











 nueva página del texto (beta)
nueva página del texto (beta) Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkINTRODUCTION
Pliosaurid plesiosaurs form an extinct group of highly specialized marine sauropterygian reptiles. Although the first scientific reports of pliosaurids dates backs to the early 19th century, based on fossils recovered from English Kimmeridgian Clay Formation (Conybeare, 1821, 1822); the careful study of a rather scattered fossil record of these limbs reptiles shows them as a cosmopolitan group that dominated the oceanic large-bodied predator niche from the Callovian (Middle Jurassic) Turonian (Late Cretaceous) (Ketchum and Benson, 2011a, b; Schumacher et al., 2013; among others). Recent phylogenetic analyses recovered Pliosauridae as a well supported group including 12 genera, 6 of which lived during the Middle-Late Jurassic Pliosaurus Owen, 1840; Peloneustes Lydekker, 1889; Liopleurodon Sauvage, 1873; Simolestes Andrews, 1909; Gallardosaurus Gasparini, 2009; and Marmornectes Ketchum and Benson, 2011a (also see O'Keefe, 2002; Benson et al., 2013; Vincent et al., 2013; Benson and Druckenmiller, 2014; Foffa et al., 2014).
Today, our knowledge of Middle-Late Jurassic pliosaurs is mainly based on northern hemisphere records, principally from the Oxford and Kimmeridge Clay formations of England and France (Sauvage, 1873; Andrews, 1913; Tarlo, 1960; Benson et al., 2013; among others), and well as from the Agardhfjellet Formation of Norway (Knutsen et al., 2012). Another boreal Jurassic pliosaurid is Megalneusaurus rex Knight, 1898, collected in Kimmeridgian marine deposits in the Pacific domain in the northwestern (Alaska) and western United States region (Wyoming) (Weems and Blodgett, 1996; and Wahl et al., 2007a, b); however, additional specimens are needed to define its relationships within Pliosauridae. Contrasting with the abundant northern hemisphere records, Southern hemisphere pliosaurids are scarce and restricted to the Early and Middle Jurassic of Australia (Kear, 2012) and the Middle and Late Jurassic of Argentina (Gasparini, 1997; O'Keefe, 2002; Gasparini and O ́Gorman, 2014).
In Mexico, the fossil record of pliosaurids has been extremely scarce, fragmentary, and taxonomically confusing. Wieland (1910) named the first Mexican plesiosaur as Plesiosaurus (Polypticodon) mexicanus, which is based on an isolated fragment of rostrum with teeth. Unfortunately, this author did not properly record the site of origin of this fossil or the evidence that led him to suggest its Neocomian age. Later, Buchy (2008) analyzed a cast of this specimen and identified it as a metriorhynchine crocodyliform, whose possible age is Late Jurassic. The direct examination of the Wieland ́s specimen carried out by one of the authors (JBL) confirms the identification. The second record of Mexican pliosaurs was published by Ferrusquía-Villafranca and Comas (1988), who describe few vertebral and jaw remains from supposedly Neocomian age strata exposed near Papalutla town, Huajuapan de León Municipality, State of Oaxaca. These authors conclude that these fossils represent a new genus related to Pliosaurus. Unfortunately, this proposal has not been due the lack of additional and the apparent loss of these fossils (Gracía-Barrera, 2014 pers. comm.). Other copious remains of pliosaurids have been recently recovered in northern Mexico, in numerous localities along the States of Coahuila and Nuevo León, where the marine sediments of La Caja and La Casita formations are exposed. After the review of these fossils, including a large specimen that could reach 15 m in length and commonly named as the Monster of Aramberri, the conclusion is that all pliosaurs found in northern Mexico are indeterminate members of the family Pliosauridae (Buchy, 2007; Buchy et al., 2003, 2006a-b-c).
Since 2008, as part of a research program Mesozoic fishes of Mexico, sponsored by the Universidad Nacional Autónoma de México (UNAM), have been conducted fieldworks in different Mesozoic sedimentary outcrops in this country. A significant result of this program was the discovery Yosobé, near Tlaxiaco, Oaxaca, Mexico (Figure 1).

This site, a diverse vertebrate assemblage has been recovered from the Kimmeridgian-Tithonian sequence of shale and limestone belonging to the informally named Sabinal formation (Alvarado-Ortega et al., 2014; Barrientos-Lara, 2013), which is part of the Jurassic-Cretaceous sedimentary deposits of the Tlaxiaco Basin (Meneses-Rocha et al., 1994). The fossil assemblage of Yosobé comprises numerous invertebrates (ammonites and bivalves, among others), carbonized wood, and vertebrates including fishes and marine reptiles. Among these fossils, the pliosaurs are represented by two specimens. The aims of this manuscript are to provide a detailed description of the Late Jurassic pliosaurid remains recovered in Yosobé and a review of their biogeographical implications.
MATERIAL AND METHODS
The two specimens described in this manuscript are deposited in the Colección Nacional de Paleontología, which is housed in the Museo María del Carmen Perrilliat, Instituto de Geología, UNAM. The Yobosé site is recorded as IGM-locality 3871 in the Catalog of Mexican localities of this collection. These fossils were prepared mechanically using fine air-scribe tools and dental needles, and they were hardened with plexygum. The institutional acronyms used in this manuscript are: IGM, Colección Nacional de Paleontología, Instituto de Geología, UNAM; BHN, Musée d ́Histoire Natrelle, Boulogne-sur-Mer, France.
SYSTEMATIC PALEONTOLOGY
Superorder Sauropterygia Owen, 1860
Order Plesiosauria Blainville, 1835
Family Pliosauridae Seeley, 1874
Unranked clade Thalassophonea Benson and Druckenmiller, 2014
Liopleurodon Sauvage, 1873.
Type species. Liopleurodon ferox Savage, 1873. BHN 3R 197; A single tooth from Wast, near Boulogne-sur-Mer, France.
Remarks. Although four species have been named within this genus (see Bardet, 1995); Noè (2001: 27-29 and 31-32) regarded Liopleurodon as a monospecific genus and L. ferox being its type species. The other species once referred to Liopleurodon, L. rossicus (Novozhilov, 1948) and L. macromerus (Phillips, 1871), today assigned to Pliosaurus (Tarlo, 1960; Halstead, 1971; Bardet, 1995; Noè et al., 2004; Knutsen, 2012; Benson et al., 2013). The species Liopleurodon ferox has been improperly also named as Pliosaurus ferox (Lydekker, 1888; Andrews, 1897; Delair, 1959; among others). Other possible synonyms of Liopleurodon ferox include nominal species as Ischyrodon meriani (von Meyer, 1838), Thaumatosaurus oolithicus (von Meyer, 1841), Pliosaurus giganteus (Conybeare, 1824), and Pliosaurus pachydeirus (Seeley, 1869) that are useless because these were erected on non-diagnostic and insufficiently characterized material (Brown, 1981; Noè, 2001).
Liopleurodon sp.
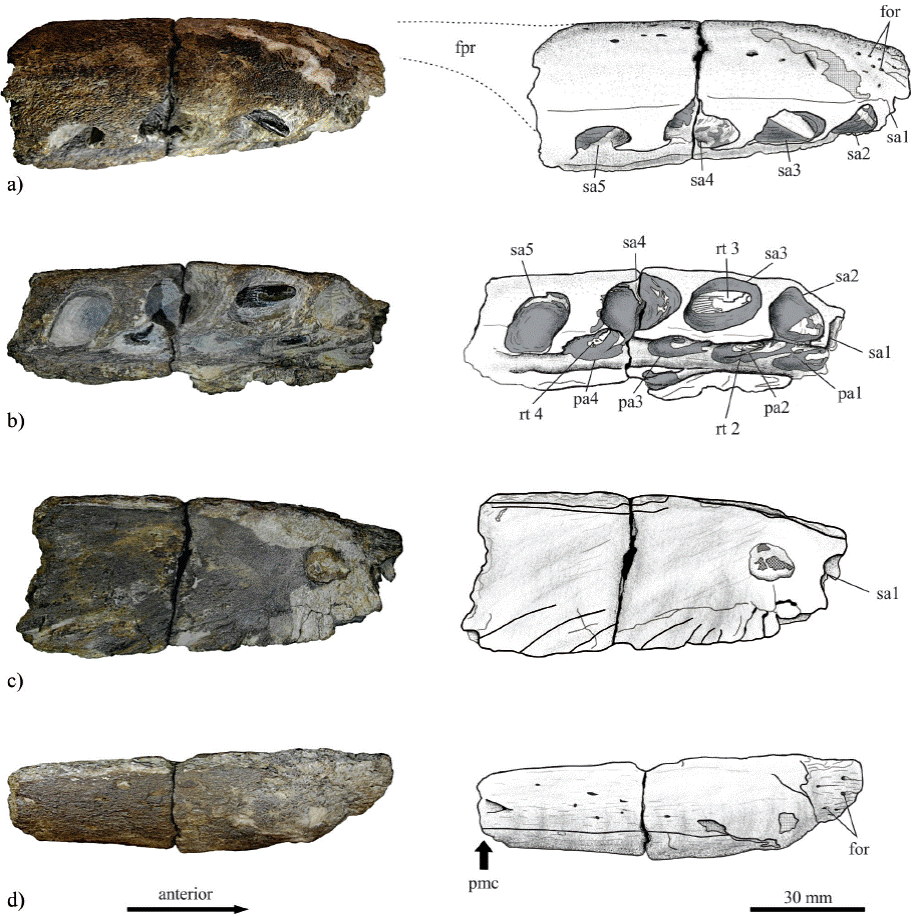
Figure 2 Photograph and line drawings of IGM 4545, fragment of right premaxilla of Liopleurodon sp. from the Kimmeridgian sediments of Yosobé , Oaxaca, southern Mexico. a) Lateral external view; b) ventral view; c) lateral medial view; d) dorsal view. Abbreviations: for, foraminae; pmc, premaxilla constriction; fpr, premaxilla facial process; rt 1-4, replacement tooth; pa 1-5, primary alveolus; sa 1-4 secondary alveolus.
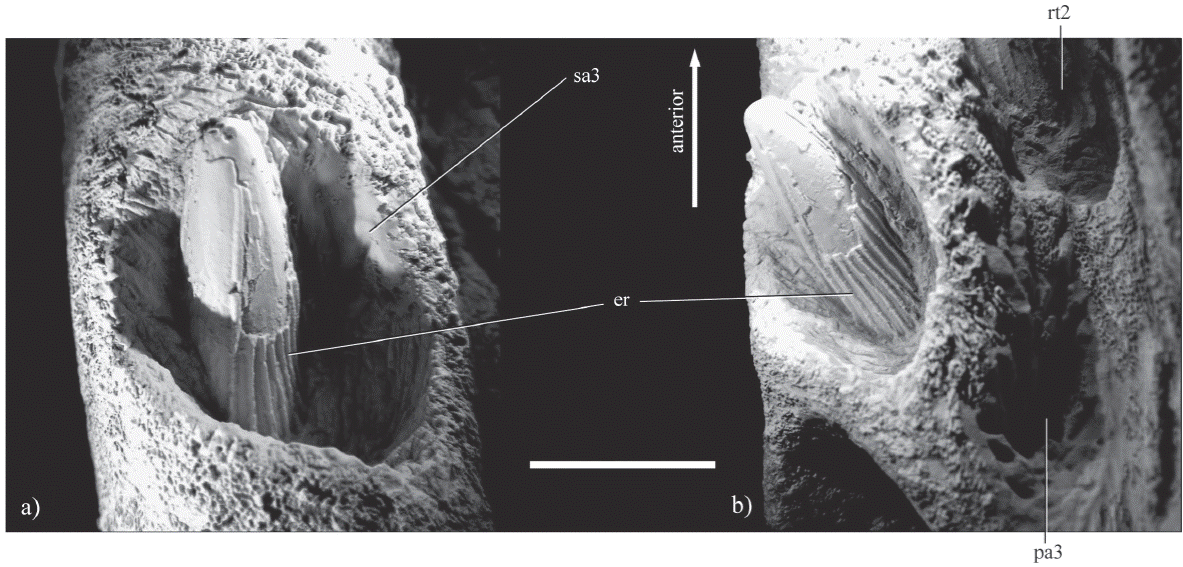
Figure 3 Replecement tooth in the functional alveolus 3 of IGM 4545, fragment of right premaxilla of Liopleurodon sp. from the Kimmeridgian sediments of Yosobé , Oaxaca, southern Mexico. shows that the enamel of near the crown tip was lost partially (the specimen is contrasted with ammonium chloride). a) lateraldorsal view; b) dorsal view. Abbreviations: er, enamel ridges; pa 3, primary alveoli 3; rt 2, replacement tooth 2; sa 3 secondary alveoli 3.
Table 1 Measurements of the teeth and alveolus of IGM 4545, a fragment of the right premaxilla of Liopleurodon sp. from Yosobé, Oaxaca, Mexico. All measurements are given in millimeters; +, means at least.

Referred material. IGM 4545, fragment of a right premaxilla divided into parts and preserving three replacement teeth.
locality and horizon. The shale layer L4-L5 of the Kimmeridgian sequence belonging to Sabinal formation at Yosobé, near Tlaxiaco town, Oaxaca State, Mexico (see Alvarado-Ortega et al., 2014). Description. Specimen IGM 4545 is a fragment of the anterior end of the right premaxilla. The shape of this bony fragment can be crudely described as similar to a quarter of a cylinder cut lengthwise. This bone is not preserved entire, its posterior end is broken and lacks the long narrow premaxilla facial process; however, its anterior stout and tooth-bearing section is well-preserved.
This premaxilla fragment, in lateral external face is a long roughly rectangular structure (130 mm long, 53 mm in height, and 32 mm width), in which the anterior end is tapered forming a rounded edge. In this view, the smooth and almost flat symphyseal surface is exposed showing some dorsal and ventral ridges. The ventral ridges are somewhat wavy; here the smaller undulations were probably interdigitated with similar undulations of the left premaxilla to make up a strong suture. The large undulation in the posterior area probably represents part of the space occupied by the vomer. The lateral external view of this bone shows a convex surface with small foraminae that increase in size and number anteriorly.
The ventral view of this bony fragment corresponds to the alveolar surface. Here the alveoli form two rows; the primary or labial row is located alongside the extrenal edge including the larger alveoli whereas the secondary or lingual row, near the symphysial edge, includes small alveoli interspersed with larger alveoli (Table 1). The labial row is composed of at least five functional alveoli, which are large oval, longer than wide, and placed so close to each other that practically occupy the entire ventral surface. In contrast, the internal or lingual row includes four replacement alveoli that open into a longitudinal canal near to the symphysis. The axes of all alveoli are projected laterally and forward suggesting that the teeth were slightly procumbent and anteriorly directed.
The tooth of the third functional alveolus is well preserved and its crown is almost totally exposed reaching the length 23 mm. This crown clearly is conical, straight, and circular in cross section. Although two teeth are preserved in replacement alveoli two and four, these are not fully developed and are embedded in the rock only part of their crowns are exposed. The enamel on the crown surface is heavily ornamented in the lingual surface with longitudinal ridges spaced uniformly. These ridges are project continuously from the crown base to its apex; also, these are more numerous and regular on the lingual surface that in the labial surface, where the ridges are most widely spaced and unevenly distributed.
Remarks. At the end of the last century, Carpenter (1996, 1999) review the family Pliosauridae, then restricted to short-necked Middle-Late Jurassic plesiosaurians species, suggesting that pliosaurids differ from all other plesiosaurs by the heavy or thick enamel ridges on the teeth (also present in the members of the subfamily Brachaucheninae today included into Pliosauridae). After Carpenter ́s works the taxonomical diversity of Pliosauridae has been significantly enriched and reordered (e.g. Benson et al., 2013; Benson and Drunkenmiller, 2014; among others). If the Carpenter ́s observations are considered, IGM 4545 could be recognized an unquestionable representative of the family Pliosauridae; however, today the thick enamel ridges on teeth are not part of the last diagnosis of this family , as it was described by Ketchum and Benson (2010). Among Pliosauridae, only Liopleurodon and Simolestes share a group of characteristics that includes the presence of heavy (= coarse, thick) enamel ridges on dental crowns, a transverse constriction in the rostrum at the level of premaxilla-maxilla suture, and five conical and robust functional teeth in the premaxilla (see Druckenmiller and Russell, 2008; Benson and Druckenmiller, 2014; and O'Keefe, 2001). In the present context, despite the incompleteness of IGM 4545, this specimen is certainly included into the family because it shares the combination of characteris of two Liopleurodon and Simolestes.
Among middle-Late Jurassic of pliosaurids, Pliosaurus and Gallardosaurus have trihedral teeth with two carinae separating the lingual and labial tooth faces (Knutsen, 2012; Gasparini, 2009). In teeth of IGM 4545, as in Simolestes and Liopleurodon, the enamel ridges runs almost continuously along the crown, from the base to apex (Tarlo 1960; Noè, 2001); however, the enamel ridges in Simolestes are comparatively less developed than those of Liopleurodon and IGM 4545. In Peloneustes and Marmornectes these ridges are present only in the basal half or three-quarters of the height of the crown (Ketchum and Benson, 2011a, b).
Liopleurodon and Simolestes clearly differ to each other if the shape of the premaxilla is considered (see Noè, 2001, vol. II, figs. 87-88, 94-95, 98-99, 108-111, and 130). In Simolestes the premaxilla is arrow shaped in dorsal view; it has a deep transverse constriction at the level of the premaxilla-maxilla suture, and its labial and symphysial edges are angled to each other. In Simolestes, the premaxilla is also twice wider than high. In contrast, the premaxilla of Liopleurodon is more rectangular, where the labial and symphyseal edges run parallel to each other almost all along and the transverse constriction is rather shallow. In Liopleurodon, the premaxilla is also higher than wide. IGM 4545 shares the premaxilla characters of Liopleurodon supporting its inclusion in this genus.
Although the specimen IGM 4545 shows the stout continuous enamel ridges that constitute the most remarkable features in the diagnosis of the monospecific genus Liopelurodon, which is also applicable to Liopleurodon ferox; currently, this species is restricted to the Callovian strata of Europe. Therefore, the specific determination of the Kimmeridgian Liopleurodon specimen from Tlaxiaco will require additional material and morphological data.
Genus and species indet.

Figure 4 Photograph of selected bones of IGM 4546, an indeterminate Pliosauridae specimen from the Kimmeridgian sediments of Yosobé , Oaxaca, southern Mexico. a) Outline draw of a pliosaurid showing the position of bony elements illustrated here; b-e) a dorsal centrum (IGM 4546, element 11) in anterior, lateral, dorsal and ventral view respectively; f-i) a caudal centrum (IGM 4546, element 7) in anterior, lateral, dorsal and ventral view respectively; j) caudal centrum in lateral view (IGM 4546, elements 3-7) highlighting the facet for rib; k-n) neural arch (IGM 4546, element 17) in posterior, lateral, anterior and ventral view respectively. Abbreviations: fna, facet for neural arch; for, foramen; ribf, rib facet; nc, neural canal; ns, neural spine; rib, rib remains; st, shark tooth.
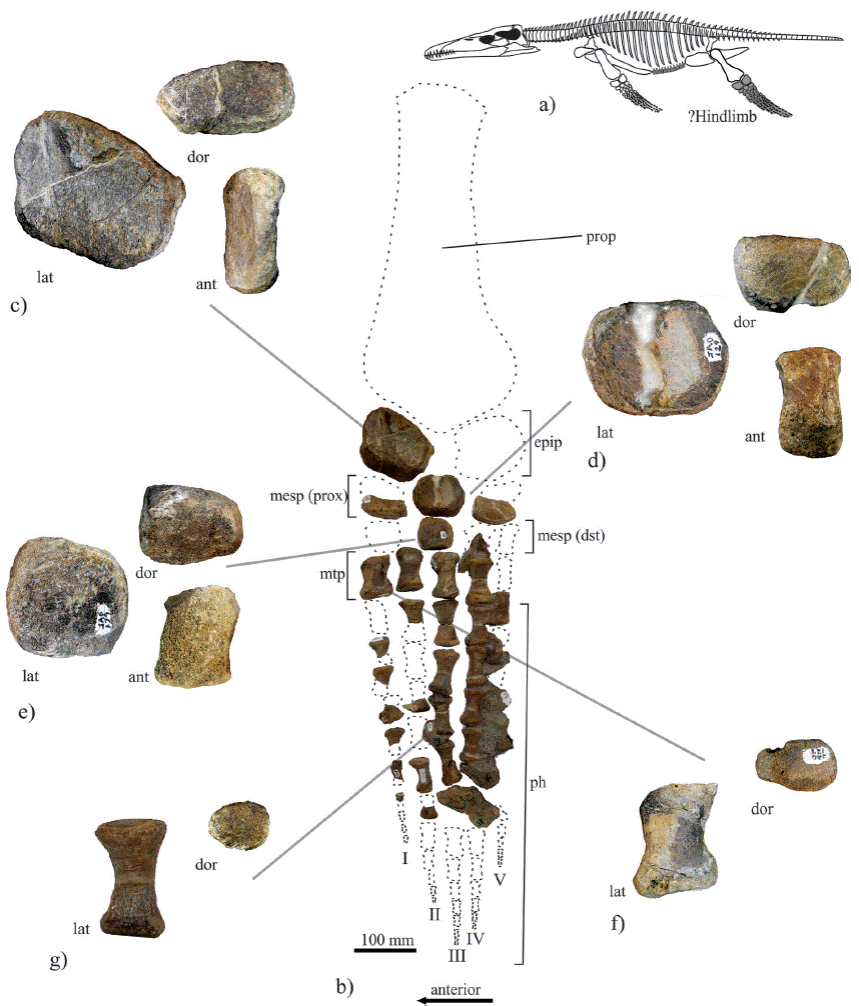
Figure 5 Photograph s of selected bones of the limb of IGM 4546, an indeterminate Pliosauridae specimen from the Kimmeridgian sediments of Yosobé , Oaxaca, southern Mexico. a) Outline draw of a complete pliosaurid showing the position of the bone elements illustrated here; b) bones of the limb showing their natural positions; c) epipodial (IGM 4546, element 19); d) Proximal medial mesopodial (IGM 4546, element 20); e) Distal mesopodial (IGM 4546, element 23); f) metapodial I (IGM 4546, element 24); g) phalanx 3-III (IGM 4546, element 37). Abbreviations: ant, anterios view; dor, dorsal view; dst, distal; epip, epipodial; I-V, digits; lat, lateral view; mesp, mesopodial; metp, metapodial; ph, phalanges; prop, propodial; prox, proximal.
Table 2 Measurements and proportion of caudal and dorsal elements of the vertebral column of the specimen IGM 4546, an indeterminate Pliosauridae from Yosobé, Oaxaca, Mexico. Means do not include missing (-) or uncertain (?) measurements.

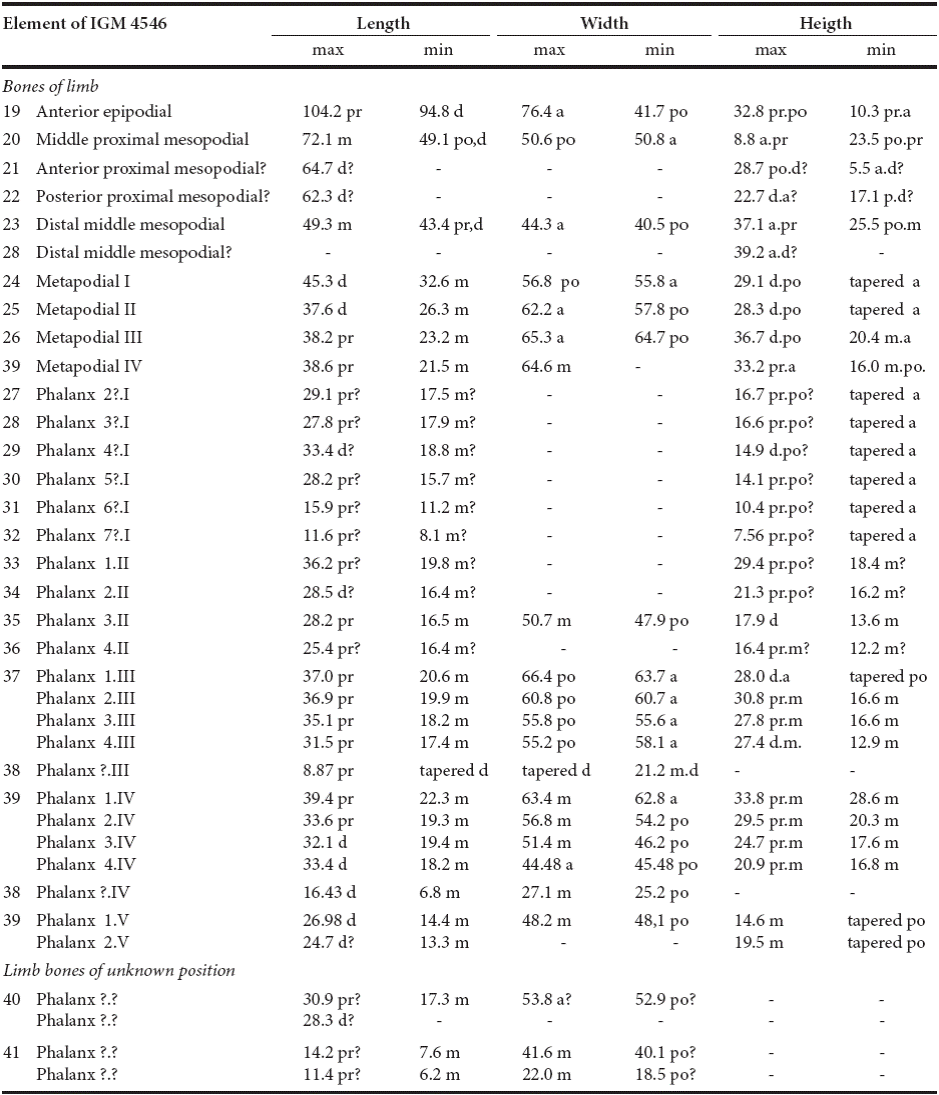
Table 3 Measurements and proportion of elements of IGM 4546 that represents the limb (expressed in millimeters). Arabic numerals show the position of the phalanges in a proximal-distal order; roman numerals indicate the digit to which they belong; anterior and posterior refers to the position of the element in the limb.
Abbreviations to show the position of the maximum and minimum measurements are indicated as: a, anterior border; d, distal border; po, posterior border; pr, proximal border; m, middle.
Referred material. IGM 4546. This specimen consists of 16 vertebrae (nine dorsals and seven caudals (IGM 4546, elements 8-16 and 1-7, respectively); a neural arch almost complete and an articular pedicel of a neural arch (IGM 4546, elements 17 and 18 respectively); a epipodial, tree mesopodial proximals, two distal mesopodials, four metapodial elements, and twenty two phalanges of the same limb (11 of them articulated to each other) (IGM 4546, elements 19-39). There are also four phalanges of unknown position (preserved as part of the elements 40 and 41 of IGM 4546), plus several rib fragments and other unidentified bones. Here, all these skeletal elements are considered as belonging of the same individual because they were collected in the same spot with some of them in life position. A shark tooth of Planohybodus Rees and Underwood, 2008, was found associated to one of the vertebrae of this specimen (Figure 4f) (Alvarado-Ortega et al., 2014).
Ocurrence. Shale layer M-L1 of the Kimmeridgian bituminous shales belonging to Sabinal formation at Yosobé, Oaxaca, southern Mexico (see Alvarado-Ortega et al., 2014).
Description. Axial skeleton. Vertebrae are thick and short cylindrical structures. Their surfaces are strongly weathered and therefore the occurrence or absence of a ventral keel in these vertebrae cannot be confirmed. Large nutritive foramina are present on the lateral and ventral surfaces of the vertebrae. The inter-vertebral articular surfaces are weakly concave. There elongate oval facets for the attachment of the neural arch the dorsal surface of each centra. These facets frame the floor of the neural canal, is flat and hour-glass shaped. None of the centra are fused with their respective neural arches suggesting that IGM 4546 is a juvenile individual.
Table 2 summarizes the measures of vertebrae preserved of IGM 4546 and their basic proportions. The width and height of these bones range between 60 and 90 mm; these are approximately equal in length with a mean value of 45 mm. In IGM 4546, the dorsal centra are recognized for absence of rib facets. The caudal centra are proportionally wider than the dorsals and their inter-vertebral surfaces are ovate in outline. Additionally, caudal centra bear a single oval concave rib facet on each lateral surface; this is located on the top of a small lateral projection (Figure 4f and 4g). The anatomical position of these vertabrae within the caudal series cannot be confidently determined; however, five of these (IGM 4546, elements 3-7 in Figure 4j) have the rib facets placed on the upper half of the centra height, whereas this facet is placed very close to its dorsal edge in one (IGM 4546, element 7) (Figure 4g). The position of the rib facet suggests the caudal position of these vertebrae, with the latter being the closest to the sacral region.
There are two neural arches IGM 4546, one almost compete (element 17 in Figure 4k-4n) and other that only comprise the ventral processes that attach with the centra (element 18 of IGM 4546, not figured). In anterior view, the first of these arches shows a triangular shape, in which the neural cavity comprises a quarter of the height of the neural arch, and the rest of this bone forms a thin neural spine. In lateral view, this bone is sigmoid because the neural spine is tilted backward and has the anterior and posterior edges slightly curved. Laterally, the neural cavity is formed by two stout ventral pedicels that are ventrally expanded. In ventral view and at the end of these pedicels, the articular facet triangular with rounded edges. The lack of zygapophyses of the neural arches preserved, suggests that this elements correspond to the posterior caudal area.
Limb. The 20 element (IGM 4546, elements 9 to 39) of the limb bones described here, are considered as part of the same limb because they were collected together (Figure 5). The position of the some phalanges and other bones preserved in two rock fragments (IGM 4546, elements 40 and 41) is unknown. Table 2 summarizes the measurements of all these bones.
In dorsal and ventral view the epipodial (IGM 4546, element 19) is stout and roughly trapezoid in shape with a curved convex anterior border (Figure 5c). Is 1.5 times longer than wide, all its edges are almost straight but of unequal size; the proximal edge is the longer edges, it is followed by the distal, then the anterior is wider than the posterior, and the fifth edge that is located in the posterior distal end is the smaller. Although the ventral and dorsal surfaces of this bone are smooth; each of them is almost entirely occupied by a wide and shallow groove that is extended from anterior to posterior edge. The height of this bone is not uniform. The proximal and probably the distal articular surfaces are arrow shaped, whereas the posterior and posterior distal articular surfaces are rectangular. In this bone all the articular facets are rough and slightly convex.
In dorsal and ventral view, the proximal mesopodial located in the middle section is hexagonal shaped (IGM 4546, element 20), about 1.5 times longer than wide (Figure 5d). As the epipodial, the ventral and dorsal surfaces of this bone are smooth and concave. All its articular facets are rough and slightly convex (Figure 5d). However, unlike the epipodial, in the proximal middle mesopodial the height is 1.3 to three times higher in the anterior edge than in the posterior. Although the proximal and distal articular facets of this bone are trapezoids and similar in size; its transversal facets are different in shape and size. The anterior proximal transverse facet that articulates with the epipodial is rectangular and about 1.5 times higher than wide. The anterior distal transverse facet is square. The posterior proximal transverse facet is rectangular, about 1.2 to 1.3 times higher than wide, and with the dorsal edge strongly concave.
Two very poorly preserved bones collected near to the epipodial and proximal middle mesopodial are here identified as possible remains of the laterals proximal mesopodial element (IGM 4546, elements 21 and 22, respectively). Although, a large part of the proximal end is lost in these bones; Is estimated that these bones were rectangular shape in dorsal and ventral view, in this face is observe surfaces are smooth and concave.
A complete stout middle distal mesopodial is preserved (IGM 4546, element 23). A small bone fragment that probably represents another stout middle distal mesopodial of the same limb is preserved at the proximal end of rock that bears a series of metapodial and six phalanges in life position (IGM 4546, element 39). The 3-D shape of the complete middle distal mesopodial is almost cubic. In dorsal and ventral view, this bone is almost square, slightly longer than wide, and the anterior edge is slightly wider than the posterior edge. The ventral and dorsal surfaces of this bone are smooth and concave. Although the height of this bone tapers posteriorly, its anterior height is about 1.25 times the height of its rear. All the articular facets of this bone are coarse and slightly convex. Its anterior articular facet, the larger, is rectangular, slightly wider than high, and its ventral and dorsal borders are notably concave. The posterior facet also is rectangular and its ventral and dorsal borders are concave, but this facet is about 1.6 times wider than high and the ventral border is notably less curved. The proximal and distal facets of this bone are trapezoid shape and similar in size; in these the length is 1.125 times longer than the anterior height and 1.5 times the posterior height.
Other bones of this limb partially recovered include elements of the four metapodial element and other elements of the five digits (Figure 5b). All these bones show the typical "I"shape, in that the proximal and distal end form articular facet that are greatly expanded in relation to the central region of these bones. The four metapodial bones (IGM 4546, elements 24 to 26, and the second proximal bone in IGM 4546, element 39) were recovered. These are the most robust and long elements in the digits. The metapodial I tapers anteriorly. The metapodial I to IV are arranged in parallel to each other forming a line.
There are 26 phalanges of the limb preserved (IGM 4546, elements 27 to 39, in Figure 5b and Table 3). This series includes fragments of six thin phalanges that are tapered anteriorly and probably belong to digit I. This also includes one complete and three fragments of the phalanges of digit II, five complete phalanges belonging to digit III, as well as other of digit IV. Digit V only is represented by two complete phalanges. The size of the phalanges decreases in proximal distal order; those of digits I to IV are also arranged in lines and those belonging to the digit V are misaligned. The phalanges of digits I and V are tapered anteriorly and posteriorly, respectibvely. The distal end of the most distal phalanx of digit III identified here (in Table 4) is rounded.
Remarks. Vertebral morphology, including the proportions and the presence of paired or unpaired nutritive foramina on the ventral or lateral surfaces of caudal and dorsal centra, as well as the morphology of the limb guarantee the identification of the IGM 4546 as a Plesiosauria. Probability the limb described above of this specimen represents a hind limb, given its filed position near to the caudal vertebrae; however, there is no additional evidences to support this possibility. Therefore, it is not possible to use the features of this limb to reach an accurate taxonomical identification. Nevertheless, IGM 4546 shares with all the Jurassic Pliosauridae genera the proportion of the vertebrae; these areapproximately two times higher than long (Buchy et al., 2003). Recently, Benson and Druckenmiller (2014) defined the Thalassophonea clade within Pliosauridae; however, characters supporting this clade are not preserved in the IGM 4546 and therefore this material is identified here just to the family level, as Pliosauridae.
DISCUSSION AND CONCLUSION
Four European species (Liopleurodon ferox Sauvage, 1873; Pliosaurus pachideirus Lydekker, 1888; Stretosaurus macromerus Tarlo, 1959; and Pliosaurus rossicus Novozhilov, 1948) have been included within the genus Liopleurodon; however, today this is considered a monospecific genus represented by the species L. ferox. According Noè (2001: 31-32), the remains of L. ferox has been recorded from Bathonian-Tithonian marine deposits in France, England, Switzerland, and Germany, as well as probably from Poland and Russia; nonetheless, this author and Benson et al. (2013: 32) recognized that those fossils with indubitable diagnostic characters of this species are restricted to the Callovian, mainly to the Oxford Clay Formation of England. The morphology of premaxilla fragment and teeth preserved in IGM 4545 are consistent with those expressed in the diagnosis of Liopleurodon (Noè 2001); also the combination of dental features in IGM 4545 contrasts with those of all other Jurassic pliosaurid genera (see Systematic Paleontology). Considering the age and geographical distance between the Callovian of the Oxford Clay Formation, in Europe, and the Kimmeridgian deposits of the Sabinal formation, near Tlaxiaco, Mexico; there is a possibility that IGM 4545 could represents a new species. The incompleteness of IGM 4545 precludes the execution of a suitable morphological comparative exercise and the identification of this possible new species; hence, the authors prefer to wait for the discovery of more fossils in the Sabinal formation that allow us to determine the species of these Mexican Liopleurodon representatives. In any case, the identification of this specimen has significant implications because this finding extends the stratigraphic range of Liopleurodon into the Late Jurassic along the entire Tethys Sea domain, from Europe to Mexico.
Pliosaurids from the Sabinal formation are significant from a paleobiogeographical point of view. During the Jurassic one important paleogeographic process was the breakup of Pangea landmass in two large continents, Gondwana and Laurasia, whose final separation at the west resulted in a narrow and tight sea corridor, the Hispanic Corridor (Hallam, 1977). Since the Early Jurassic, this marine pathway connected the western Tethys with the eastern Pacific acting as an important dispersion route for marine benthonic organisms from both sides, which had to adjust to the consequent reorganization of marine flows patterns at global level triggered by this connection (Riccardi, 1991; Parrish, 1992; Damborenea et al., 2013). It has been hypothesized that the Hispanic Corridor was a barrier for most pelagic invertebrate faunas until Middle or even Late Jurassic (Damborenea and Manceñido, 1979; Boomer and Ballent,1996; Damborenea, 2000; Aberhan, 2001); contrary, records of Jurassic marine reptiles indicate the use of the Hispanic Corridor as a dispersal route for marine reptiles during Middle and Late Jurassic, with no evidence of selective barriers for pelagic versus costal forms. The use of this sea way by marine reptiles have been proposed since the 90 ́s based on the similarities between Middle and Late Jurassic western Tethys marine reptiles assemblages with those of the eastern Pacific, mainly the Late Jurassic assemblages of the Neuquén Basin in northwest Patagonia (e.g. Gasparini, 1992; Gasparini and Fernández, 1996, 1997, Gasparini and O ́Gorman, 2014). The first direct evidences of marine reptiles within the Hispanic Corridor have been yield by Oxfordian marine reptiles assemblage of the Jagua Formation of Cuba including off-shore predators such as ophthalmosaurid ichthyosaurs, metriorhynchid crocodylifomrs and cryptoclidid and pliosaurid plesiosaurs up to more costal forms such as pleurodiran turtles (Fernández and Iturralde-Vinent, 2000; Gasparini and Iturralde-Vinent, 2001; De la Fuente and Iturralde-Vinent, 2001; Gasparini et al., 2002; Gasparini, 2009). To these evidences must be added latter discoveries of Late Jurassic (Tithonian) marine assemblages of north-east México (Buchy et al., 2003; Buchy, 2007).
In the particular case of pliosaurid recovered in the Hispanic Corridor area, previously represented for Gallardosaurus iturraldei from the Oxfordian of Cuba and the Pliosauridae indeterminate from the Late Kimmeridgian-Tithonian strata of La Casita Formation, northeastern Mexico, both Kimmeridgian specimens described here from the Sabinal formation at Tlaxiaco basin, complement and continue the geographical and temporal distribution known of the thalassophonean non-brachauchenine pliosaurids up to Kimmeridgian. Today, these pliosaurids are known in the Thitonian deposits of the Vaca Muerta Formation, Neuquen basin, Argentina Gasparini and O ́Gorman, 2014), the Kimmeridgian Clay Formation exposed in England and France (Ketchum and Benson, 2011a,b), the Volgian strata in Russia (Novozhilov, 1964), the Svalbard Archipelago, Norway (Knutsen, 2012), as well as other localities in Germany and Switzerland (Noè, 2001) and deposits in the southern tip of North America (Figure 6).
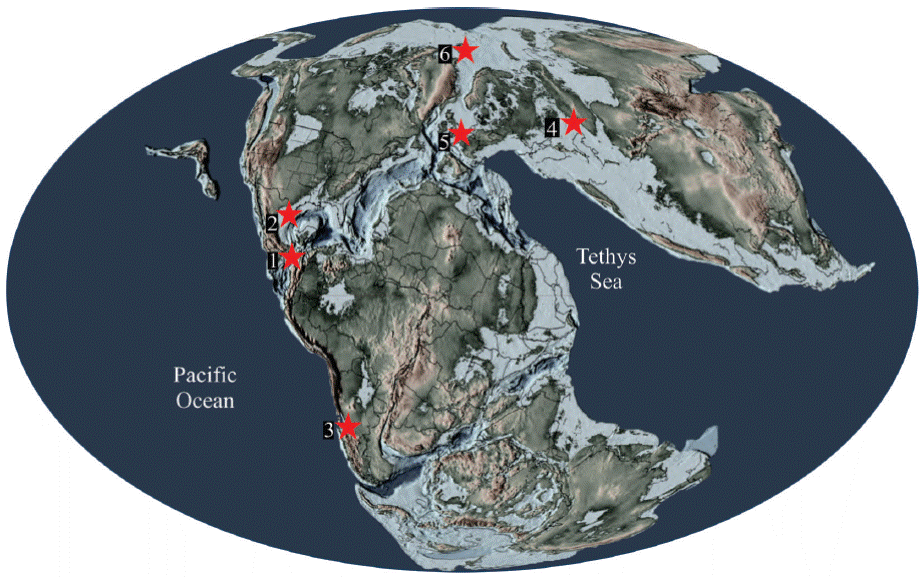
Figure 6 Distribution of Kimmeridgian-Thithonian pliosaurids 1, Liopleurodon ferox, and ndet. pliosaurids Sabinal formation Tlaxiaco Basin, Oaxaca, southern Mexico. 2, Indet. pliosaurids from La Caja and La Casita formations, Coahuila and Nuevo León, northern Mexico. 3, Pliosaurus patagonicus, Vaca Muerta Formation, Neuquen Basin, Argentina. 4, Pliosaurus rossicus, Volgian strata, Russia. 5) Pliosaurus brachyspondylus, Pliosaurus brachydeirus, Pliosaurus macromerus, Kimeridge Clay, England. 6), Pliosaurus funkei, Middle Volgian, Norway (redrawn from Scotese, 2014).
Although, IGM 4546 does not have enough features to support its taxonomical identity to genus and species level; IGM 4545 has a diagnostic character (the conical teeth with enamel ornamented with stout and continuous ridges) to include it as part of the genus Liopleurodon. This occurrence of IGM 4545 confirms the presence of Liopleurodon in the Kimmeridgian of the Hispanic Corridor area and represents the first occurrence of this taxon outside the European eastern Tethys Sea domians.

