jhilmili that demonstrate that the occurrence of the K-Pg boundary flanked by the lower traps is in 29r and upper trap is located at C29r/C29n, which are correlative to the upper Ambenali and lower Mahabaleshwar formations, correspondingly (Jay and Widdowson, 2008; Keller et al., 2008). Therefore the lower volcanic flows at Jhilmili put the K-Pg boundary close to the end of the main Deccan volcanic eruptions, as indicated by early Danian planktonic foraminiferal assemblages overlying the last part of lower traps (Keller et al., 2009a).
CONCLUSIONS
Diverse ostracod and planktonic foraminifera assemblages (mixed fauna) have been recorded earlier by Keller at al. (2009a, b) from the Jhilmili intertrappeans, central India. The presence of planktonic foraminifera modifies the age from earlier assigned Maastrichtian to early Danian. The ostracod assemblage recovered from unit 3 (60 cm thick) indicates the presence of a freshwater lake or pond, dominated by two species (Limnocythere deccanensis and Zonocypris viriensis). Active swimmers include Mongolianella, Paracypretta and Zonocypris, whereas the bulk of the ostracod assemblage is dominated by poor swimmers (Darwinula and cytheraceans Gomphocythere, Frambocythere and Limnocythere). Two brackish water ostracod species (Neocyprideis raoi and Buntonia sp.) and planktonic foraminifera recovered from sample numbers (JH21 to JH26) indicates brackish-marine environment. The assemblage further reveals the existence of a marine seaway in central India during the Maastrichtian to early Paleocene along the Narmada and Tapti rift zones (Keller et al., 2009a, b). Palaeoenvironmentally, deposition at Jhilmili occurred in predominantly terrestrial semi-humid to arid environments, followed by a short aquatic interval (<60 ka) of fresh water ponds and lakes, which led to shallow coastal marine/ estuarine conditions with brackish ostracods and early Danian P1a planktonic foraminifera.
Palaeobiogeographically, the ostracod fauna from the Jhilmili intertrappeans is somewhat endemic to India. The Jhilmili fauna includes endemic forms such as Gomphocythere and Paracypretta and two of its dominant forms like Cypridopsis and Mongolianella. Most of the Jhilmili ostracod taxa, such as Cypridopsis, Eucypris, Limnocythere and Mongolianella appear to have disseminated out from India to sundry parts of the Upper Cretaceous (Maastrichtian) world.

