











 nueva página del texto (beta)
nueva página del texto (beta) Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkINTRODUCCIÓN
La Región Mesopotámica de Argentina, delimitada por los ríos Paraná y Uruguay, abarca un área de 200,000 km2 aproximadamente e incluye a los territorios de las provincias de Corrientes, Entre Ríos y Misiones (Aceñolaza, 2007) (Figura 1). Los estudios realizados en los últimos años permitieron un notable incremento en el grado de conocimiento de las comunidades faunísticas de vertebrados que habitaron la región durante el Cuaternario (e.g., Zurita y Lutz, 2002; Carlini et al., 2004, 2008; Ferrero y Noriega, 2009; Zurita et al., 2014). En el caso particular de la provincia de Corrientes, los depósitos de la Formación Toropí/Yupoí son portadores de una fauna rica y diversa que incluye principalmente mamíferos y, en menor medida, reptiles (serpientes y tortugas) (Albino y Carlini, 2008; Zacarías et al., 2013; Zurita et al., 2014).
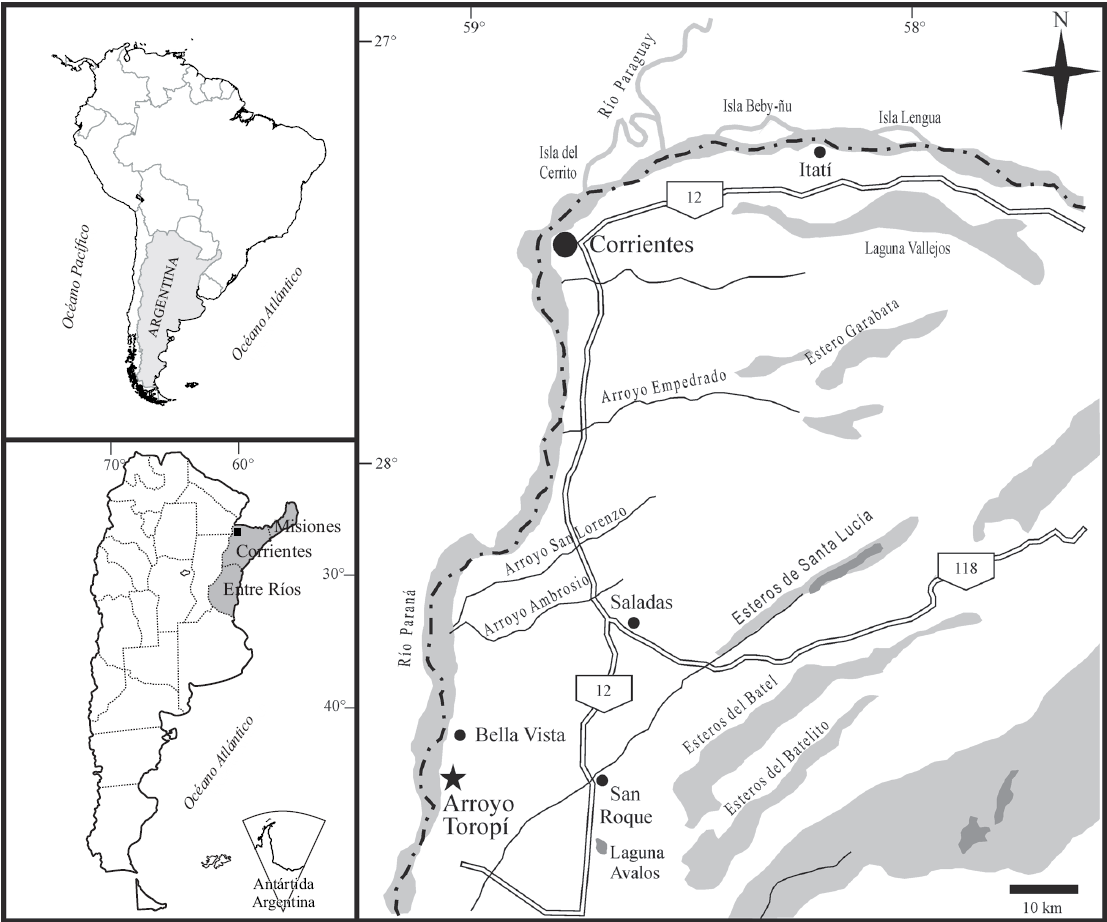
Figura 1 Ubicación geográfica de la localidad fosilífera Arroyo Toropí, provincia de Corrientes, Argentina.
Los mamíferos de la Formación Toropí/Yupoí están representados por Toxodontidae (Notoungulata), Mylodontidae y Megatheriidae (Tardigrada), Glyptodontidae, Dasypodidae y Pampatheriidae (Cingulata), Proterotheriidae (Litopterna), Caviidae y Myocastoridae (Rodentia), Cervidae, Camelidae y Tayassuidae (Artiodactyla), Equidae y Tapiridae (Perissodactyla), Felidae y Canidae (Carnivora) y Gomphotheriidae (Proboscidea) (Zurita et al., 2014, y referencias allí citadas). Diversos autores (e.g., Carlini et al., 2004; Miño-Boilini et al., 2006) destacaron que Toxodon Owen, 1837 (Toxodontidae) constituye el taxón con mayor cantidad de registros en esta unidad, representado por las especies T. platensisOwen, 1837 y T. gracilisGervais y Ameghino, 1880.
El hallazgo de estos vertebrados favoreció el desarrollo de numerosos trabajos sistemáticos, bioestratigráficos, paleoambientales y paleobiogeográficos, entre otros (Zurita y Lutz, 2002; Carlini et al., 2008; Francia, 2012; Zurita et al., 2014); sin embargo, hasta el momento, no existen estudios tafonómicos de detalle que involucren a sus restos. En este contexto, se destaca que si bien en los últimos años en Argentina, en particular en la región Pampeana, se incrementaron las contribuciones tafonómicas que incluyen vertebrados del Cenozoico (e.g., Montalvo, 2004; Montalvo et al., 2008; Montalvo et al., 2012; Tomassini, 2012; Tomassini y Montalvo, 2013; Tomassini et al., 2014a), esta temática ha sido escasamente considerada en el análisis de las asociaciones de megamamíferos cuaternarios (e.g., Fariña, 2002; Pomi, 2009; Tomassini et al., 2010, 2014b).
El objetivo del presente trabajo es describir las características tafonómicas, considerando particularmente aspectos mineralógicos, geoquímicos y paleohistológicos, presentes en costillas de T. platensis, procedentes de los depósitos de llanura de inundación de la Formación Toropí/Yupoí asignados al Pleistoceno Tardío, que afloran en el área de Arroyo Toropí (provincia de Corrientes, Argentina). Teniendo en cuenta los datos obtenidos se evalúan las modificaciones ocurridas durante la etapa fosildiagenética y los procesos responsables de las mismas, a fin de interpretar las posibles historias tafonómicas de los restos.
MARCO GEOGRÁFICO, ESTRATIGRÁFICO Y PALEOAMBIENTAL
El sitio fosilífero Arroyo Toropí (S 28°36', O 59°02') se encuentra ubicado al NE de la provincia de Corrientes (Argentina), aproximadamente 10 km al sur de la localidad de Bella Vista (Figura 1). En el área se reconocen importantes afloramientos de depósitos continentales cuaternarios, con espesores aproximados de 10 m. Herbst (1971) definió la Formación Yupoí para incluir dichos depósitos y propuso una subdivisión de la misma en dos secciones, inferior y superior, separadas por una costra silícea. Posteriormente, sobre la base de diferencias litológicas, mineralógicas y faunísticas, Herbst y Álvarez (1975) plantearon una modificación al esquema estratigráfico original de Herbst (1971) y denominaron Formación Toropí a los depósitos de la sección inferior y Formación Yupoí a los depósitos de la sección superior (ver Herbst y Santa Cruz, 1999). Por su parte, Iriondo (1993) consideró que las diferencias existentes no avalaban esta subdivisión y reunió los depósitos en una única unidad, denominada Formación Toropí/Yupoí.
La Formación Toropí/Yupoí (sensuIriondo, 1993) está dispuesta en discordancia erosiva sobre los depósitos de la Formación Ituzaingó asignados al Mioceno tardío-Plioceno (Brunetto et al., 2013) y muestra muy poca variación desde la base al techo, aunque pueden diferenciarse dos secciones (Figura 2). La sección inferior, de donde proceden los materiales aquí estudiados, está constituida por arenas finas a arenas limosas con colores que varían entre gris amarillento y verdoso. Los componentes minerales principales incluyen cuarzo y, en menor medida, feldespato, plagioclasa, opacos y vidrio volcánico. Presenta laminación paralela y son comunes los nódulos de manganeso. Una capa de hasta 50 cm con la misma litología que la descripta, pero más endurecida ("costra no calcárea" sensuHerbst, 1971; "costra silícea" sensu Tonni, 2009), la separa de la sección superior. Esta última comprende arenas limosas a arcillosas y limos arenosos, con colores variables entre gris verdoso y rosado a rojizo. Los componentes minerales principales tienen una representación semejante a la de la sección inferior. La estructura es maciza y presenta en forma dispersa escasos nódulos de yeso y de carbonato de calcio.
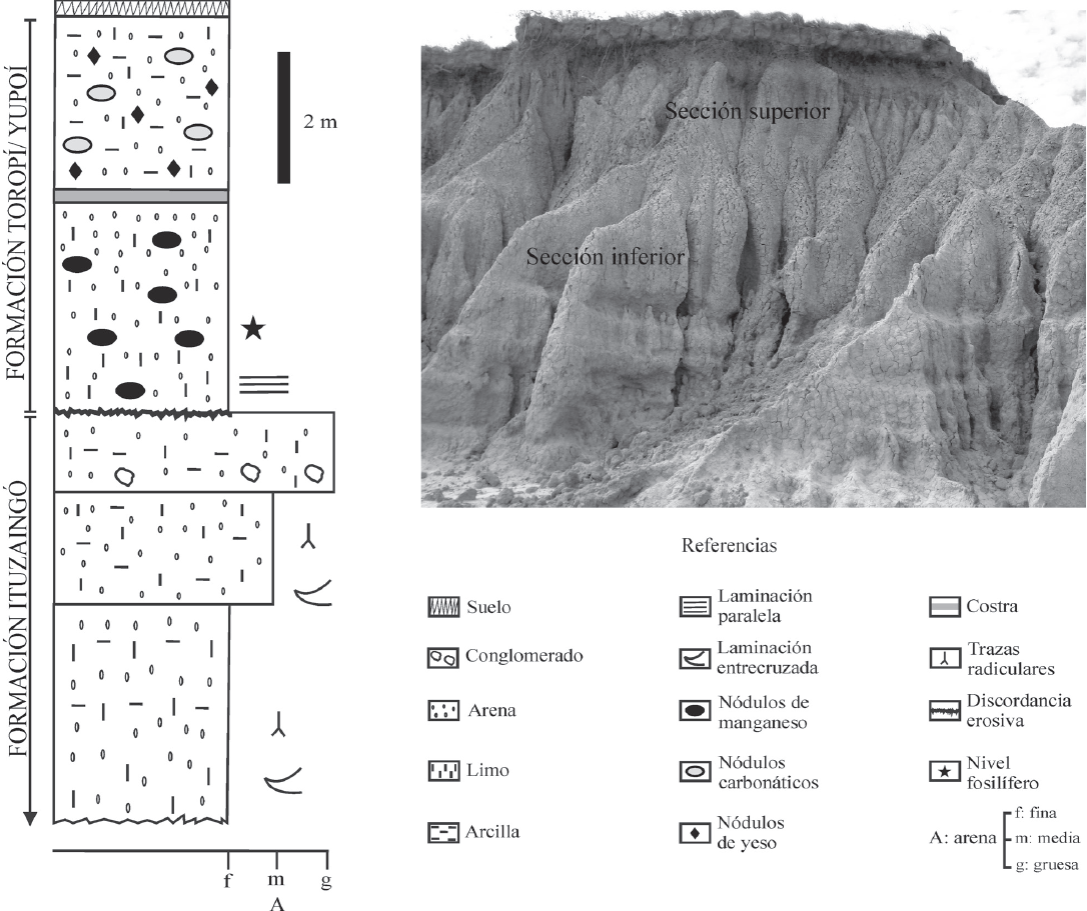
Figura 2 Sección estratigráfica en la localidad fosilífera Arroyo Toropí. Modificado de Zacarías et al. (2013). La foto del afloramiento muestra las dos secciones reconocidas en la Formación Toropí/Yupoí.
Iriondo (2010) y Erra et al. (2013) indicaron que los sedimentos de la Formación Toropí/Yupoí corresponden a depósitos de llanura de inundación acumulados a partir de la dinámica del río Paraná. Tonni et al. (2005) realizaron dataciones mediante OSL (Optically Stimulated Luminescence ) en la localidad de Arroyo Toropí, tanto en la parte inferior como en la superior de esta unidad, que arrojaron edades variables entre 52 y 36 ka AP. Otras dataciones más recientes, en la misma localidad y utilizando la misma metodología, proporcionaron edades entre 98 y 131 ka AP; sin embargo, se indicó que la mayor antigüedad obtenida en estas muestras podría estar relacionada con la naturaleza de los sedimentos y una posible contaminación con materiales más antiguos de la Formación Ituzaingó (Francia et al., 2012). Sobre la base de estos valores y de la asociación faunística de vertebrados presentes, los depósitos de esta formación fueron asignados al Pleistoceno Tardío (Francia, 2012; Zurita et al., 2014; y referencias allí citadas).
Desde el punto de vista faunístico, en la Formación Toropí/Yupoí se recuperaron taxones típicamente pampeanos (e.g., T. platensis, Notiomastodon platensis, Neosclerocalyptus paskoensis, Pampatherium typum) junto con otros de afinidades brasílicas (e.g., Boa constrictor, Holmesina paulacoautoi, Tapirus sp.). Teniendo en cuenta esto, se propuso que, al menos durante un intervalo del Pleistoceno Tardío, este sector de la Región Mesopotámica estuvo sujeto a variaciones climáticas, con alternancias de períodos fríos y áridos/semiáridos y períodos cálidos y húmedos (Carlini et al., 2004; Erra et al., 2013; Zurita et al., 2014).
MATERIALES Y MÉTODOS
Se analizó la mineralogía, geoquímica y paleohistología de ocho fragmentos de costillas pertenecientes a T. platensis (número mínimo de individuos = 2), procedentes de la localidad de Arroyo Toropí. Cuatro fragmentos corresponden al ejemplar CTES-PZ 1564 y cuatro al ejemplar CTES-PZ 1595. La asignación a nivel de especie de los especímenes estudiados se basó en su asociación con restos craneanos y mandibulares del taxón mencionado (ver Miño-Boilini et al., 2006). Los materiales se encuentran depositados en la Colección Paleontológica "Rafael Herbst" de la Facultad de Ciencias Exactas y Naturales y Agrimensura, Universidad Nacional del Nordeste (provincia de Corrientes, Argentina) bajo el acrónimo CTES-PZ.
Según Miño-Boilini et al. (2006: 96) los materiales estudiados en este trabajo provienen de las "formaciones Toropí y Yupoí", sin mayores detalles; sin embargo, observaciones personales de uno de los autores (ARM-B) permiten asignarlos con mayor precisión a la parte inferior de la Formación Toropí/Yupoí (sensuIriondo, 1993). Se optó por evaluar los restos de T. platensis ya que constituye el taxón mejor representado en esta formación.
En forma complementaria a las cuestiones que aquí se analizan, también se evaluaron otros atributos tafonómicos presentes en los restos, incluyendo articulación, meteorización, abrasión, rotura, tipos de fracturas y encostramiento. Para la determinación del color de la superficie externa de los fósiles se utilizó la Rock Color Chart (Goddard et al., 1948).
Las características de preservación de la microestructura ósea de las costillas se evaluaron a partir del análisis de secciones delgadas transversales, las cuales se confeccionaron en el Laboratorio de Petrotomía del INGEOSUR-Departamento de Geología de la Universidad Nacional del Sur (Bahía Blanca, Buenos Aires), siguiendo la metodología indicada por Tomassini (2012), y fueron observadas con un microscopio Nikon ECLIPSE 50i POL y un microscopio Leica DMLP. La composición mineralógica se determinó utilizando un difractómetro Rigaku D-Max III-C perteneciente al INGEOSUR-Departamento de Geología de la Universidad Nacional del Sur. La composición química elemental de los fósiles y los sedimentos hospedantes se determinó mediante un sistema EDX (Energy Dispersive X-ray) marca EDAX, modelo DX-4, con capacidad de detección desde Boro (B) hasta Uranio (U). Algunas secciones delgadas se fotografiaron con un microscopio electrónico de barrido JEOL 35 CF SEM. Ambos equipos pertenecen a la Unidad de Administración Territorial del Centro Científico y Tecnológico CONICET Bahía Blanca (CCT, Bahía Blanca, Buenos Aires).
RESULTADOS
Los fragmentos de costillas de los dos ejemplares de T. platensis presentan claras diferencias con relación al color de la superficie externa. Los especímenes incluidos en CTES-PZ 1564 son pardo claro amarillentos (10YR 6/2) (Figura 3a), mientras que los de CTES-PZ 1595 son completamente negros (N1) (Figura 3b).

Figura 3 Variaciones de color en la superficie externa de las costillas, a) ejemplar pardo claro amarillento (CTES-PZ 1564). Las flechas señalan la presencia de costras carbonáticas, b) ejemplar negro (CTES-PZ 1595). Escala 4 cm.
En cada caso, los materiales estudiados fueron recuperados in situ junto con otros restos craneanos y postcraneanos, y pueden considerarse como desarticulados pero asociados (sensuBehrensmeyer, 1991). En todas las costillas se observaron signos de meteorización, evidenciados por un leve agrietamiento de la superficie externa (estadío 1 sensuBehrensmeyer, 1978). No se reconocieron indicios de abrasión (categoría 0 sensuAlcalá, 1994). Todas están incompletas y presentan fracturas longitudinales y escalonadas con superficies irregulares y fracturas transversales con superficies lisas (sensuMarshall, 1989). Los ejemplares pardo claro amarillentos (CTES-PZ 1564) presentan además la superficie externa parcialmente cubierta por costras carbonáticas de diferentes dimensiones (Figura 3a).
En los diagramas de rayos X se identificaron dos fases minerales principales, hidroxiapatita [Ca5(PO4)3(OH)] en los ejemplares de color negro (Figura 4a) y carbonatohidroxiapatita o dahllita [Ca5(PO4CO3)3(OH)] en los ejemplares de color pardo claro amarillento (Figura 4b). Los diagramas de EDX mostraron la presencia de P, Ca, C, O, Na, Mg, Al, Si y K en todas las muestras (los últimos siete elementos no representados en los diagramas) (Figura 5a-5b); además, en los ejemplares de color negro también se determinó Fe (Figura 5b). A excepción del P, los diagramas de EDX de los sedimentos hospedantes revelaron la presencia de los mismos elementos que los registrados en los restos fósiles (Figura 5c).
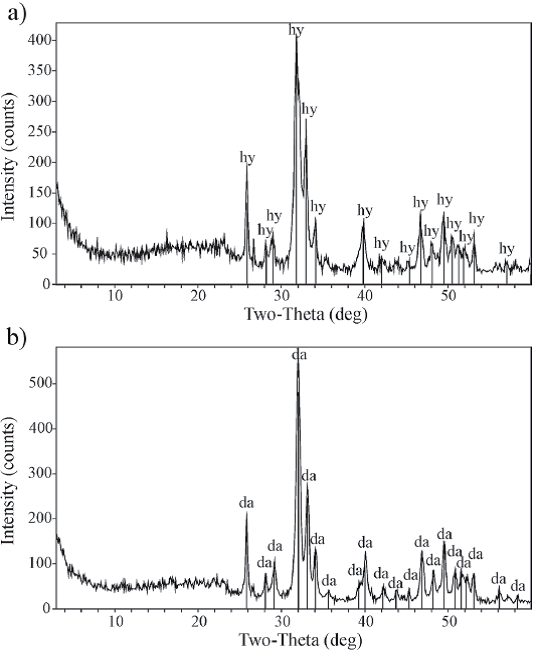
Figura 4 Difractogramas de rayos-X de las costillas, a) ejemplar negro (CTES-PZ 1595), la fase mineral principal es hidroxiapatita, b) ejemplar pardo claro amarillento (CTES-PZ 1564), la fase mineral principal es dahllita.
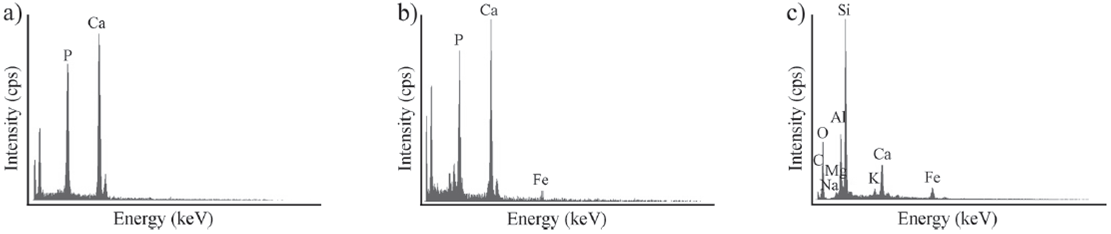
Figura 5 Diagramas de composición química elemental de las costillas, a) ejemplar pardo claro amarillento (CTES-PZ 1564), b) ejemplar negro (CTES-PZ 1595), c) sedimento hospedante.
En las secciones delgadas no se identificaron diferencias microestructurales que se pudieran vincular con el color de los especímenes. En todos los cortes se identificaron claramente dos tipos de tejido óseo. Por un lado, hueso compacto con desarrollo de osteonas secundarias, canales vasculares de Havers y Volkmann, lagunas de osteocitos y canalículos y, por otro, hueso esponjoso con desarrollo de trabéculas y cavidades intertrabeculares (Cuezva y Élez, 2000; Chinsamy-Turan, 2005) (Figura 6a).
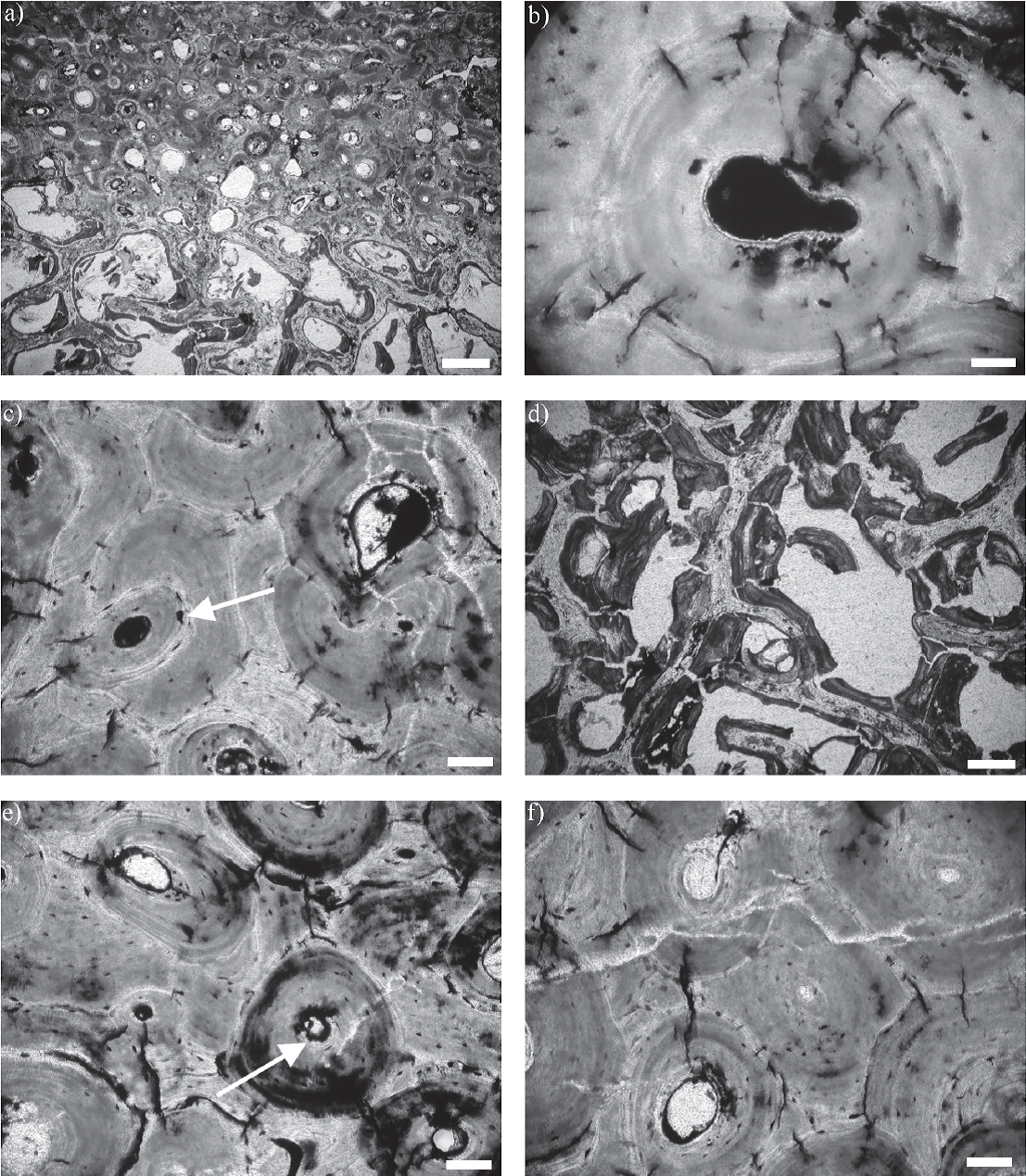
Figura 6 Secciones delgadas transversales de costillas (CTES-PZ 1595) analizadas con microscopio petrográfico, a) detalle de las zonas de hueso compacto y hueso esponjoso. Escala 400 µm, b) osteona secundaria con microgrietas radiales. Escala 10 µm, c) elementos microestructurales y microgrietas radiales rellenas con óxidos de hierro. La flecha señala una laguna de osteocito modificada debido al reemplazo con óxidos de hierro. Escala 50 µm, d) trabéculas fragmentadas y colapsadas correspondientes a la zona de hueso esponjoso; en algunas de ellas se observa la matriz ósea reemplazada con óxidos de hierro. Todos los espacios intertrabeculares están vacíos. Escala 200 µm, e) reemplazo de la matriz ósea con óxidos de hierro en las laminillas concéntricas y en las líneas de cemento. La flecha señala un canal de Havers con desarrollo de márgenes irregulares debido al reemplazo con óxidos de hierro. Escala 50 µm, f) fisuras vacías, con arreglo poligonal, atravesando la microestructura ósea. Escala 50 µm.
En todos los cortes realizados se observó la presencia de pequeñas microgrietas asociadas a las osteonas secundarias (Figura 6b y 7a). Estas microgrietas atraviesan las líneas de cemento y conectan osteonas adyacentes. Están orientadas radialmente y se extienden, por un corto trayecto, desde el margen externo hacia el interior de las osteonas, sin alcanzar el centro de las mismas. La longitud varía entre 27 µm y 105 µm aproximadamente. La cantidad de microgrietas por osteona varía entre 4 y 11, con un promedio de 7. Se obtuvo un diagrama de rosas para estas estructuras que refleja un patrón de orientaciones de tipo polimodal (Figura 8).
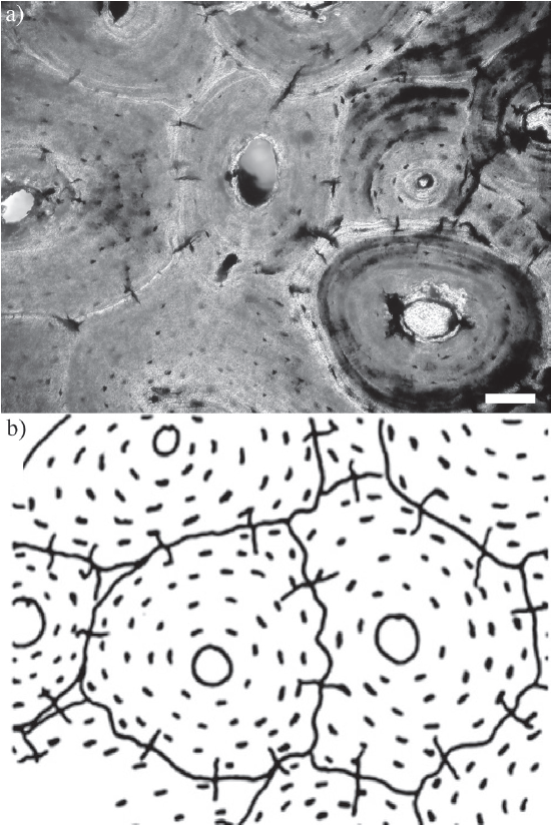
Figura 7 Microgrietas radiales en osteonas secundarias, a) sección delgada transversal de costilla (CTES-PZ 1595). Escala 50 µm, b) esquema propuesto por Pfretzschner y Tütken (2011). Sin escala.
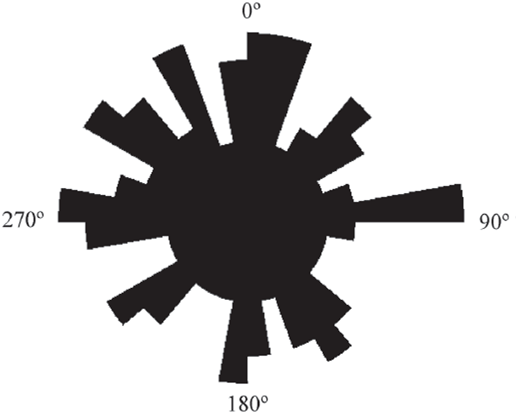
Figura 8 Diagrama de rosas mostrando las orientaciones de las microgrietas radiales. Se tomaron 171 mediciones en 28 osteonas secundarias de una costilla (CTES-PZ 1595).
Las cavidades correspondientes a canales vasculares, lagunas de osteocitos y canalículos, así como también las microgrietas radiales, están en general rellenas con óxidos de hierro. Si bien en algunos casos el relleno sólo afecta los márgenes de estas cavidades, en la mayoría de los casos las rellena totalmente (Figura 6c). Las Figuras 9a y 9b muestran canales vasculares parcialmente rellenos, en los que se distinguen cristales tabulares de óxidos de hierro. Los espacios intertrabeculares correspondientes a la zona de hueso esponjoso, por otro lado, están vacíos (Figura 6d).
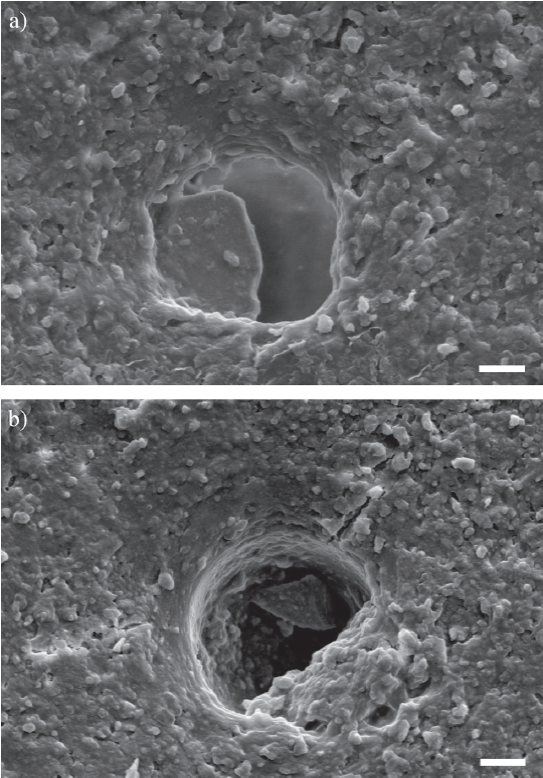
Figura 9 Secciones delgadas transversales de costillas analizadas con micros-copio electrónico de barrido. Se observa la presencia de canales vasculares parcialmente rellenos, a) ejemplar negro (CTES-PZ 1595), b) ejemplar pardo claro amarillento (CTES-PZ 1564). Escala 3 µm.
Se observó que la mayoría de las lagunas de osteocitos que están totalmente rellenas tienen un tamaño mayor que las vacías e inclusive con modificación de la forma original típica de estas estructuras (Figura 6c). Por otro lado, algunos de los canales de Havers con indicios de relleno presentan márgenes irregulares (Figura 6e). Cabe destacar que la presencia de óxidos de hierro también se determinó en las líneas de cemento y las laminillas concéntricas de las osteonas secundarias (Figura 6e), en este último caso en forma de parches irregulares, y en las trabéculas correspondientes a la zona de hueso esponjoso (Figura 6d).
En dos cortes, uno correspondiente a CTES-PZ 1564 (negro) y otro a CTES-PZ 1595 (pardo claro amarillento), se identificaron numerosas fisuras de mayor tamaño que las microgrietas radiales, dispuestas siguiendo un arreglo poligonal, que atraviesan parcial o totalmente la zona de hueso compacto (Figura 6f). Todas estas fisuras están vacías y, en algunos casos, cortan elementos microestructurales rellenos. Además, en ambos cortes se observó que muchas de las trabéculas correspondientes a la zona de hueso esponjoso se encuentran rotas y colapsadas, lo que da lugar al desarrollo de estructuras brechosas formadas por fragmentos óseos de diferentes dimensiones (Figura 6d).
DISCUSIÓN
Toxodon platensis es un notoungulado nativo de Sudamérica, de grandes dimensiones y hábitos pastadores, vinculado a áreas abiertas con presencia de cuerpos de agua estables (Bond et al., 1995; Bond, 1999); en consecuencia la preservación de sus restos suele estar asociada a depósitos fluviales. El biocrón de este taxón abarca desde el Pleistoceno Medio (Piso/Edad Bonaerense) hasta el Pleistoceno Tardío (Piso/Edad Lujanense) y los registros son muy numerosos tanto en Argentina como en Uruguay, Brasil, Paraguay, Bolivia y Venezuela (Bond et al., 1995; Miño-Boilini et al., 2006). Como se mencionó, en los depósitos de la Formación Toropí/Yupoí asignados al Pleistoceno Tardío sus restos son particularmente abundantes (Carlini et al., 2004; Miño-Boilini et al., 2006).
En el caso de las muestras analizadas, la presencia de especímenes desarticulados pero asociados y con indicios de meteorización leve, indicaría que el tiempo de permanencia de estos elementos esqueléticos en la superficie fue relativamente corto (Behrensmeyer, 1978, 1991). No se reconocieron evidencias concretas que sugieran que hayan sido afectados por transporte. Tanto las fracturas longitudinales como las escalonadas pueden vincularse a procesos previos al enterramiento, cuando el hueso aún se encontraba fresco (Behrensmeyer et al., 1989); mientras que las fracturas transversales se asocian a procesos ocurridos luego del enterramiento, una vez que los huesos perdieron sus componentes orgánicos y se mineralizaron (Alcalá y Martín Escorza, 1998). El encostramiento es un proceso posterior al enterramiento que estaría vinculado a la precipitación de soluciones carbonáticas a partir de los fluidos que circulaban por los niveles portadores (Fernández-López, 2000). Todas estas características son concordantes con un ambiente de llanura de inundación.
El reconocimiento de dos fases minerales en los diagramas de rayos X refleja diferencias composicionales entre los especímenes de los dos individuos identificados. La presencia de hidroxiapatita en los especímenes de color negro indicaría que la estructura interna original se preservó sin cambios importantes, mientras que la presencia de dahllita en los especímenes de color pardo claro amarillento representaría un enriquecimiento en carbonato a partir de la sustitución iónica de PO3- 4 por CO2- 3. Este reemplazo habría tenido lugar durante la fosildiagénesis temprana (Elorza et al., 1999; Luque et al., 2009; Merino y Buscalioni, 2013) y estaría vinculado al desarrollo del encostramiento superficial observado en estos especímenes.
La determinación de P y Ca en los diagramas de EDX avala la composición fosfato cálcica de los materiales. A su vez, las similitudes reconocidas en la composición química entre los fósiles y los sedimentos hospedantes permiten inferir que la incorporación de nuevos elementos en los restos se produjo, a lo largo de la fosildiagénesis, por un intercambio con el ambiente donde los mismos fueron enterrados y preservados. Este fenómeno se relaciona directamente con la composición de los fluidos que circulaban por los niveles portadores y que movilizaban los elementos disueltos (e.g., Henderson et al., 1983; Martin et al., 2005; Luque et al., 2009; Merino y Buscalioni, 2013). Tal como fue propuesto por diversos autores para otros yacimientos (e.g., Bao et al., 1998; Pfretzschner, 2001; Farlow y Argast, 2006), en esta oportunidad se interpreta que el enriquecimiento con Fe y su posterior precipitación en forma de óxidos, generó las impregnaciones que dieron lugar a la coloración oscura de la superficie externa en el caso de los especímenes de CTES-PZ 1595.
El análisis de las secciones delgadas permitió reconocer el desarrollo y relleno de microgrietas radiales, permineralización de las cavidades correspondientes a elementos microestructurales, reemplazo de la matriz ósea y desarrollo de fisuras. Sin embargo, a pesar de esto, la microestructura ósea evidenció un grado de preservación muy bueno en todos los ejemplares, correspondiente a las categorías 4 y 5 (porcentaje de integridad >85%) de la clasificación histológica propuesta por Hedges et al. (1995).
Las microgrietas radiales identificadas presentan características similares a las descriptas por Pfretzschner (2000, 2004, 2006) (Figura 7a-7b). Según este autor, las mismas se producen durante la etapa final de la fosildiagénesis temprana, en osteonas secundarias de elementos esqueléticos preservados bajo condiciones acuáticas. Una vez que ocurre la gelatinización del colágeno, debido a su ruptura en cadenas de proteínas más cortas, la absorción de agua y la consecuente hinchazón por hidratación generan condiciones de estrés tensional en las paredes mineralizadas de las osteonas hasta que finalmente se alcanza un punto crítico y ocurre el agrietamiento que atraviesa las líneas de cemento. Estas microgrietas tienen un rol muy importante en la fosilización de los elementos esqueléticos, ya que constituyen caminos adicionales (junto con los canales vasculares y los canalículos) que favorecen la movilización de fluidos por difusión en la zona de hueso compacto y, por lo tanto, mejoran el intercambio de iones (Pfretzschner, 2000, 2004; Pfretzschner y Tütken, 2011).
La presencia de microgrietas radiales resulta coherente con la preservación de las costillas en un ambiente de llanura expuesto a eventos de inundación. Por otra parte, la obtención de un diagrama de rosas de tipo polimodal refleja que la distribución del esfuerzo que le dio origen a estas estructuras fue homogéneo y sin orientaciones preferenciales, probablemente debido a que los depósitos se encontraban periódicamente saturados de agua.
Los fluidos ricos en Fe favorecieron la permineralización de los distintos elementos microestructurales (e.g., canales vasculares, lagunas de osteocitos, canalículos) y el relleno de las microgrietas radiales. La precipitación de los óxidos de hierro habría tenido lugar en una etapa próxima a la finalización de la fosildiagénesis temprana, con posterioridad al desarrollo de las microgrietas radiales pero antes que la recristalización del apatita cerrara los espacios vacíos y restringiera la circulación de fluidos (ver Pfretzschner, 2001).
Durante esta etapa, se puede interpretar que los fluidos ricos en Fe también aprovecharon el desarrollo de nuevos caminos de difusión (e.g., microgrietas radiales) para distribuirse por distintos sectores de la microestructura. La precipitación de los óxidos de hierro generó el reemplazo de la matriz ósea observado tanto en las laminillas concéntricas y en las líneas de cemento de las osteonas secundarias como en las trabéculas correspondientes a la zona de hueso esponjoso. La amplia distribución de estos óxidos, en particular en el sector de las líneas de cemento, estructuras fuertemente mineralizadas in vivo y que además se mantienen muy estables en la diagénesis (Pfretzschner, 2000, 2001), permitiría estimar que este proceso fue muy intenso. Al respecto, de acuerdo a lo expuesto por Pfretzschner (2001) y Pfretzschner y Tütken (2011), el mayor tamaño de las lagunas de osteocitos que están completamente rellenas y el desarrollo de márgenes irregulares de algunos canales de Havers rellenos constituirían evidencias de reemplazo en las zonas adyacentes al elemento microestructural.
La formación de los óxidos se produjo a partir de la precipitación del Fe que estaba disuelto en los fluidos circulantes. El comportamiento de las diferentes formas de hierro depende directamente de las condiciones de reducción/oxidación y acidez/alcalinidad. Si bien no se conocen con certeza los parámetros exactos, la precipitación de los óxidos de hierro está asociada a entornos alcalinos y oxidantes, es decir con valores altos de pH y de Eh (Luque et al., 2009; Pfretzschner y Tütken, 2011). El comportamiento de las distintas formas de Fe probablemente estuvo vinculado a la circulación de fluidos en los depósitos de llanura de inundación; durante los períodos de saturación en agua habría permanecido en su fase soluble y móvil, mientras que durante los períodos de sequía y aridez precipitó en forma de óxidos.
En la etapa inicial de la fosildiagénesis temprana los valores de pH son elevados (entre 8 y 10), debido a la composición fosfato-cálcica de los restos. Por otro lado, los valores de Eh son muy bajos (-200 mV), fundamentalmente por el consumo de oxígeno implicado en la degradación del colágeno y la limitada conexión que existe con el exterior ya que la movilización por difusión en las osteonas está restringida. Durante la etapa final de la fosildiagénesis temprana, momento para el que se interpretó la formación de los óxidos de hierro presentes en los fósiles analizados, los valores de pH se mantienen altos por la cantidad de fosfato presente en el apatito generado, mientras que los de Eh se incrementan hacia los valores del ambiente de preservación debido a la apertura de nuevos caminos (e.g., microgrietas radiales) que favorecen la conexión con el entorno exterior (Pfretzschner, 2000, 2004; Pfretzschner y Tütken, 2011).
Las características de las fisuras de mayor tamaño (e.g., arreglo poligonal, disposición independiente de la estructura histológica), sumado a que todas ellas están vacías y que, en algunos casos, cortan elementos microestructurales permineralizados, permiten inferir que su formación tuvo lugar durante alguna etapa de la fosildiagénesis tardía. Su desarrollo se puede vincular con la presión generada por la carga de los sedimentos suprayacentes. Se estima que esta presión también sería responsable de la intensa fragmentación y colapso observado en las trabéculas correspondientes a la zona de hueso esponjoso, probablemente favorecida por la ausencia de relleno en las cavidades intertrabeculares.
CONCLUSIONES
Las asociaciones de vertebrados de la Región Mesopotámica de Argentina no habían sido, hasta el momento, objeto de análisis tafonómicos de detalle. En este trabajo se realiza una primera aproximación a este tipo de estudios a partir de la evaluación de fragmentos de costillas de T. platensis del Pleistoceno Tardío de la provincia de Corrientes, que permitió interpretar los diversos procesos fosildiagenéticos que los afectaron.
Los fósiles analizados evidenciaron modificaciones en sus características mineralógicas, geoquímicas e histológicas originales, debido al desarrollo de diversos procesos post-enterramiento relacionados con el ambiente de llanura de inundación en el que se preservaron. Los cambios en la composición mineralógica y geoquímica, impregnación de la superficie externa, formación y relleno de microgrietas radiales, permineralización de elementos microestructurales y reemplazo de la matriz ósea, ocurrieron durante la fosildiagénesis temprana, mientras que el desarrollo de fisuras y la fragmentación de las trabéculas tuvieron lugar durante la fosildiagénesis tardía. La precipitación de óxidos de hierro generó la coloración oscura que muestran algunos de los especímenes. Las similitudes encontradas entre los especímenes interpretados como provenientes de dos individuos (CTES-PZ 1564 y CTES-PZ 1595) reflejan el desarrollo de historias tafonómicas semejantes.
Teniendo en cuenta que los restos estudiados corresponden al taxón más abundante registrado en la Formación Toropí/Yupoí, se estima que los resultados obtenidos son representativos de toda la asociación. Por lo tanto, las interpretaciones realizadas pueden extrapolarse a otros especímenes procedentes de los mismos niveles portadores, particularmente aquellos pertenecientes a megamamíferos.

