










Servicios Personalizados
Revista
Articulo
 Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista mexicana de ciencias geológicas
versión On-line ISSN 2007-2902versión impresa ISSN 1026-8774
Rev. mex. cienc. geol vol.32 no.1 Ciudad de México abr. 2015
Ostracode and C/N based paleoecological record from Santiaguillo basin of subtropical Mexico over last 27 cal kyr BP
Registro paleoecológico basado en ostracodos y C/N de la cuenca de Santiaguillo en la parte subtropical de México durante los últimos ca.27 ka cal AP
Claudia M. Chávez-Lara1, Priyadarsi D. Roy2*, Liseth Pérez2, Gowrappan Muthu Sankar3, and Víctor H. Lemus Neri4
1 Posgrado en Ciencias de la Tierra, Universidad Nacional Autónoma de México, Ciudad Universitaria, C.P. 04510, México DF, México.
2 Instituto de Geología, Universidad Nacional Autónoma de México, Ciudad Universitaria, C.P. 04510, México D.F., México. roy@geologia.unam.mx, priyadarsi1977@gmail.com
3 French Institute of Pondicherry, 11 St Louis Street, P.B. 33, Puducherry 605 001, India.
4 USAI, Facultad de Química, Universidad Nacional Autónoma de México, Ciudad Universitaria, C.P. 04510, México D.F., México.
Manuscript received: April 30, 2014
Corrected manuscript received: Nobember 17, 2014
Manuscript accepted: November 19, 2014
ABSTRACT
Changes in ostracode species assemblage suggest presence of oligohaline and mesohaline waters in the Santiaguillo basin of subtropical Mexico over the last ca.27 cal kyr BP. Paleoecological reconstruction is based on the distribution of Limnocythere bradburyi Forester, Eucandona cf. patzcuaro Tressler, Cypridopsis vidua Brady and Limnocythere ceriotuberosa Delorme. Source of organic productivity is inferred from C/N ratios. The basin hosted shallow and oligohaline water during the late glacial (ca.27–19 cal kyr BP) and lacustrine phytoplankton contributed to the organic productivity. However, the water body of Last Glacial Maximum (LGM) was relatively warmer (>13°C) and more diluted compared to the pre-LGM. Increased salinity and reduced lacustrine productivity at ca.19 cal kyr BP marked a shift in the paleoecological conditions. Water body was mesohaline during ca.9.5–4 cal kyr BP and periodic desiccation of the basin possibly caused the absence of ostracode valves in sediments of the last ca.3.5 cal kyr BP. Comparison with other proxy-records from the region indicates that the hydrological variations in different parts of subtropical north and northwestern Mexico were not synchronous during the late Pleistocene and Holocene.
Keywords: lacustrine ostracodes; C/N ratio; Paleoecology; Late Quaternary; Santiaguillo basin; Mexico.
RESUMEN
Los cambios en el conjunto de especies de ostrácodos sugieren presencia de oligohalina y mesohalina aguas en la cuenca de Santiaguillo ubicado en la parte subtropical de México durante los últimos ca.27 ka cal AP. La reconstrucción paleoecológica se realizó con base en la distribución de Limnocythere bradburyi Forester, Eucandona patzcuaro Tressler, Cypridopsis vidua Brady y Limnocythere ceriotuberosa Delorme. La fuente de la productividad orgánica se infiere a partir de la relación C/N. La cuenca albergó agua somera y oligohalina durante el pasado último glacial (ca. 27–19 ka cal AP) y el fitoplancton lacustre contribuyó a la productividad orgánica. Sin embargo, el cuerpo de agua durante el Último Máximo Glacial (UMG) fue relativamente cálido y más diluido comparado con el pre-UMG. Un incremento en la salinidad y la reducción de productividad lacustre a los ca.19 ka cal AP marcaron un cambio en las condiciones paleoecológicas. El cuerpo de agua fue mesohalino durante ca.9.5–4 ka cal AP y sequías periódicas de la cuenca causaron posiblemente la ausencia de valvas de ostrácodos en los sedimentos de los últimos ca.3.5 ka cal AP. Comparaciones con otros registros de la región indican que las variaciones hidrológicas en diferentes partes de la zona subtropical del norte y noroeste de México no fueron sincrónicas durante el Pleistoceno tardío y Holoceno.
Palabras clave: ostrácodos lacustre; relación de C/N; Paleoecología; Cuaternario tardío; cuenca de Santiaguillo; México.
INTRODUCTION
In the last two decades, the reconstruction of late Quaternary paleoecological and paleoclimatic conditions of the subtropical northern Mexico has received a significant increase in the scientific attention (Urrutia-Fucugauchi et al., 1997; Metcalfe et al., 1997, 2002; Murillo de Nava et al., 1999; Ortega-Guerrero et al., 1999; Palacios-Fest et al., 2002; Lozano-García et al., 2002; Holmgren et al., 2003, 2007; Davis, 2003; Caballero et al., 2005; Castiglia and Fawcett, 2006; Ortega-Rosas et al., 2008; Roy et al., 2010, 2012a, 2012b, 2013a, 2013b, 2014a, 2014b). The initial proxy registers and climate models reported humid conditions during the late last glacial and associated them to more winter precipitation. The subtropical high pressure zone in the eastern Pacific displaced southward and increased the frequency of westerly winter storms (COHMAP members, 1988; Manabe and Broccoli, 1985; Kutzbach and Wright, 1985; Kutzbach et al., 1998). Most of the paleoclimatic records suggested the possible absence of summer precipitation in the region (e.g. Thompson and Anderson, 2000).
The perception about the late Quaternary climate changed after Holmgren et al. (2006) and Kirby et al. (2006) reported that the influence of summer precipitation was reaching the USA-Mexico borderlands and coastal southwestern USA during different intervals in the last glacial period. Recently, Roy et al. (2013a) reconstructed paleohydrological conditions of the northern part of subtropical Mexico over the last ca. 80 cal kyr BP and hypothesized that the major influence of the winter storms remained in the region located to the north of 29°N latitude. The hydrology of northern Mexico was dominantly controlled by the summer precipitation and the intervals of increased summer precipitation were contemporary to warm interstadials (Roy et al., 2013a). Summer precipitation was less during the Last Glacial Maximum (LGM) and it increased over the deglaciation (Roy et al., 2010; Roy et al., 2012b). During the Pleistocene-Holocene transition, increased summer precipitation led to wetter conditions in the region (Roy et al., 2014a). However, the synthesis of Barron et al. (2012) suggested that the influence of summer precipitation reached different regions during different intervals over the Holocene.
Compared to the northern part of subtropical Mexico, the paleoecological and paleohydrological information are scarce from the central and southern parts (Figure 1a). Similarly, most of the proxy-records are based on the geochemical and magnetic properties of lacustrine sediments and the use of ostracodes as a paleoenvironment indicator has not been sufficiently explored. Ostracodes are micro-crustaceans with low-Mg calcite valves (Turpen and Angell, 1971) and are sensible to the environmental and climatic changes. Their abundance and diversity in lacustrine sediments provide important information about the past ecology and physicochemical conditions of the water column (e.g. water chemistry, salinity, conductivity and temperature) (Forester, 1986; Pérez et al., 2013). The results obtained from previous studies of ostracode paleoecology from the region (i.e. Babicora and Bahia Kino) are promising (Caballero et al., 2005; Palacios-Fest et al., 2002; Chávez-Lara et al., 2012). In this work, we infer the paleoecology of the central-southern part of the subtropical Mexico over the last ca. 27 cal kyr BP by studying the distributions and abundances of fossil ostracode valves in two different sediment profiles from the western margin of Santiaguillo basin (Figure 1b). Productivity is inferred from the concentrations of total organic carbon and the source of productivity is distinguished by the C/N ratio.
REGIONAL SETTING
Subtropical Mexico hosts large parts of Sonora and Chihuahua Deserts. The Santiaguillo basin (24º39'–24º58' N, 104º39'–104º57' W and ~1960 m asl) is located in a NW-SE oriented tectonic basin in the central part of the Chihuahua Desert of Mexico and forms part of the "Basin and Range" sub-province of Sierra Madre Occidental Mountains (Figure 1b). The geology and tectonic evolution of the basin is described by Nieto-Samaniego et al. (2012). The semi-graben was formed between ca.39 and 32 Ma and igneous rocks (rhyolite, ignimbrite and basalt) comprise the geology of the basin. Minor exposures of lacustrine and alluvium deposits and conglomerates are also present in the basin surroundings. The lacustrine basin covers an area of ~2000 km2 and is surrounded in the east and west by ~2500 to 3000 m high hills.
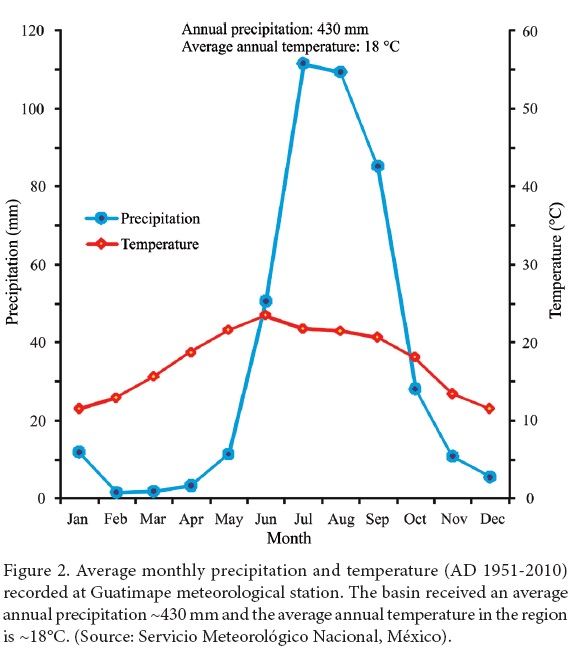
The meteorological station at Guatimape (western part of the basin) registered an average annual precipitation of ~430 mm between AD 1951 an 2010 (Servicio Meteorológico Nacional, México). More than 80% of this annual precipitation occurred during the summer months of June to September as a result of monsoonal circulation and tropical cyclones, whereas the precipitation during winter months of November to March represented less than 10% of the annual rainfall (Figure 2). The registered maximum average temperature was ~34°C during June, and minimum temperature reached sub-zero during December-January. According to the classification of Köppen, the region represents semi-arid climate with precipitation during the summer (BSk) (García, 1973).
MATERIAL AND METHODS
Sediment samples were collected from two different pits (S1 and S2) at the western margin of the basin (Figure 1b). S1 and S2 pits have depths of 300 cm and 200 cm, respectively (Figure 3). Chronology of the sediment sequence from S1 pit is based on 4 different Accelerator Mass Spectrometry radiocarbon(AMS 14C) dates on organic carbon present in the bulk sediments. Similarly, the sediment profile of S2 pit is chronologically constrained by 2 different AMS 14C dates on sediment organic carbon (Figure 3). The 14C dates were calibrated in Calib 6.0 program using the IntCal09 calibration curve of Reimer et al. (2009) and the values of maximum probability within the 2σ range were considered for the reconstruction of age models.
A total of 61 samples at an interval of ~5 cm from the S1 pit and 20 different samples at an interval of ~10 cm from S2 pit were processed for the ostracode analysis. Each sample (~10 g) was oven dried and disintegrated by soaking in water for 2–3 days. The samples were washed through two sieves of 74 µm and 177 µm. The sieve residues were oven dried and examined under a Leica stereomicroscope to identify the different species and determine their abundance based on the descriptions provided by Morkhoven (1963), Delorme (1971a, 1971b, 1971c), Forester (1985) and Carreño (1990). Total ostracode abundance in each sample was calculated by adding the number of adult and juvenile valves of all the observed species and is reported as the number of valves in 1 g of dry sediment (valves/g). Adult valves of each species were counted in the dry sediment and presented as relative abundance (in %). The adult valves were extracted and stored in Plummer type microfossil slides. Images of the ostracode valves were taken using a Jeol JSM-5600LV LCM scanning electron microscope.
Concentration of total organic carbon (TOC) in the bulk sediments was analyzed in all the samples in a Thermo Scientific HiperTOC solid analyzer after oven drying (50 °C) and powdering the sediments in an agate pestle. Concentration of total carbon (TC) was measured by heating the sample to 980°C and the amount of inorganic carbon (TIC) was measured by acidifying the sediment with 10% H3PO4. Total organic carbon (TOC) content was estimated by subtracting the inorganic carbon (TIC) from total carbon (TC). Concentration of total nitrogen (TN) was measured in 51 samples (31 samples from S1 pit and 20 samples from S2 pit) in a CHNS/O 2400 series II Perkin Elmer elemental analyzer.
RESULTS
Sediment and Chronology
Figure 3 shows the sediment profiles of S1 and S2 pits. Sediments of S1 pit comprise clay and calcareous silt intercalations from 300 to 278 cm depth and massive calcareous silt and silty-sand from 278 to 75 cm depth. Silty-clay and silt intercalations are deposited between 75 and 50 cm. Massive silty-sand with abundant root remnants are present between 50 and 0 cm. Almost 65 cm long vertical desiccation fissures are present at 75 cm depth. In the S2 pit, the sediments are calcareous silt and silty-sand between 200 and 100 cm depth. Almost 10 cm long desiccation fissures were observed at depths of 140 cm and 100 cm. Intercalations of massive silt and silty-clay are present at 100–50 cm and massive silty-sand with abundant root remnants represents the sediments of 50–0 cm depth.
Table 1 presents the 14C dates and calibrated values in the 2σ range. The maximum probability within the 2σ range of calibrated age vs. depth plot (Figure 4) provides the age models for both profiles. Age model of S1 pit is reconstructed from 4 different radiocarbon dates. The rate of sedimentation varies between 10.1 cm/kyr and 12.1 cm/kyr for sediments deposited during ca. 24.9–3.9 cal kyr BP (279–49 cm). The base of the sequence is assigned an age of ca.27 cal kyr BP by extrapolating the rate of sedimentation (10.5 cm/kyr) calculated between 205 and 279 cm. Sediments between 49 and 205 cm depth are assigned ages by interpolating the sedimentation rates calculated between consecutive 14C dates. We estimate sedimentation of 12.5 cm/kyr for the upper 49 cm of the sequence by assuming that the surface sediments represent present day. It is similar to the higher side of sedimentation rates calculated for S1 pit. Desiccation fissures at 75 cm depth indicate absence of any water body and dry conditions at the site of S1 pit at ca. 6.5 cal kyr BP.
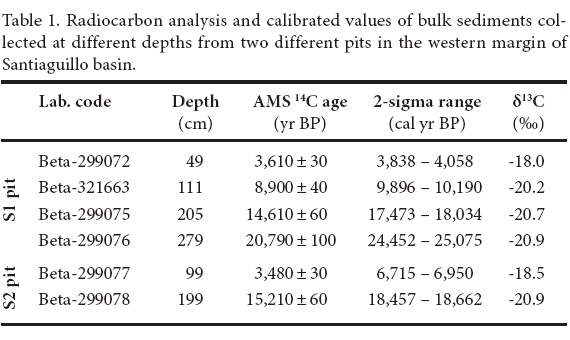
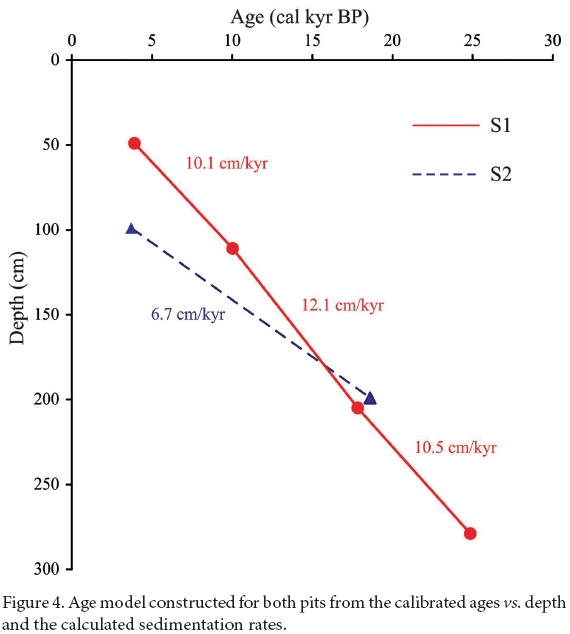
The age model of S2 pit includes 2 different calibrated 14C dates and assigns ca.19 cal kyr BP to the base. Assuming the surface sediments to represent the present day, a sedimentation rate of 26.7 cm/kyr was estimated for the upper 99 cm of the sequence. Estimated sedimentation rate for upper part of the sediment profile is very different from the sedimentation rate calculated between 199 and 99 cm depths (6.7 cm/kyr). The vertical fissures at 140 cm and 100 cm depths indicate that the site of S2 pit experienced multiple events of desiccation between ca.18.6 and 3.7 cal kyr BP. Lower sedimentation calculated between 199 and 99 cm could be due to the aeolian activity eroding the earlier deposited sediments during the dry intervals and lake desiccation. Due to limitations associated with both the age models, we did not undertake detailed paleoecological reconstruction occurred over last ca.3.9 cal kyr BP in S1 pit and last ca.3.7 cal kyr BP in S2 pit.
Ostracode species assemblages
The ostracode assemblages comprise 5 species (Figure 5). Except for one, the species were present as adults and juveniles. Low abundance and presence of only juvenile instars of one species precluded an accurate taxonomic identification. The paleoecological inferences are strictly based on the adult counts as the last ostracode moult takes place during the optimal environmental conditions (De Deckker, 2002). Two of the species belong to the genus Limnocythere (L. bradburyi Forester, 1985 and L. ceriotuberosa Delorme, 1967) and the other two are Eucandona cf. patzcuaro Tressler (1954) and Cypridopsis vidua Brady (1868). Most of the shells are well preserved and almost all are covered with authigenically precipitated carbonate minerals. Figures 6 and 7 summarize the total ostracode abundance (valves/g) and the relative abundance of each species (%) in both pits. Based on the variation in total ostracode abundance and relative abundances, S1 profile is divided into 5 different zones and S2 profile is divided into 3 different zones. The zones in both pits have comparable temporal distributions.
In the sediments of S1 pit, the total ostracode abundance varies between 0 and 125 valves/g (Figure 6). The juvenile shells are more frequent (up to 102 valves/g) than the adult shells (up to 57 valves/g). L. bradburyi (0–50 valves/g) is the most abundant species and it is followed by E. patzcuaro (0–9 valves/g). Both C. vidua (0–1 valves/g) and L. ceriotuberosa (0–1 valves/g) are the least abundant species. The zones are characterized by different total ostracode abundances and associations.
Sediments of zone 5 (300–265 cm, ca. 27–23.5 cal kyr BP) display a total ostracode abundance between 4 and 24 valves/g, and most of them are juveniles (up to 18 valves/g). This zone is characterized by highest species richness (4 species). L. bradburyi and E. patzcuaro constitute 69–100% and up to 31% of the adult ostracodes. Adult shells of L. ceriotuberosa are observed at depths of 299 and 281 cm. Similarly, one adult shell of C. vidua was observed at a depth of 299 cm. Sediments of zone 4 (265–220 cm, ca.23.5–19 cal kyr BP) shows the highest and variable ostracode abundance (8–125 valves/g) and most of them are juveniles (up to 102 valves/g). L. bradburyi and C. patzcuaro constitute 52–92% and up to 47% of the adult ostracodes. C. vidua is present in sediments at depths of 265–225cm. Zone 3 (220–105 cm, ca.19–9.5 cal kyr BP) displays total ostracode abundance between 2 and 113 valves/g. Both adults and juveniles are present in almost equal proportions. L. bradburyi represents 67–100% of the adult shells and E. patzcuaro constitutes up to 33% of the adult ostracodes. Sediments at depths of 185, 165–160 and 125–111 cm contain C. vidua. The depth interval 105–50 cm (ca.9.5–4 cal kyr BP) represents zone 2 and have a total ostracode abundance of 1–15 valves/g. This is monospecific and contains only L. bradburyi (100%). Zone 1 comprises the sediments at depths of 50–0 cm (last ca.4 cal kyr BP) and does not contain ostracodes.
The ostracode abundance is lower in the sediments of S2 pit compared to S1. Total ostracode abundance varies between 0 and 38 valves/g (Figure 7). Abundance of adult (up to 17 valves/g) and juvenile shells (up to 19 valves/g) are similar. Sediments of this pit do not contain L. ceriotuberosa, while L. bradburyi (0–17 valves/g) is the most abundant species. The abundances of E. patzcuaro (0–3 valves/g) and C. vidua (0–1 valve/g) are similar.
Sediments of zone 3 (200–135 cm, ca.19–9 cal kyr BP) have ostracode abundance of 3–38 valves/g. This zone is characterized by the presence of L. bradburyi, E. patzcuaro and C. vidua. L. bradburyi and E. patzcuaro display relative abundances of 83–100% and of 0–15%, respectively. Adult shells of C. vidua were observed at depths of 200–195 cm and 175–165 cm. Zone 2 (135–85 cm, ca.9–3.5 cal kyr BP) has an ostracode abundance of 3–12 valves/g and is characterized by the absence of C. vidua. L. bradburyi represents 80–100% of the adult shells and E. patzcuaro was observed at depths of 125 cm and 115 cm. Zone 1 comprises sediments deposited at depths of 85–0 cm (last ca.3.5 cal kyr BP) and lacks ostracode valves.
Total organic carbon (TOC) and organic carbon/nitrogen (C/N)
Concentrations of TOC and C/N in bulk sediments from both pits are presented in Figures 6 and 7. Sediments of S1 pit have TOC of 0.2–1.2%, total nitrogen (TN) of 0.02–0.10% and C/N values between 3.5 and 34 (Figures 6). Sediments rich in TOC generally contained lower number of ostracodes (i.e., 300–255 cm and 50–0 cm). Zones 5 and 4 (ca.27–19 cal kyr BP) have lower and uniform values of C/N (3.5–7). Sediments from zone 3 (ca.19–9.5 cal kyr BP) and zone 2 (ca.9.5–4 cal kyr BP) have higher and variable C/N (7–34). Zone 1 (last ca.4 cal kyr BP) have intermediate C/N values (8–16).
Sediments of S2 pit have TOC concentrations of 0–1.0%, total nitrogen contents of 0.09–0.16% and C/N values of 0–10 (Figure 7). TOC abundance is inversely related to total ostracode abundance. Sediments of all the 3 zones have similar values of C/N. Zone 3 (ca.19–9 cal kyr BP) has C/N values between 0 and 10 and zone 2 sediments (ca. 9–3.5 cal kyr BP) are characterized by C/N values of 2–10. Zone 1 sediments (last ca.3.5 cal kyr BP) have C/N values of 1–10.
DISCUSSION
Paleoecological reconstruction
Ostracodes preserved in the sediments of Santiaguillo basin tolerate almost similar ranges of temperature (5–32°C) and pH (8–10) (Delorme, 1989; Palacios-Fest, 2007, 2010). It was not possible to reconstruct the water column temperature in the absence of trace element geochemistry of ostracode valves and modern training sets in the area. However, the changing ostracode species assemblages suggest that the water column salinity varied between oligohaline and mesohaline over the last ca. 27 cal kyr BP. The salinity reconstruction is based on the classification proposed by Meisch (2000). Accordingly, the oligohaline waters are characterized by salinity range of 500–5,000 ppm and the mesohaline waters have 5,000–18,000 ppm of salinity. Except for the last ca. 3.5 cal kyr BP, the sediments preserve 4 different ostracode species and L. bradburyi is the most abundant. Forester (1985) identified L. bradburyi in different shallow oligohaline to euhaline lakes from central Mexico. We relate the highest abundance of L. bradburyi to the presence of shallow water bodies at the western margin of Santiaguillo between ca. 27 and 3.5 cal kyr BP. Palacios-Fest (2010) relates the occurrence of E. patzcuaro to intervals of fresh to oligohaline water as a result of increased fresh water inflow into the basin. Similarly, the presence of C. vidua was observed in warm (>13°C) and fresh to oligohaline lakes in southwestern USA (Palacios-Fest, 2010). L. ceriotuberosa tolerates oligohaline to mesohaline conditions in USA and Canada (Delorme, 1971a, 1971b, 1971c; Palacios-Fest, 2007). In the interior plains of Canada, Smith and Delorme (2010) observed that L. ceriotuberosa was present in waters with low dissolved oxygen content (3 mg/L).
The paleoecological conditions of Santiaguillo are divided into pre-LGM, LGM and post-LGM. The interpretations are supported by TOC and C/N data. TOC content in bulk sediment reflects the amount of primary productivity (Cohen, 2003) and C/N ratio reflects the contribution of terrestrial vegetation and lacustrine algae on the deposited organic matter (Meyers and Ishiwatari, 1995). Lacustrine algae and terrestrial plants have different contents of nitrogen and the increasing contribution of one of them to the organic matter is reflected by the C/N ratio. Sediment with higher C/N (>10) represents dominant contribution from terrestrial plants and lower C/N (<10) indicates that the organic matter originated from aquatic plants (Talbot and Johannessen, 1992; Meyers and Ishiwatari, 1995; Cohen, 2003).
Pre-LGM (ca.27–23.5 cal kyr BP): Sediments of S1 pit (zone 5, 300–265 cm) documented the paleoecological conditions that occurred prior to the last glacial maximum (pre-LGM). The ostracode assemblage suggested presence of shallow water column and oligohaline conditions. Presence of a permanent water body in the western margin of the basin is indicated by lacustrine phytoplankton (C/N<10) dominantly contributing to the organic productivity, as well as the continuous increase in ostracode productivity. Occurrence of both C. vidua and L. ceriotuberosa indicated relatively low salinity at ca.27 cal kyr BP. The absence of C. vidua during rest of the interval suggested a slight increase in salinity in the oligohaline range. L. ceriotuberosa at 299 cm and 281 cm depths represented the intervals of less oxygenated water column (e.g. Smith and Delorme, 2010).
LGM (ca.23.5–19 cal kyr BP): Paleoecological condition of the last glacial maximum (LGM) was recorded by the lower part of S1 pit (zone 4, 265–220 cm). Existence of the shallow and permanent water body continued during this interval and the organic productivity was contributed by the lacustrine phytoplankton (i.e. C/N≤10). The ostracode assemblage of L. bradburyi, E. patzcuaro and C. vidua suggested the presence of a warm (>13°C) and more diluted oligohaline water body. Total ostracode abundance was higher at ca.20–19 cal kyr BP (60–125 valves/g) compared to ca.23.5–20 cal kyr BP (8–35 valves/g). The inverse relationship between total ostracode abundance and TOC could be due to the result of ostracode dissolution in an acidic environment formed by the organic carbon decomposition.
Post-LGM (last ca.19 cal kyr): Sediments of zone 3 (220–105 cm) and zone 2 (105–50 cm) of S1 pit documented the paleoecological conditions that occurred post the last glacial maximum (post-LGM). This interval was also represented by sediments of zones 3 and 2 (200–85 cm) of S2 pit. Ostracodes of S1 pit represented ca.19–4 cal kyr BP and ostracodes of S2 pit indicated the paleo-hydrochemical characteristics occurred during ca.19–3.5 cal kyr BP. The ostracode assemblages suggested presence of oligohaline to mesohaline water bodies in the western margin.
Post ca.19 cal kyr BP, the decreasing abundance of E. patzcuaro and increasing C/N values reflected a shift in the paleoecological conditions. Higher C/N values (>10) indicated that the terrestrial vegetation contributed more to the organic matter as the lacustrine productivity decreased. Gradual reduction in the amount of pluvial discharge into the basin was reflected by the decreasing abundance of E. patzcuaro. More salinity (Na and HCO3-Cl enriched water body) was reflected by an increase in the abundance of L. bradburyi (e.g. Forester, 1985). Absence of both E. patzcuaro and C. vidua in the sediments of ca.13–11.5 cal kyr BP indicated less fresh water inflow into the basin and more salinity (mesohaline) during the Younger Dryas (YD). However, the reappearance of both the species in sediments of the Pleistocene-Holocene transition suggested presence of relatively diluted oligohaline water column. Based on the concentrations of Ti and K in bulk sediments from another core in Santiaguillo, Roy et al. (2014) also identified a humid interval during the Pleistocene-Holocene transition and related it to enhanced summer precipitation in the region. During ca.9.5–4 cal kyr BP, the water body was ephemeral and was possibly present only during the summer months. Ostracode abundance gradually decreased, while the concentration of TOC increased. The occurrence of only L. bradburyi at 111–50 cm depths in S1 pit suggests presence of mesohaline water body in the western margin of the basin. Arid conditions caused enhanced aeolian activity in the surroundings of Santiaguillo during ca. 9.3–4.3 cal kyr BP (Roy et al., 2014a). Similarly, the desiccation fissures at 75 cm depth indicate that the location of S1 pit remained dry for a longer duration at ca.6.5 cal kyr BP.
Although both the pits are located in the western margin of the basin, the sediments collected from them have different characteristics. Compared to sediments of S1 pit, the sediments of S2 pit are characterized by lower total ostracode abundance (up to 38 valves/g), C/N relation (<10) and rate of sedimentation (6.7 cm/kyr). Total ostracode abundance (up to 70 valves/g), C/N relation (>10) and sedimentation rate (10.1–12.1 cm/kyr) are higher in sediments of S1 pit. The desiccation fissures at 140 cm and 100 cm depths and lower sedimentation rate suggest that the site of S2 pit experienced multiple events of desiccation and erosion of previously deposited sediments during ca.18.6–3.7 cal kyr BP. The arid intervals might have caused oxidation of organic matter and poor preservation of ostracode valves in S2 pit.
Sediments representing the last ca. 4 cal kyr BP in S1 pit and last ca.3.5 cal kyr BP in S2 pit do not contain ostracodes. Sediments of this interval are relatively coarser (i.e. fine sand) compared to the lower parts (i.e. clay, silt, silty-sand and silty-clay). Over the last ca. 3.5 cal kyr BP, the amount of sediments deposited at the location of S2 pit (99 cm) is almost twice of the sediment deposited at S1 site (<50 cm). This was possibly due to the higher inflow into the site of S2 pit. The difference in the amount of inflow into two different sites in the western margin is also reflected by the C/N values indicating a terrestrial source for the organic matter deposited at S1 pit and lacustrine source for the organic matter deposited at S2 pit.
Comparison with regional records
Comparison of the paleoecological information from Santiaguillo with other paleoclimatic records from northern and northwestern Mexico (see Figure 1a for locations) suggests that the region experienced different climatic and hydrological conditions over the last ca.27 cal kyr BP. Paleohydrological and paleoclimatic information are available from Babicora (Metcalfe et al., 2002; Roy et al., 2012b), El Fresno and Santa María (Castiglia and Fawcett, 2006) from Chihuahua, San Felipe (Lozano-Garcíaet al., 2002; Roy et al., 2010, 2012a) and Chapala (Davis, 2003) from Baja California, Purísima-Iray-Magdalena from Baja California Sur (Murillo de Nava et al., 1999) and Las Cruces from San Luis Potosi (Roy et al., 2013b). Between ca.27 and 19 cal kyr BP, the salinity of Santiaguillo basin was lower compared to the salinity of water column at Babicora. The ostracode species assemblage suggested presence of oligohaline water body at Santiaguillo and oligohaline to mesohaline water bodies at Babicora. Multielement geochemistry of the sediments indicated reduced runoff into Babicora and San Felipe basins (Roy et al., 2010, 2012a, 2012b). During this interval, the vegetation in the surroundings of Babicora comprised Pinus and Picea (Metcalfe et al., 2002) and vegetation in the surroundings of San Felipe basin were Abies, Pinus and Eryngium (Lozano-García et al., 2002).
Over the last ca.19 cal kyr BP, the runoff into Santiaguillo gradually reduced and the water body became more saline. However, Babicora received more precipitation post ca.18 cal kyr BP (Roy et al., 2013b) and the runoff into San Felipe increased after ca.13 cal kyr BP (Roy et al., 2010). The formation of linear and mega-barchan dunes indicate hot and arid conditions at the Purísima-Iray-Magdalena basin during ca.14–8 kyr BP (Murillo de Nava et al., 1999). During the early to middle Holocene, the Santiaguillo sediments indicated mesohaline conditions (ca.9.5–4 cal kyr BP), enhanced aeolian activity (ca.9.3–4.3 cal kyr BP) and multiple events of desiccation. In general, both Babicora and San Felipe received more runoff over the Holocene. However, Babicora experienced an event of desiccation at ca.3 cal kyr BP and San Felipe sediments documented more aeolian activity at ca.8 cal kyr BP. Laguna El Fresno and Laguna Santa María experienced wet conditions and higher lake stands at ca.9.4–9.3 cal kyr BP, ca.7.5–7.2 cal kyr BP and ca.4.9–4.2 cal kyr BP (Castiglia and Fawcett, 2006). The pluvial discharge into Las Cruces was more during ca.8.4–5 cal kyr BP compared to the last ca.5 cal kyr (Roy et al., 2013b). A permanent lake was present in the Chapala basin till ca.8.4 cal kyr BP and dune formation commenced post 7.5 cal kyr BP (Davis, 2003). At Purísima-Iray-Magdalena, the dune activity was stabilized and the present lagoon system was developed at ca.6–5 kyr BP.
CONCLUSIONS
Several multi-proxy studies have attempted to reconstruct the late Pleistocene-Holocene paleoecological and paleoclimatic conditions of subtropical Mexico. However, the information is less from the central and southern parts compared to the northern part. Similarly, ostracodes have received limited attention so far compared to the study of sediment geochemistry and palynology. In the present study, we intend to improve the paleoecological information of the central-southern part of subtropical Mexico by studying the abundances and species associations of 4 different ostracode species in combination with TOC and C/N data in sediments deposited over the last ca.27 cal kyr BP in the Santiaguillo basin. More specifically;
Ostracode assemblage consists of 4 species (Limnocythere bradburyi, Eucandona cf. patzcuaro, Cypridopsis vidua and Limnocythere ceriotuberosa) and the highest (125 valves/g) abundance occurred during the LGM.
During pre-LGM (ca.27–23.5 cal kyr BP), the western margin of the basin hosted shallow and oligohaline water body and experienced dominant lacustrine productivity. Changes in the abundances of C. vidua and L. ceriotuberosa indicated relatively lower salinity at ca.27 cal kyr BP compared to ca.27–23.5 cal kyr BP.
L. bradburyi, E. patzcuaro and C. vidua suggested presence of warm (>13°C) and more diluted oligohaline water body during the LGM (ca.23.5–19 cal kyr BP).
Lower abundance of E. patzcuaro and higher C/N ratio reflected a shift in the paleoecological conditions at ca.19 cal kyr BP. Both the pluvial discharge into the basin and lacustrine productivity decreased. Higher abundance of L. bradburyi suggested that the water column became more saline.
The water body was mesohaline during ca.9.5–4 cal kyr BP and the absence of ostracode shells in sediments of last 3.5 cal kyr BP was possibly caused by ephemeral conditions and periodic desiccation.
Comparison with other proxy-records from the region suggests that the paleohydrological and paleoclimatic changes occurred over the last ca.27 cal kyr BP in different parts of sub-tropical northern Mexico were not synchronous.
ACKNOWLEDGEMENTS
The data presented in this paper were generated with the financial support from DGAPA-PAPIIT project IN100413. CMCL acknowledges the scholarship from CONACYT for her postgraduate studies. SEM pictures were obtained from the Central Microscopy Laboratory of the Institute of Physics (UNAM). Technical assistance comes from José Luis Sánchez Zavala and Nayeli Lopez Balbiaux. Dr. S. Srinivasalu, Fernando Nuñez and David Quiroz participated in the field work. We are thankful to the staff of Heróico Cuerpo de Bomberos and Protección Civil (Durango) for providing assistance and security during the expedition. The comments and suggestions from Angel Baltanás and an anonymous reviewer improved the interpretations.
REFERENCES
Barron, J.A., Metcalfe, S.E., Addison, J.A., 2012, Response of the North American monsoon to regional changes in ocean surface temperature: Paleoceanography, 27, PA3206, doi: 10.1029/2011PA002235. [ Links ]
Brady, G.S., 1868, A synopsis of the recent british Ostracoda: Intellectual Observer, 12, 110-130. [ Links ]
Caballero, M., Peñalba, M.C., Martínez, M., Ortega-Guerrero, B., Vázquez, L., 2005, A Holocene record from a former coastal lagoon in Bahía Kino, Gulf of California, Mexico: The Holocene, 15(8), 1236-1244. [ Links ]
Carreño, A.L., 1990, Ostrácodos lacustres del paleolago de Texcoco, México: Revista de la Sociedad Mexicana de Paleontología, 3(1), 117-129. [ Links ]
Castiglia, P.J., Fawcett, P.J., 2006, Large Holocene lakes and climate change in the Chihuahuan Desert: Geology, 34(2), 113-116. [ Links ]
Chávez-Lara, C.M., Roy, P.D., Caballero, M.M., Carreño, A.L., Lakshumanan, C., 2012, Lacustrine ostracodes from the Chihuahuan Desert of Mexico and inferred Late Quaternary paleoecological conditions, Mexico: Revista Mexicana de Ciencias Geológicas, 29(2), 422-431. [ Links ]
Cohen, S.A., 2003, Paleolimnology: the history and evolution of lake systems, E.U.A.: Oxford University Press, 500 pp. [ Links ]
COHMAP members, 1988, Climatic change of the past 18,000 years: observations and model simulations: Science, 241, 1043-1052. [ Links ]
Davis, L.G., 2003, Geoarcheology and geochronology of pluvial Lake Chapala, Baja California, Mexico: Geoarcheology, 18, 205-223. [ Links ]
De Deckker, P., 2002, Ostracod palaeoecology, in Holmes, J.A., Chivas, A.R. (eds.), The Ostracoda: Applications in Quaternary research: Washington, American Geophysical Union, Geophysical Monograph Series, 131, 121-134. [ Links ]
Delorme, L.D.., 1967, New freshwater Ostracoda from Saskatchewan, Canada: Canadian Journal of Zoology, 45(3), 357-363, 2 plates. [ Links ]
Delorme, D.L., 1971a, Freshwater ostracodes of Canada. Part II, Subfamily Cypridopsinae and Herpetocypridinae, and family Cyclocyprididae, Canada: Canadian Journal of Zoology, 48, 253-266. [ Links ]
Delorme, D.L., 1971b, Freshwater ostracodes of Canada. Part III, Family Candonidae, Canada: Canadian Journal of Zoology, 48, 1099-1127. [ Links ]
Delorme, D.L., 1971c, Freshwater ostracodes of Canada. Part V, Families Limnocytheridae, Loxoconchidae, Canada: Canadian Journal of Zoology, 49, 43-64. [ Links ]
Delorme, D.L., 1989, Methods in Quaternary ecology #7: Freshwater ostracodes, Canada: Geoscience Canada, 16, 85-90. [ Links ]
Forester, R.M., 1985, Limnocythere bradburyi n. sp.: a modern ostracode from central Mexico and a possible Quaternary paleoclimatic indicator: Journal of Paleontology, 59(1), 8-20. [ Links ]
Forester, R.M., 1986, Determination of the dissolved anion composition of ancient lakes from fossil ostracodes: Geology, 14(9), 796-799. [ Links ]
García, A.E., 1973, Modificaciones al Sistema de Clasificación Climática de Köppen: México,, Universidad Nacional Autónoma de México, D.F.,Boletín del Instituto de Geografía, 246 pp.
Holmgren, C.A., Peñalba, M.C., Rylander, K.A., Betancourt, J.L., 2003, A 16,000 14C yr B.P. packrat midden series from the USA-Mexico Borderlands: Quaternary Research, 60, 319-329. [ Links ]
Holmgren, C.A., Norris, J., Betancourt, J.L., 2007, Inferences about winter temperatures and summer rains from the late Quaternary record of C4 perennial grasses and C3 desert shrubs in the northern Chihuahuan Desert: Journal of Quaternary Science, 22(2), 141-161. [ Links ]
Kirby, M.E., Lund, S.P., Bird, B.W., 2006, Mid-Wisconsin sediment record from Baldwing Lake reveals hemispheric climate dynamics (Southern CA, USA): Palaeogeography, Palaeoclimatology, Palaeoecology, 241, 267-283. [ Links ]
Kutzbach, J.E., Wright, H.E., 1985, Simulation of the climate of 18,000 yr B.P.: Results for North America/North Atlantic/European Sector: Quaternary Science Reviews, 4, 147-187. [ Links ]
Kutzbach, J., Gallimore, R., Harrison, S., Behling, P., Selin, R., Laarif, F., 1998, Climate and biome simulations for the past 21,000 yr: Quaternary Science Reviews, 17, 473-506. [ Links ]
Lozano-García, M.S., Ortega-Guerrero, B., Sosa-Nájera, S., 2002, Mid to Late Wisconsin pollen record of San Felipe basin, Baja California: Quaternary Research, 58, 84-92. [ Links ]
Manabe, S., Broccoli, A.J., 1985, The influence of continental ice sheet on the climate of an ice age: Journal of Geophysical Research, 90, 2167-2190. [ Links ]
Meisch, C., 2000, Freshwater Ostracoda of western and central Europe, in Schwoerbel, J. Zwick, P. (eds), Süßwasserfauna von Mitteleuropa 8(3): Spektrum Akademischer Verlag GmbH, Heidelberg, Berlin, 1-522. [ Links ]
Metcalfe, S.E., Bimpson, A., Courtice, A.J., O'Hara, S.L. and Taylor, D.M., 1997, Climate change at the monsson/ Westerly boundary in Northern Mexico: Journal of Paleolimnology, 17, 155-171. [ Links ]
Metcalfe, S.E., Say, A., Black, S., McCulloch, R. O'Hara, S., 2002, Wet Conditions Turing the Last Glaciation in the Chihuahuan Desert, Alta Babicora basin, Mexico: Quaternary Research, 57, 91-101. [ Links ]
Meyers, P.A, Ishiwatari, R., 1995, Organic matter accumulation records in lake sediments, in Lerman, A., Imboden, D., Gat J. (eds.) New York: Physics and Chemistry of Lakes: Springer-Verlag, 279-328. [ Links ]
Morkhoven, F.V., 1963, Post-palaeozoic Ostracoda: Their Morphology, Taxonomy, and Economic Use Vol. II, Generic Descriptions: Elsevier, 478 pp. [ Links ]
Murillo de Nava, J.M., Gorsline, D.S., Goodfriend, G.A., Vlasov, V.K., Cruz-Orozco, R., 1999, Evidence of Holocene climatic changes from aeolian deposits in Baja California Sur, Mexico: Quaternary International, 56, 141-154. [ Links ]
Nieto-Samaniego, A.F., Barajas-Gea, C.I., Gómez-González, J.M., Rojas, A., Alaniz-Álvarez, S.A., Xu, S., 2012, Geología, evolución estructural (Eoceno al actual) y eventos sísmicos del Graben de Santiaguillo, Durango, México: Revista Mexicana de Ciencias Geológicas, 19(1), 115-130. [ Links ]
Ortega-Guerrero, B., Caballero Miranda, M., Lozano García, S., De la O Villanueva, M., 1999, Palaeoenvironmental record of the last 70 000 yr in San Felipe basin, Sonora desert, Mexico: preliminary results, México: Geofísica Internacional, 38(3), 1-11. [ Links ]
Ortega-Rosas, C.I., Peñalba, M.C., Guiot, J., 2008, Holocene altitudinal shifts in vegetation belts and environmental changes in the Sierra Madre Occidental, northwestern Mexico, based on modern and fossil data: Review of Palaeobotany and Palynology, 151, 1-20. [ Links ]
Palacios-Fest, M.R., 2007, Superfamily Cytheroidea: an assisting tool for identifying ostracode species and their basic ecological requirements: Terra Nostra Earth Sciences Research, LLC and California State University, Poster available online in www.docstoc.com/docs/117353499 and CSUB Department of Physics and Geology. [ Links ]
Palacios-Fest, M.R., 2010, Late Holocene Paleoenvironmental History of the Upper West Amarillo Creek Valley at Archaeological Site 41PT185/C, Texas, USA, México: Boletín de la Sociedad Geológica Mexicana, 62(3), 399-436. [ Links ]
Palacios-Fest, M.R., Carreño, A.L., Ortega-Ramírez, J.R., Alvarado-Valdéz, G., 2002, A paleoenvironmental reconstruction of Laguna Babícora, Chihuahua, Mexico based on ostracode paleoecology and trace element shell chemistry: Journal of Paleolimnology, 27(2), 185-206. [ Links ]
Pérez, L., Lorenschat, J., Massaferro, J., Pailles, C., Sylvestre, F., Hollwedel, W., Brandorff, G.O., Brenner, M., Islebe, G., Lozano, M.S. , Scharf, B., Schwalb, A., 2013, Bioindicators of climate and trophic state in lowland and highland aquatic ecosystems of the northern Neotropics: Revista de Biología Tropical, 61(2), 603-644. [ Links ]
Reimer, P.J., Baillie, M.G.L., Bard, E., Bayliss, A., Beck, J.W., Blackwell, P.G., Bronk Ramsey, C., Buck, C.E., Burr, G.S., Edwards, R.L., Friedrich, M., Grootes, P.M., Guilderson, T.P., Hajdas, I., Heaton, T.J., Hogg, A.G., Hughen, K.A., Kaiser, K.F., Kromer, B., McCormac, G., Reimer, R.W., Richards, D.A., Southon, J.R., Talamo, S., Turney, C.S.M., van der Plicht, J., Weyhenmeyer, C., 2009, IntCal09 and Marine09 radiocarbon age calibration curves, 0-50,000 years cal BP: Radiocarbon, 51, 1111-1150. [ Links ]
Roy, P.D., Caballero, M., Lozano, R., Ortega, B., Lozano, S., Pi, T., Morton, O., 2010, Geochemical record of Late Quaternary paleoclimate from lacustrine sediments of paleo-lake San Felipe, western Sonora Desert, Mexico: Journal of South American Earth Sciences, 29(3), 586-596. [ Links ]
Roy, P.D., Caballero, M., Lozano, S., Morton, O., Lozano, R., Jonathan, M.P., Sánchez, J.L., Macías, M.C., 2012a, Provenance of sediments deposited at paleolake San Felipe, western Sonora Desert: implications to regimes of summer and winter precipitation during last 50 cal kyr BP: Journal of Arid Environments, 81, 47-58. [ Links ]
Roy, P.D., Jonathan, M.P., Pérez-Cruz, L.L., Sánchez-Córdoba, M.M., Quiroz-Jiménez, J.D., Romero, F.M., 2012b, A millennial-scale late Pleistocene-Holocene palaeoclimatic resgister from western Chihuahua Desert, Mexico: Boreas, 41, 707-717. [ Links ]
Roy, P.D., Quiroz-Jiménez, J.D., Pérez-Cruz, L.L., Lozano-García, S., Metcalfe, S.E., Lozano-Santacruz, R., López-Balbiaux, N., Sánchez-Zavala, J.L., Romero, F.M., 2013a, Late Quaternary palaeohydrological conditions in the dry lands of northern Mexico: a summer precipitation proxy record of the last 80 cal kyr BP: Quaternary Science Reviews, 78, 342-354. [ Links ]
Roy, P.D., Rivero-Navarette, A., Lopez-Balbiaux, N., Pérez-Cruz, L.L., Metcalfe, S.E., Muthu Sankar, G., Sánchez-Zavala, J.L., 2013b, A record of Holocene summer-season palaeohydrological changes from the southern margin of Chihuahua Desert (Mexico) and possible forcings: The Holocene, 23, 1105-1114. [ Links ]
Roy, P.D., Quiroz-Jiménez, J.D., Chávez-Lara, C.M., Sánchez-Zavala, J.L., Pérez-Cruz, L.L., Muthu Sankar, G., 2014a, Humid Pleistocene-Holocene transition at sub-tropical northern Mexico and possible Gulf of California forcing: Boreas, 43, 577-587. [ Links ]
Roy, P.D., Charles-Polo, M.P., López-Balbiaux, N., Pi-Puig, T., MuthuSankar, G., Lozano-Santacruz, R., Lozano-García, S., Romero, F.M., 2014b, Last glacial hydrological variations at the southernmargin of sub-tropical North America and a regional Comparison: Journal of Quaternary Science, 29(5), 495-505. [ Links ]
Servicio Meteorológico Nacional (SMN), México. Estación 10137 Guatimape (DGE) (24°48'25" N., 104°55'19" W., 1,974.0 msnm). Normales Climatológicas 1951-2010. <http://smn.cna.gob.mx/climatologia/Normales5110/NORMAL10137.TXT>, August, 2012 [ Links ]
Smith, A.J., Delorme, L.D., 2010, Chapter 19, Ostracoda, in Thorp J.H., Covich A.P. (eds.) Ecology and Classification of North American Freshwater Invertebrates: London, Academic Press, 725-772. [ Links ]
Talbot, M.R., Johannessen, T., 1992, A high resolution palaeoclimatic record for the last 27,500 years in tropical West Africa from the carbon and nitrogen isotopic composition of lacustrine organic matter: Earth and Planetary Science Letters, 110(1), 23-37. [ Links ]
Thompson, R.S., Anderson, K.H., 2000, Biomes of western North America at 18,000, 6,000 and 0 14Cyr BP reconstructed from pollen and packrat midden data: Journal of Biogeography, 27, 555-584. [ Links ]
Tressler, W.L., 1954, Fresh-water Ostracoda from Texas and Mexico: Journal of the Washington Academy of Science, 44, 138-149. [ Links ]
Turpen, J.B., Angell, R.W., 1971, Aspects of molting and calcification in the ostracod Heterocypris: The Biological Bulletin, 140(2), 331-338. [ Links ]
Urrutia-Fucugauchi, J., Ortega-Ramirez, J., Cruz-Gatica, R., 1997, Rock magnetic study of late Pleistocene-Holocene sediments from the Babicora lacustrine basin, Chihuahua northern Mexico: Geofísica Internacional, 10, 77-86. [ Links ]

