










Servicios Personalizados
Revista
Articulo
 Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista mexicana de ciencias geológicas
versión On-line ISSN 2007-2902versión impresa ISSN 1026-8774
Rev. mex. cienc. geol vol.31 no.2 Ciudad de México ago. 2014
Study of Cedral Horses and their place in the Mexican Quaternary
Estudio de los caballos de Cedral y su lugar en el Cuaternario mexicano
María Teresa Alberdi1, Joaquín Arroyo-Cabrales2, Alejandro H. Marín-Leyva3, and Oscar J. Polaco2†
1 Departamento de Paleobiología, Museo Nacional de Ciencias Naturales, CSIC, José Gutiérrez Abascal, 2, 28006 Madrid, España.* malberdi@mncn.csic.es
2 Laboratorio de Arqueozoología "M. en C. Ticul Álvarez Solórzano", Moneda 16, Col. Centro, 06060 México, D. F., Mexico.
3 Universidad Michoacana de San Nicolás de Hidalgo, Morelia, Michoacán, Mexico.
Manuscript received: July 24, 2013
Corrected manuscript received: April 11, 2014
Manuscript accepted: April 28, 2014
ABSTRACT
A detailed study has been undertaken with an unique horse bone deposit at Cedral, San Luis Potosí, central Mexico. Morphological and morphometrical characters are used, as well as bivariate and multivariate statistics for both cranial and postcranial elements, and additional data incorporated for specimens from other Pleistocene Mexican localities. Measurements for most of the studied materials are provided, as well as estimates of body mass for each species. Three species are represented in several Mexican late Pleistocene deposits, coincident with the Rancholabrean Land Mammal Age. All three may have been contemporaneous: a large-sized horse Equus mexicanus Hibbard, 1955 known from the western USA throughout Mexico and Central America; a widespread medium-sized horse Equus conversidens Owen, 1869 occurring in most of North and Central America; and a new small-sized horse Equus cedralensis sp. nov., presently known only from Mexican localities. Recognizing the co-occurrence of three late Pleistocene horse species (genus Equus sp.) in Mexico is important for understanding the diversity and extinction patterns at the time of the early presence of humans in the continent. Additionally, environmental inferences are proposed, but further research is warranted to test those.
Key words: taxonomy; Equus; new species, Late Pleistocene; Mexico; Cedral.
RESUMEN
Se realizó un estudio detallado de un depósito único de huesos de caballo en Cedral, San Luis Potosí, centro de México. Se usaron caracteres morfológicos y morfométricos, así como análisis estadísticos bivariantes y multivariantes de los restos del cráneo y del esqueleto postcraneal y se compararon con restos de otras localidades del Pleistoceno mexicano. Se suministran las medidas del material estudiado así como la estimación de la masa corporal de cada una de las especies. Tres especies de caballo están representadas en varios depósitos del Pleistoceno superior de México, correspondientes a la edad de mamíferos Rancholabreana, los cuales pueden haber sido contemporáneos: un caballo de gran tamaño Equus mexicanus Hibbard, 1955 conocido desde la porción occidental de Estados Unidos de América hasta México y América Central; un caballo de tamaño mediano ampliamente distribuido Equus conversidens Owen, 1869 que se encuentra en la mayor parte de América del Norte y Central; y un nuevo caballo de pequeño tamaño Equus cedralensis sp. nov., conocido hasta ahora sólo en localidades mexicanas. El conocimiento de la presencia conjunta de estas tres especies en el Pleistoceno tardío de México (género Equus sp.) es importante para entender los modelos de diversidad y extinción en los primeros tiempos de la presencia humana en el continente. Adicionalmente, se proponen algunas inferencias ambientales, pero se requerirá de más estudios para ponerlas a prueba.
Palabras clave: taxonomía; Equus; especie nueva; Pleistoceno Tardío; México; Cedral.
INTRODUCTION
The family Equidae Gray, 1821 is a monophyletic group first recorded from the Early Eocene of North America. This group shows a remarkable phylogenetic and adaptive radiation from the Miocene to Pleistocene, when the most derived cursorial and grazer types are developed (Simpson, 1951; MacFadden and Hulbert, 1988; MacFadden, 1992). This radiation occurs throughout all continents except Australia.
Fossil horse taxonomy has been complicated worldwide, and that is exactly the case for the species occurring in Pleistocene deposits of Mexico. Historically, fossil horse taxonomy in Mexico starts with the occurrence (Owen, 1869) and subsequent description based exclusively on photographs, of two species of fossil horse for the Basin of Mexico, Equus conversidens Owen, 1869 and E. tau Owen, 1869. More than 25 species have been reported from Mexico, including 13 names with type locality occurring in Mexico, most of which were only described based on the occlusal surface variation of the molars, and are currently under the synonymy of other species (see Álvarez and Ocaña, 1987 for a synthesis of the available information at the time, but also Winans 1985, 1989).
Most recent studies include reports on the horse remains from two localities. The first is the Irvingtonian-Rancholabrean Arroyo Cedazo, Aguascalientes, from where some of those 13 names were applied to specimens from the deposit. A detailed study by Reynoso-Rosales and Montellano-Ballesteros (1994), however, has concluded that only one species is present in the deposit, Equus conversidens. They allocate most of the previously reported variation to the species wide variation (contra Schafer and Dalquest, 1991). The other study involves specimens from the Rancholabrean locality at Cedral, San Luis Potosí. Based on bi- and multivariate analyses, at least two valid species have been recognized, with the possibility of an unnamed third one, assigning the names E. mexicanus Hibbard, 1955 for the large-sized species, and E. conversidens for the medium-sized (Alberdi et al., 2003). Based on the same specimens studied by Alberdi et al. (2003), but also with others from Arroyo Cedazo and Tequixquiac, and ANOVA analyses, Melgarejo-Damián and Montellano-Ballesteros (2008) support similar conclusions for Cedral specimens. A third study (Pichardo, 2004, 2006) is anecdotal and based mainly on reviewing the available literature from all over the Americas, but mostly focused on a specific region, Valsequillo, Puebla, with the use of restricted data for comparison with literature data. Although a new species is described, the study is poorly supported by the currently available data. Furthermore, both publications are almost identical.
Since the former study by our team at Cedral (Alberdi et al., 2003), we have added large samples from the late Pleistocene Mexican localities: Valsequillo, Arroyo Cedazo, and Tequixquiac. The aim of this paper is to study in detail the taxonomic position of horses from Cedral, setting a standard for Pleistocene horse studies in Mexico.
STUDY SITE: GEOGRAPHY AND STRATIGRAPHY
Rancho La Amapola is located 1.7 km SE of Cedral, San Luis Potosí, Mexico (Figure 1). It is at 23o49'N, 100o43'W at an altitude of 1,700 m. In general, the area has several Pleistocene fossiliferous deposits resulting from the presence of now dry springs that used to form a lake in the past, as recently as the XIX Century (Lorenzo and Mirambell, 1986, 1999). The present scrubland vegetation is distinct from the one that existed in the past. Based on the known past fauna, more humidity allowed the existence of a grassland (e.g., the meadow vole Microtus pennsylvanicus (Ord, 1815) - Álvarez and Polaco, 1982; the Columbian mammoth Mammuthus columbi Falconer, 1857- Arroyo-Cabrales et al., 2003; and the terrestrial mollusks Pupilla syngenes (Pilsbry, 1890a), Grastrocopta pellucida hordeacella (Pilsbry, 1890b), and Helicodiscus singleyanus (Pilsbry, 1890c) -Olivera-Carrasco, 2012), or even a tropical deciduous forest (Tapirus haysii Leidy, 1860 - Arroyo-Cabrales et al., 1996).
From 1977 to 1984, interdisciplinary excavations were undertaken at Rancho La Amapola through the "Endorreic Basins" project under the direction of the personnel of the Departamento de Prehistoria (currently Subdirección de Laboratorios y Apoyo Académico, Instituto Nacional de Antropología e Historia (INAH)). The project main objective was to learn in depth about human settlements at the site from the late Pleistocene to the Early Holocene with an environmental perspective. Faunal studies were reported either as an overall listing (Álvarez and Polaco, 1981) or as specific taxonomic studies (Álvarez and Polaco, 1982; Arroyo-Cabrales, 1992; Arroyo-Cabrales et al., 1996). Recently, a monograph on the site (Mirambell, 2012) had a chapter dedicated to the faunal remains (Álvarez et al., 2012).
At the site, several hearths were found, accounting for a 15,000-16,000 year temporal range, the oldest was found in Layer XII, and dated more than 35,000 years BP (37,694 ± 1,963 years BP, Lorenzo and Mirambell, 1999), the most recent dated at 21,468 ± 458 years BP (Layer X). Several faunal horizons were found in the deposit. Those layers had abundant and generally split bone remains, mostly isolated and quite disperse. However, a unique large horse bone concentration was uncovered between layers X and XII (Figure 2).
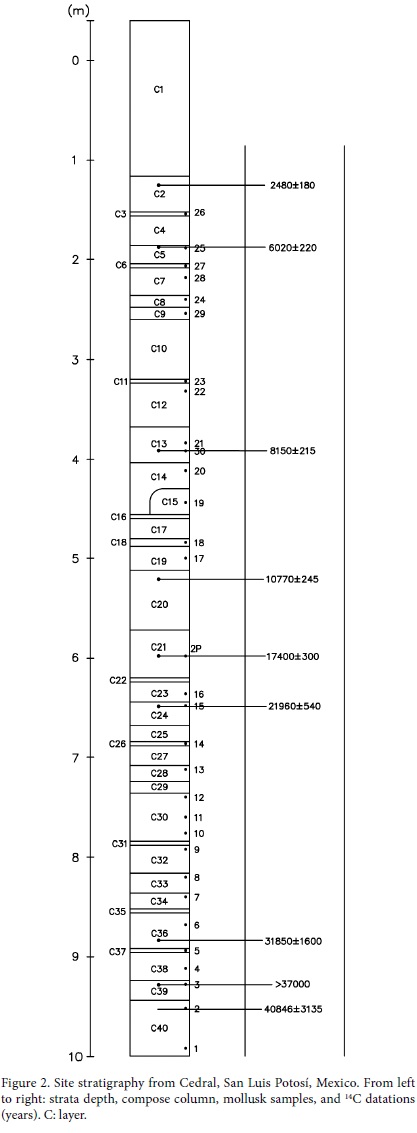
MATERIAL AND METHODS
The fossil remains studied belong to the large collection of horse bone remains from Cedral archaeological site (San Luis Potosí, Mexico) on deposit at the Paleontological Collection (DP-INAH) of the Laboratorio de Arqueozoología "M. en C. Ticul Álvarez Solórzano" Subdirección de Laboratorios y Apoyo Académico, INAH in Mexico City. This collection is very rich and contains a good representation of different parts of the skeleton and several remains of horse skulls and jaws. With exception of the medium-sized horse group, the large-sized and small-sized groups are very well represented by long bones belonging to the zeugo- and stylopodium (humeri, radii, femora, tibia), metapodials, phalanges, carpal and tarsal bones, and vertebrae. Skulls and mandibles are scarce and fragmentary. The morphology is similar in the three groups of horses and they significantly vary in size.
The skull and maxillary remains with teeth are: one symphysis; eight P2-M3 right and left rows; plus one P2-P4 right, one P3-M2 left, one P4-M3 right, one P3-M1-M3 right, one P3-M3 left, one P2-M2 left, and three P2-M1 right and left, and one juvenile; isolated teeth: eight P2, three right and five left; 31 P3-4,15 right and 16 left; 48 M1-2, 25 right and 23 left; 27 M3,15 right and 12 left; the several upper deciduous or milk teeth and incisors not included in the analyses because they are not significant at the taxonomical level. The mandible remains with teeth are: one symphysis; six mandible fragments with complete lower rows (p2-m3), and two p2-p4, three p3-m3, one p2-m1, and two m1-3 (right and left); and several lower isolated teeth: 34 p2, 16 right and 18 left; 46 p3-4, 31 right and 25 left; 16 m1-2, six right and 10 left; 25 m3, 11 right and 14 left; the lower deciduous teeth and incisors not included in this study (Appendix 1, electronic supplement).
The postcranial skeleton is represented by abundant bones from all of the skeleton elements, generally with very good conservation; several vertebrae and ribs are not included in our analysis because they do not contribute for identification at the taxonomic level. Other bones include: eight scapula fragments, five right and three left; 16 humeri fragments (two complete), six left and four right, plus six without determination; 26 radii fragments (seven complete), 14 left and 11 right, plus one without determination; two oleocranon fragments, one right and one left; 33 magnums, 16 right and 17 left; 46 scaphoids, 29 right and 17 left; 24 lunatum, 11 right and 13 left; 18 pyramidals, six right and 12 left; four trapezoid, one right and three left; nine unciforms, six right and three left; 12 pisiforms, four right and left; among the metacarpals (MC), seven MCII, two right and five left; 24 MCIII (20 complete), 14 right and 10 left; 10 MCIV, five right and five left; five pelvic fragments, only the acetabulum area, one right and four left; 11 femuri fragments (one complete), three right and five left; 29 tibia fragments (three complete), seven right and 15 left, plus seven without determination; one rotula large in size; 25 calcanei fragments (12 completes), 12 right and 13 left, plus 13 without determination; 67 astragali (55 complete), 33 right and 34 left; 31 tarse naviculars, 14 right and 17 left; seven ectocuneiforms, two right and five left; 22 cuboids, nine right and 13 left; among the metatarsals (MT), five MTII, one right and four left; 28 MTIII (25 completes), 14 right and 11 left; 15 MTIV, four right and 11 left; among the phalanges (PH), without distinction between front part and back part, 100 1PHIII (88 complete), 108 2PHIII (91 complete), and 34 3PHIII, mostly incomplete; also 41 proximal sesamoids and four distal sesamoids (Appendix 2, electronic supplement).
Morphological and morphometric features of the equid dental and skeleton remains from Cedral have been compared with those of Equus remains from several localities. Those remains included in both bivariate and multivariate analyses are Arroyo Cedazo, Aguascalientes (CED) (stored at the Museo de Paleontología, Instituto de Geología, Universidad Nacional Autónoma de México (UNAM)); Tequixquiac, State of Mexico (TEQ), stored at DP-INAH, and Museo Geológico del Seminario (Barcelona); Valsequillo, Puebla (VAL) stored at DP-INAH and C-INAH Puebla. The Valsequillo equid remains are from different localities considered as part of the Valsequillo paleontological complex Tepenextec, Caulapan, Zacachimalpa, Tetela, Arenillas, Xochitenango, La Mata, Atepetzingo, and Alseseca.
Morphological and morphometrical characters and bivariate and multivariate analyses were executed with the metrical parameters. Those analyses and comparisons were mainly based on teeth and limb bones because they are the most abundant remains for statistical purposes, and with more diagnostic characters. The several maxillae and mandibles were insufficient for statistical analysis. The hypsodonty index was calculated on unworn or only slightly worn teeth (P3-4/p3-4 and M1-2/m1-2) as H/L ratio, where H is the maximal tooth height, and L the maximal length at 1 cm of the base of the crown.
Robustness or slenderness index of third metacarpals (MCIII) and metatarsals (MTIII) were calculated as minimal breadth at the middle of the diaphysis versus maximal length (MCIII3 x 100/ MCIII1; MTIII3 x 100/ MTIII1). We added another slenderness index of metapodial in regard to the distal maximal articular breadth (MCIII11 x 100/ MCIII1; MTIII11 x 100/ MTIII1), following Bernor et al. (1990). We also included the body mass estimation, following Alberdi et al. (1995) methodology, using the variable MCIII13 rather than 1PHIII5, due to the large variability of this last variable for this group of horses where the differences between anterior and posterior phalanges are quite strong.
Bivariate and multivariate analyses were performed to estimate variation and differences in size and proportions. Metacarpals, metatarsals, astragali, calcanei, first and second phalanges (MCIII, MTIII, AST, CAL, 1PHIII, 2PHIII), the most characteristic bones of the appendicular skeleton. We analysed them using principal component analysis (PCA) to explore and evaluate similarities or differences in size among the Equus remains from Cedral and the Mexican localities in order to assess their relationship with equids from other localities. Then, discriminate analysis (DA) was performed using variable independents all together or Mahalanobis methods on groups previously identified by PCA. This was executed to maximize the separation between the groups already recognized, as well as for evaluating whether the groups differ significantly or not among them. The results of PCA enabled us to group the remains by differences in size, while DA puts out how significant the differences are among three groups.
The PCA and DA were performed on 23 MCIII, 40 MTIII, 64 AST, 18CAL, 112 1PHIII, and 118 2PHIII of Equus from Cedral plus the comparative remains from the late Pleistocene Mexican localities. Teeth were not utilized for these analyses because their measures change throughout their life due to their wear, biasing any possible differences that were found. For description, morphological analysis, nomenclature, and measurements, we followed the recommendations of the "Hipparion Conference" (New York, November 1981) (Eisenmann et al., 1988). All dimensions were taken in millimeters.
Calculations were made using SPSS 15.0. For more information about the multivariate analyses see Marcus (1990) and Reyment (1991).
RESULTS
Description
The remains of fossil equids found in Cedral do not show any significant differences as the main morphological traits of teeth among them. The dimensions and proportions indicate the presence of three groups with similar features, but differences in size, as previously indicated by Alberdi et al. (2003). In regard to limb bones the differences in size are also large, and dimensions and proportions also indicate the presence of three groups with similar features but large differences in size and slenderness. The middle-sized group is poorly represented, while the remains of the larger and smaller-sized groups are very well represented.
Morphology and size
A complete skull is lacking, and all of the teeth have much cement. The main morphological feature of the upper cheek teeth (premolars and molars) is the elongate protocone, more distal than mesial with a slight groove lingual and sub-triangular in shape on P3-P4, with a more elongate subtriangular outline on M1-M2; the pli caballin is generally present (Figure 3). The enamel fossette plications are scarce in general in the three different sizes of horses. Parastyle and mesostyle, in general, are wide in premolars with grooves, and narrow in the molars without grooves. The hypocone is generally open and triangular in shape, with marked distal grooves and slightly marked hypoconal constriction. The grooves are more developed on the unworn teeth and nearly lost on the strongly worn ones. The hypocone is isolated inside the loph on some M3. The hypsodonty index is middle-high, ranging from 2.66 to 3.33 ( =2.94) in large size horses, and from 2.92 to 3.21 (=3.07) in small size horses on P3-4 (no premolar middle in size without wear); on M1-2 (no middle in size without wear) the hypsodonty index ranging from 3.09 to 3.5 (=3.24) in large size horses and from 2.99 to 4.2 (=3.47) in small size horses.
=2.94) in large size horses, and from 2.92 to 3.21 (=3.07) in small size horses on P3-4 (no premolar middle in size without wear); on M1-2 (no middle in size without wear) the hypsodonty index ranging from 3.09 to 3.5 (=3.24) in large size horses and from 2.99 to 4.2 (=3.47) in small size horses.
Those morphological features are observable as well on the occlusal surface of the upper cheek teeth rows. According to the tooth size, right and left rows are: P2-M3 length between 189.5-195 large size, and between 144-148.5 small size; P2-P4 length 94.9-105.0 large size, 77.3-81 middle size, and 63.4-74 small size; M1-M3 length between 85.8-89.3 large size, and 63-68 middle size. There are not remains of a skull (Table 1). Measures of the isolated upper check teeth are in Appendix 1 of the electronic supplement and Figure 4.
Several incomplete mandibles and all of the teeth show much cement. The occlusal morphology of the lower premolars and molars is of a caballin type, characterized by an elongate double knot with rounded metaconids and more or less angular outline metastylids, and elongate linguaflexids deeper on molars than on premolars. The ectoflexid on premolars is shallow and does not enter the isthmus, while it is deeper in molars and sometimes it penetrates the isthmus, joining sometimes the linguaflexid in much worn molars. The ectoflexids often show a small pli, better marked in premolars than in molars, and quite evident on the unworn teeth (Figure 3). Premolars are larger than molars. The hypsodonty index, calculated on p3-4 and m1-2 unworn cheek teeth, ranges from 2.54 to 3.3 in p3-4 (=3.03) large in size, 2.54 middle in size, and 2.94 small in size, and between 2.85 to 3.86 in m1-2 (=3.48) large in size, 2.85-3.4 (=3.08) middle in size, and 2.89-3.49 (=3.19) small in size. These values are consistent with the field of variation of this group. Molars are slightly more hypsodont than premolars, both upper and lower teeth, but no morphological differences among the three groups of horses based on size. Premolars and molars of Cedral mandibles (Figure 3) show the same caballin occlusal morphology as in isolated teeth. The scarce remains of the complete mandibles are characterized by a mandibular corps high and robust and the muzzle short and large. According to their size, the maximal length of p2-m3 varies between 181.5-196.2 for large size, 156.6 for middle size, and 140.2 small size; p2-p4 length between 91.3-104.0 for large size, 83-85.97 for middle size, and 73 for small size; and m1-m3 length between 85.4-93.0 for large size, 73.4 for middle size, and 68.3-69.3 for small size (Table 1). Morphological differences occur in P2/p2 and M3/m3 due to the larger variability of those teeth in equids. Measurements of the isolated lower check teeth are in Appendix 1.
Remains of the appendicular skeleton are well represented, and mostly belong to the larger and smaller sizes (Figure 5), as shown in the multivariate analysis. Fragmentary remains of scapula and pelvis, mostly consisting of glenoid articular surface and acetabulum, respectively, do not provide any sound taxonomic information; complete remains of humerii, radii and ulnae, femuri, tibiae, metapodials, phalanges, carpal and tarsal bones occur. Among other skeleton remains, the most important are metapodials, phalanges, and several carpal and tarsal bones because they play a very important role in the movement and in the transmission of the body weight. The data about the body mass are shown in each group, where the three different sizes are clear. Measurements of the skeleton bones are in Appendix 2 of the electronic supplement.
Statistical analysis
The bivariate analyses of length versus width of the upper (P3-4 and M1-2) and lower (p3-4 and m1-2) cheek teeth at the base of the crown (to 1 cm), as well as length/protocone length ratio in upper teeth and length/double knot (formed by metaconid-metastylid) ratio and length/liguaflexid ratio in lower teeth separate the equid remains from Cedral in three groups of different size. The spatial distribution of these teeth from Cedral overlaps with the comparative remains of El Cedazo, Valsequillo and Tequixquiac remains of large and small size, respectively. The bivariate analyses minimal breadth (near the middle of the bone) on the MCIII and MTIII vs. maximal length on MCIII and MTIII, and distal maximal articular breadth on the MCIII and MTIII vs. maximal length on MCIII and MTIII were done following Bernor et al. (1990) (Figure 6).
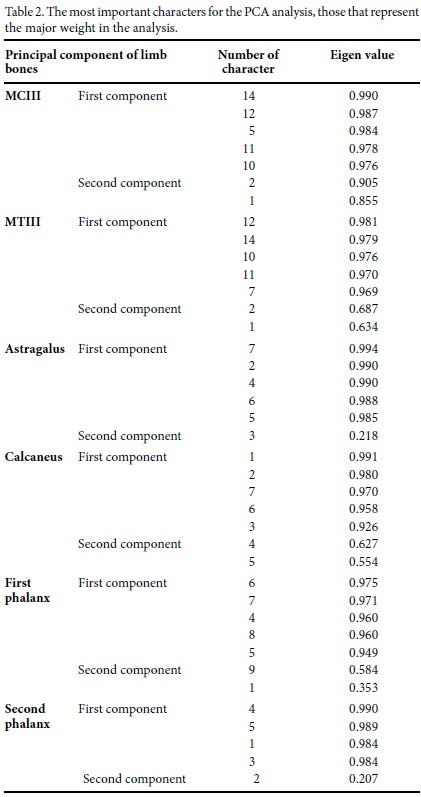
The PCA of MCIII grouped the different remains from Cedral site in three groups separated by size, along with different individuals from Arroyo Cedazo, Valsequillo and Tequixquiac. The three first components explain 97.6% of the variance. The MTIII PCA results are very similar to those for MCIII, both in grouping two secondary groups in the distribution of the small size remains from the Mexican localities. The three first components explain 96.5% of the variance (Figure 7). Also, the PCA results for the first and second phalanges of the third digit (1PHIII and 2PHIII) indicate very similar distributions among different groups and their relations with other mexican remains included here for comparison. The three first components explain 97.4% and 99.2% of the variance, respectively (Figure 7). The astragali and calcanei PCAs show three groups, each clearly differentiated and similar in size relationships to the other bones, the calcanei are not numerous. The three first components explain 98.9% and 97.1% of the variance, respectively. The most important characters for the PCA, those that had the major weights in the analyses, are indicated in Table 2.
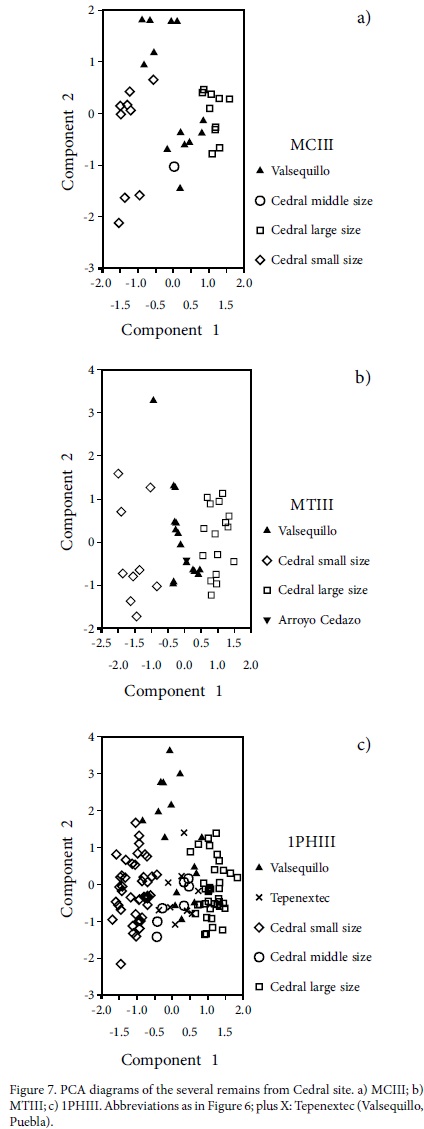
Based on the results of PCA grouping different size associations provides a global view concerning the size of the remains. The remains of different mexican horses are grouped together to execute a discriminant analysis (DA). Specifically, a group occurs with remains of a large size form; a middle size from different localities, which are scarce; and a small size form with a wide variation.
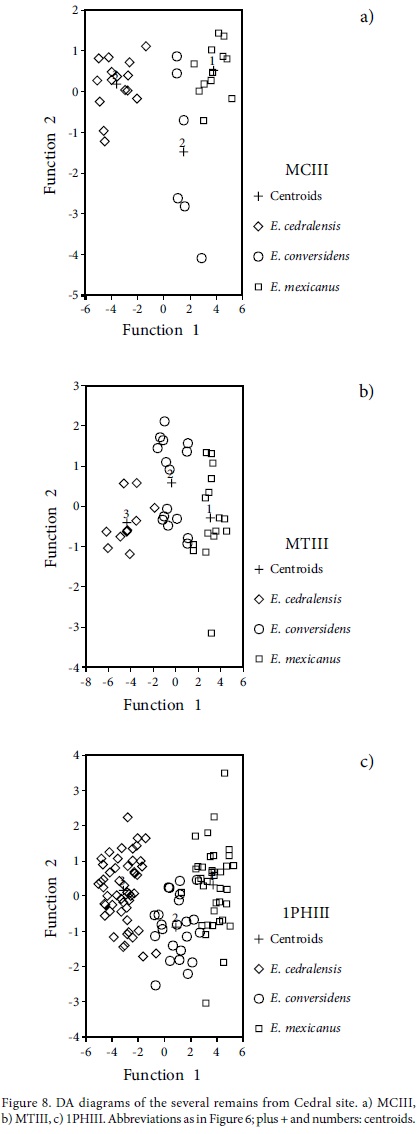
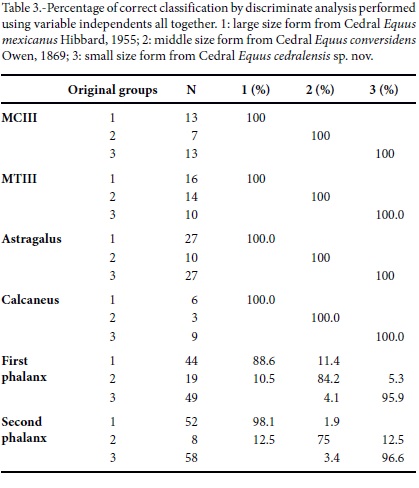
The DA results, both by using variable independents all together or Mahalanobis methods, give very similar results. DA MCIII results, based on different groups resulting from the PCA analysis, indicate that a 100.0% and 97.0% of original grouped cases are correctly classified, respectively. Either the small size subgroups are considered separated or joined. The DA MTIII results are the same those for the MCIII, based on the different groups resulting from the PCA analysis, indicating that 100% and 97.5% respectively of original grouped cases are correctly classified. In the Mahalanobis method the large and middle size are 100% correctly classified, and the small size group is correctly classified in 90.0% of original grouped cases (Figure 8; Table 3). The 1PHIII DA results, based on different groups resulting from the PCA analysis, indicate a 91.1% and 93.8% of original grouped cases correctly classified, respectively for both methodologies of DA. The small size group is very well classified in 95.9% and 93.9% of the original grouped cases, respectively (Figure 8; Table 3). The 2PHIII DA results, based on different group resulting of the PCA analysis, indicate a 95.8% and 94.1% of original grouped cases correctly classified, respectively. The small size group is very well classified in 96.6% and 93.1% respectively of original grouped cases (Figure 8; Table 3). The astragali are well classified in 98.4% of the original grouped cases by Mahalanobis method, and 100% by using variable independents together. The middle size and the small size groups are correctly classified in 100% of original grouped cases (Table 3). The calcanei are very scarce and they are all correctly classified using both methodologies.
SYSTEMATIC PALAEONTOLOGY
The results of the analyses indicate the presence of three horse sizes in the Cedral and other studied localities: one large in size, a second middle in size, overlaps with the remains of Equus conversidens from other late Pleistocene Mexican localities and a third small in size. Those Mexican remains are smaller than other small-size remains of horses that inhabited North America (USA and Canada).
Order Perissodactyla Owen, 1848
Family Equidae Gray, 1821
Subfamily Equinae Gray, 1821
Tribu Equini Gray, 1821
Genus Equus Linnaeus, 1758
Equus mexicanus Hibbard, 1955
(Figures 6 to 8)
Synonyms. Equus occidentalis Leidy, 1865; Equuspacificus Leidy, 1868; Equus scotti Gidley, 1900
Holotype. Hibbard (1955) described as: a nearly perfect skull lacking the LM3 (No. 48 (HV-3) stored at Museo Nacional de Historia Natural. More recently Carranza-Castañeda and Miller (1987) rediscovered this specimen, mentioned that the skull now has the nasal bones broken and the incisors missing; the specimen is cataloged as IGM4009.
Type locality. Tajo de Tequixquiac, Estado de México.
Geographical distribution. E. mexicanus is cited at different localities in Mexico, United States of America and Central America.
Stratigraphic distribution. Rancholabrean local faunas from the Pleistocene.
Age and geographic range. Late Rancholabrean, known from the middle-western United States of America and Central America.
Diagnosis. The skull is large. The basicranial length is greater than 500 mm at the available material. The narial notch is deep and extends over the commissure between P2 and P3. The rostrum is broad, the choanae rather narrow, the posterior palatine foramina are placed opposite the M3. The masseteric ridge is of medium length and curves downward rostrally. The upper cheek teeth are large, with elongate flattened protocones, derived features and average complexly plicate enamel. Lower molars have a shallow ectoflexid on premolars, penetrating the isthmus on m1, m2, and m3 in advance stage of wear. Following Winans (1989), the ratio of length to proximal width is generally less than 5.0 for metacarpals and 6.0 for metatarsals. Metapodials are not stilt-legged. The body mass from MCIII13 is between 354.33 and 532.88 kg and the mean is 456.23 kg.
Study material. See Table 1 and Appendices 1 and 2 of the electronic supplement.
Discussion. The dental morphology from Cedral is very similar to the morphology from the other two identified species, while the size varies significantly. The discriminant analyses indicate that the remains assigned to E. mexicanus are large in size and body mass and also they present a morphological and external anatomical body shape that we indicate above. Hibbard (1955) described and figured E. mexicanus from Tajo de Tequixquiac, State of Mexico, Mexico, differentiated by the skull morphology, mainly by the skull occipital region being pointed out at the bottom, more than any other known Pleistocene or Holocene species; ventrally, the premaxilla is mostly flat in comparison with the bend shown by other species, and included as synonyms to E. excelsus Leidy, 1858 and E. occidentalis Leidy, 1865. The availability of larger series of Pleistocene horse skulls have shown that those characters mentioned by Hibbard (1955) are more common than expected; the same is shown for recent specimens with some characters, like occiput orientation and premaxilla shape, varying with age (Osborn, 1912).
Lundelius (1972) mentioned that there are small differences in the lower and upper teeth between E. mexicanus and E. pacificus; one is based on a more complex folding of the fossetes in E. pacificus Leidy, 1868, than in E. mexicanus. This is not a diagnostic character, tooth morphology becomes more and more simpler with wear. Kurtén and Anderson (1980) include E. mexicanus as a synonym of E. giganteus Gidley, 1901, and different from E. scotti Gidley, 1900, they consider it may be referred to the subgenus Amerhippus Hoffstetter, 1950. Also, Lundelius (1972) includes E. mexicanus as a synonym of E. pacificus, while Savage (1951) considered it as a nomen nudum.
Winans (1989) consideres E. mexicanus as a valid species and included it in her E. laurentius group. At present E. laurentius Hay, 1913, is not considered a fossil, some indication that it must be considered as a synonym of horse E. caballus Linnaeus, 1758 (Scott et al., 2010). E. mexicanus represents the larger end of the prehistoric horse spectrum. Its fossil remains are found in United States, at Rancho La Brea living in grasslands. The western horse is grazer. Some evidence suggests that the species underwent a small, yet significant reduction in body size near the end of the last Ice Age. This change in body size may be the result of changing conditions in both climate and vegetation (Guthrie, 2003).
Dalquest and Schultz (1992) stated that if E. mexicanus and E. scotti are the same species, then E. mexicanus must be the synonym of E. scotti. Azzaroli (1995) proposed E. mexicanus as a synonym of E. pacificus, but later himself (Azzaroli, 1998) considered E. pacificus as a synonym of E. mexicanus (because E. pacificus is also based on a unique tooth), and E. scotti as a synonym of E. excelsus.
For Azzaroli (1998) the systematic position of E. mexicanus is a puzzle. The peculiar features of the profile and the occipital region have been observed only in species from South America, in E. fraternus Leidy, 1860, and possibly in E. conversidens. Bennett (1980) proposed that the ventral rotation of the occiput was a primitive feature, found in Dinohippus Quinn, 1955. In fact, a strong rotation is shown in Osborn's figure of the type skull of Dinohippus leidyanus Osborn, 1918 (Protohippus leidyanus in the original text: Osborn, 1918, pl.30), but the fossil is deformed. Better preserved skulls of the same species, from Nebraska and from Kansas, show backward sloping paraoccipital processes and lack any trace ofventral rotation of the occiput (Azzaroli, 1998). The specific character of E. mexicanus, out of those from South American and, possibly the North American species of Equus (E. fraternus and E. conversidens), is a derived, not a primitive feature.
This discussion clearly shows the diversity of criteria in regard to the large-sized North American horses in more than 100 years, including the complex nomenclatural issues that may be in agreement for some authors and in total disagreement by others. Because of that, it was decided to assign the large-sized Mexican horse remains from Cedral and other localities to E. mexicanus, pending a detailed and exhaustive review of all of the known species of North American Pleistocene horses to define valid and non-valid species, and the relationships among them.
Equus conversidens Owen, 1869
(Figures 3 to 8)
Holotype. MNM-403, fragmentary right maxilla with all cheek teeth.
Type locality. Valley of Mexico, Mexico. Carranza-Castañeda and Miller (1987) rediscovered this specimen, represented by a palate with complete cheek tooth dentition, assigned the catalog number IGM4008.
Geographical distribution. North America, Mexico and Central America.
Stratigraphic distribution. Pleistocene, probably Rancholabrean.
Diagnosis. The skull fragment IGM4008 (Holotype; No. 403 old number) is constituted mostly by the rostral portion, with the maxilla and teeth in situ. Each dental series includes three molars and three premolars. The premolars exceed the true molars in size (pl. LXI, fig. 1; Owen, 1869). The upper cheek teeth are middle in size, with elongate flattened protocones, derived features and average complexly plicate enamel similar to E. mexicanus but smaller in size. Lower molars have similar morphology but smaller in size than E. mexicanus teeth. Following Winans (1989), the ratio of length to proximal width is generally less than 5.0 for metacarpals and 6.0 for metatarsals. Metapodials are not stilt-legged. The body mass from MCIII13 is between 215.63 to 282.52 kg and the mean is 249.08 kg.
Owen (1869) figured an illustration based on a photograph of a specimen lost; Carranza-Castañeda and Miller (1987) reported the rediscovery of the specimen in the collections of the Instituto de Geología at Universidad Nacional Aautónoma de México; doing so they published a photograph from the ventral portion of the specimen.
Study material. See Table 1 and Appendices 1 and 2 as supplementary information.
Discussion. Owen (1869) described the specimen, a portion of the right and left series of grinders, and a considerable part of the intervening bony plate, based on a photograph. Owen (1869) gave the name Equus conversidens to the species because the tooth rows of the type specimen rostrally converge. The specimen was damaged and has been restored, so that the validity of Owen's observation was questioned by some authors (see discussion in Gidley, 1901); facial skull from the late Pleistocene deposits at Barranca del Muerto near Tequixquiac (LACM 308/123900) clearly showed this feature. They proposed that the photograph actually shows two different specimens, each corresponding to one maxillar side with different enamel patterns and molar lengths. Because of this, Gidley (1901) named as the species lectotype the right maxilla. Other than the mentioned convergence, the type specimen not has unique characters and differences between both ramii are within the normal variability for the genus.
A rich collection, including practically all the bones of the skeleton but disarticulated, comes from San Josecito Cave, Nuevo León, Mexico, LACM loc.192. Stock (1953) based his new subspecies E. conversidens leoni Stock, 1953 on these fossils that included two skulls from cave. A third skull comes from Papago Springs Cave, Arizona, and was described by Skinner (1942). All the skulls are broken and largely restored, so that they are not reliable for a definition of the characters of this species. The skull from Barranca del Muerto, although incomplete, is more diagnostic. The strongly tapering snout and the low position and small size of the orbit are close to E. fraternus; however, the pre-orbitale foramen is placed high. Hibbard (1955) considered that E. tau is a synonym of E. conversidens. The description of the leoni subspecies by Stock (1953) indicates that the differences between the type species and this subspecies are minimal or non-existent because they correspond to variable characters.
Most authors have agreed with E. conversidens name. Winans (1985) indicates that it must be considered as nomen dubium because the original description uses a non-existent character; in 1989 she includes the subspecies leoni nomen nudum in the E. alaskae; Azzaroli (1995, 1998) in E. francisci Hay, 1915. Our comparative analysis on the skeleton remains indicates that E. conversidens and middle-sized remains from Mexican localities are significantly larger than E. francisci, and do not have stilt-legged metapodials.
The nomenclatural position of the medium-size horse with only moderately slender metapodials (ratio of the length to the proximal width of the metacarpal generally less than 5.0 and that of the metatarsal less than 6.0) has been quite complex. Winans (1985) treated E. conversidens as a nomen nudum, it was named solely on the basis of dental characters. The name has been widely used. At times, the name E. alaskae Hay, 1913 has been used in the literature based on the treatment of species groups by Winans (1989).
Azzaroli (1998) indicates that the geological range of E. conversidens was Irvingtonian throughout Rancholabrean according to Kurtén and Anderson (1980). All of the specimens recorded in that book are of Rancholabrean age, as well as the specimens discussed in this paper. Azzaroli (1995) considers E. fraternus closely related to the South American species of Equus, and (Azzaroli, 1998) also indicated: "Like Equus fraternus, Equus conversidens is closely related to South American horses. It seems however too young to be their direct ancestor." We consider that all horses from the late Pleistocene (Racholabrean or Lujanian) are very similar morphologically, with small differences that correspond to adaptative considerations on the distal appendicular skeleton.
Equus nobisi Pichardo, 2004 was described based on a left mandibular fragment from Arenillas, Puebla, currently on deposit at the collection at Centro INAH Puebla (Mexico) as number CRINAHP 1305. The specimen was originally reported by Nobis (1973) as part of Nobis' group II, he related his group I with E. mexicanus and his group III with E. cf. conversidens, but group II was said not to be related to any known species. Pichardo (2004) described the new species, based solely on Nobis (1973) data, but only provided some characters as part of the comparisons between the different species from the site. Pichardo (2008) included this specimen from the greater Valsequillo complex as E. excelsus. Furthermore, Pichardo (2000, 2004, 2006) emphasized the use ofzoogeographic range and econiche preference as valid taxonomic characters, and that those should be given taxonomic weight in species diagnosis. Clearly those concepts are away from current biological tendencies in regard to the species concept (De Queiroz, 2007).
We have chosen to retain the name E. conversidens for this medium-sized, extinct horse while acknowledging that Winans (1989) may be nomenclaturally correct. The finding of the specimen (Carranza-Castañeda and Miller, 1987) whose photograph was utilized by Owen (1869) to describe the species, should counteract Winans'proposal.
Equus cedralensis new species
(Figures 3 to 9)
Holotype. A mandible ramus fragment formed by two specimens: one p2-m3 right row (DP-2675 I-2 15), and a second fragment of the symphysis with the anterior dentition (DP-2674 I-2 8), articulated together and stored at the Paleontological Collection (DP-INAH) of the Laboratorio de Arqueozoología "M. en C. Ticul Álvarez Solórzano" Subdirección de Laboratorios y Apoyo Académico, INAH in Mexico City (Figure 9).
Paratype series. Fragments of mandible ramii with remains of several teeth, p3-m3 right and left (DP-4577) and several upper row: P2-M2 left (P2 DP-3841, P3-P4 DP-3842, and M1-M2 DP- 3839), and three P2-M1 (DP-3829 left, and DP-3830 and DP-4587 right).
Type locality. "Rancho La Amapola" located 1.7 km southeastern from Cedral, San Luis Potosí, Mexico. Coordinates: 23°49'N, 100°43W, 1700 m a.s.l.
Derivation name. From the Cedral locality where the type specimens were found.
Geographical distribution. Different late Pleistocene localities inside Mexico, other than Cedral: Valsequillo (Puebla), Tequixquiac (State of Mexico), Arroyo Cedazo (Aguacalientes) and possibly from La Cinta-Portalitos and La Piedad-Santa Ana (Michoacán).
Stratigraphic distribution. Rancholabrean, late Pleistocene.
Diagnosis. One of the smallest North American horse species for late Pleistocene, larger than Equus tau Owen. Although no complete skulls or mandibles, several upper and lower teeth rows (P2/p2-M3/m3), very small in size and some of the mandible rami are almost complete like the holotype. Upper and lower cheek teeth are morphologically very similar to the other horses found at Cedral, but smaller in size. The upper teeth have a protocone elongate more distal than mesially and with a small lingual groove; with styles, parastyle, and mesostyle on premolars wide with a groove and narrow without grooves on molars. The lower teeth have metaconid-metastylid (double knot), rounded or slightly angular, respectively. The ectoflexid is superficial in premolars and molars without crossing the isthmus, only on molars with an advanced wear the ectoflexid are deeper, crosses the isthmus, and could touch the linguaflexid; the linguaflexid is large and more or less U shape. The symphysis has a linear arcade with large canines (a possible male). The body mass is between 91.83 and 169.59 kg, and the mean is 138.41 kg following Alberdi et al. (1995) on the MCIII13. The appendicular skeleton is also small in size with metapodials not stilt-legged, clearly different from the other remains from Cedral and other Mexican localities. In fact, the Mexican new species has metapodials shorter and less slender than both francisci and alaskae.
Study material. See Table 1 and Appendices 1 and 2 for supplementary information.
Discussion. Data for this horse was compared with the different data for small horses from North America to find the possible relationship with those remains. Winans (1989) proposed as small-sized horses those of E. francisci and E. alaskae groups, the first with stilt-legged metapodial, and the latter with stout-legged. The small horse remains from Mexican localities are smaller than E. francisci and E. alaskae groups; the metapodials are clearly smaller and less slender than those two groups; metacarpals show some overlapping with Winans (1989) E. alaskae (an ongoing study of the North American horses and their comparisons with the specimens shows that both E. francisci and E. alaskae were larger than E. cedralensis sp.nov.). Winans (1985) considers E. francisci as a valid species and included as synonyms E. altidens Quinn, 1957, E. arellanoi Mooser, 1958, E. calobatus Troxell, 1915, E. quinni Slaughter et al., 1962, and E. zoyatalis Mooser, 1958. Hay (1915) declared E. francisci to be the size of E. tau, but differing from it in having the transverse width of P3-M1 greater than the anteroposterior length. This difference is typical of teeth that are in different stages of wear and does not signify taxonomic difference. Hibbard and Taylor (1960) have E. francisci as a synonym of E. conversidens. Slaughter (1966) suggested an intergradation between E. francisci, E. conversidens, E. lambei Hay, 1917, and E. fraternus. Lundelius and Stevens (1970) presented evidence based on similar specimens found in Texas for maintaining E. francisci as a separate species. The species E. francisci will be maintained because it is the oldest name assigned to the stilt-legged group. If either E. conversidens or E. tau can be shown to be valid and to belong to this group, then those names will take precedence.
In the case of Equus alaskae it was elevated to species rank from E. niobrarensis alaskae Hay, 1913 by Winans (1989), who included within the E. alaskae group as synonyms: E. conversidens leoni, E. lambei and E. niobrarensis alaskae. Winans (1989) referred to E. alaskae group all specimens from San Josecito Cave, Natural Trap Cave, and small horses from Dry Cave. Typical E. niobrarens is larger than E. alaskae with a narrower premaxillae, no clear-cut characters separated these horses from other species.
Azzaroli (1998) included E. tau among the synonyms of E. francisci, Hibbard (1955) included E. tau within E. conversidens, Winans (1985) considered E. lambei not a fossil, that it could be a synonym of E. caballus or E. asinus Linnaeus, 1758. Winans(1989) included it as synonyms of E. alaskae group. Dalquest (1979) considered that North American and Mexican species of horses, only valid: E. conversidens and E. tau (includes E. francisci as a synonymous for E. tau). Based on the data for E. tau provided by Mooser and Dalquest (1975) and Dalquest (1977, 1979), dental measurements were smaller than those for E. cedralensis sp. nov. from Cedral, except few molars that overlap. Metapodials for E. cedralensis are much shorter (lenght MCIII = 181-224 mm, MTIII = 233-271 mm) than the same for E. tau (length of MCIII 235 mm, MTIII between 277-285 mm; see Table 4), and less slender. An-ongoing study for reviewing the whole North American late Pleistocene species should be the small-sized species.
For the PCA the smallest metapodials split into two subgroups with differences in gracility more considerably noticeable in MCIII than in MTIII. But in DA, these differences between both subgroups are diminished. Vera Eisenmann (personal communications August 4, 2012) deduces from the metapodials small in size the presence of three different horses: the majority of MCIII small in size and two MTIII correspond to E. quinni, three MTIII correspond to E. francisci and only two MCIII correspond to E. cedralensis sp. nov. The metapodi-als identified by V. Eisenmann as E. francisci and E. quinni present in Cedral are shorter than the data that exist in the literature for E. francisci and E. quinni (see Slaughter et al., 1962; Lundelius and Stevens, 1970). The possible existence of many species of horses in this locality could be surprising. Good representation exists of the majority of the bones in the skeleton and teeth. Only in the metapodials is a large variation while in other bones only three sizes were detected. Average ratios (length, dimension 1, to proximal width, dimension 5) for studied metacarpals and metatarsals are more variable than in E. alaskae and E. francisci (after Winans, 1989), with some specimens closer to E. alaskae and others to E. francisci. The Cedral group is significantly smaller than the remaining horses analysed by Winans (1985, 1989). The francisci group encompasses all horses with stilt-legged metapodials, having a ratio of length to proximal width generally greater than 5.0 for metacarpals and 6.0 for metatarsals (Winans, 1989). The differences between both species are linked to the metapodials being larger and slender in francisci than in alaskae, but both are more slender that the Mexican new species. After Winans (1989) the size range from largest to smallest metapodials is greater in francisci group than that for most other groups. This situation raises the possibility that the group should be subdivided, but until more information becomes available, it is best to support these specimens as a single group.
DISCUSSION
Horse taxonomy is convoluted worldwide and the same is true for Mexican horses. Many species have been described, many of which are currently synonyms of others. Although some studies have tried to assess the actual taxonomic status of the North American Pleistocene horses (e.g., Azzaroli, 1998; Winans, 1985, 1989), a unique proposal is not yet available.
Both PCA and DA assays indicated the presence of three significantly different groups of horses. From the comparative analysis among the Cedral remains with the data of other horse fossil remains from Mexican localities, those three groups were assigned to three different horse species. One group corresponded to remains large in size from Cedral, associated with data of E. mexicanus. We suggested to assign it to E. mexicanus until the taxonomic situation of the Equus species of North America are correctly described and the specimens identified. This form was already reported as E. mexicanus (Alberdi et al., 2003) and was also found in other late Pleistocene Mexican localities as Valsequillo. A second, rarer group, corresponded to remains middle in size from Cedral associated with other sized specimens as E. conversidens (data of E. Scott transfer to M. Mora in 2006), and from other late Pleistocene Mexican localities. The previous assignation appeared correct (Alberdi et al., 2003).
Finally a third group, small in size, from Cedral and compared with the small sized remains from several Mexican localities. The appendicular skeleton, and specially the distal portions, are clearly smaller than for the other two species. This comparison by PCA and DA of the metapodials remains indicates that Cedral remains are smaller and less slender than other remains studied by Winans (1985, 1989): E. francisci and E. alaskae groups. Dental remains indicate an animal smaller than most of North American small horses known in the literature, the body mass is much reduced (Alberdi et al., in litteris), with the exception of E. tau described by Mooser and Dalquest (1975) and Dalquest (1977, 1979) that is smaller and slender (Table 4). Consequently, we assign a new name to Cedral remains smaller in size than E. alaskae and E. francisci, and less slender than both (metapodials being larger and slender in francisci than in alaskae). Previously, we assigned it to Equus sp. A (Alberdi et al., 2003), and here we name it as Equus cedralensis sp.nov.
As previously indicated (Alberdi et al., 2003), 14C radiocarbon assays for Cedral locality indicates a late Pleistocene age for this site (Lorenzo and Mirambell, 1986), ranging between 37,694 and 21,468 years BP. The morphological analysis indicates a high similarity, both in the teeth as in skeletal remains, among these three groups of horses, but the body mass and size are very variable among these three groups.
Also, the morphological differences affecting horses are mostly found in the distal limb bones, being related to environmental conditions, sediment and vegetation types. Pérez-Crespo et al. (2009) conclude, based on stable isotopes analysis from a Cedral's sample, that E. mexicanus and the Columbian Mammuthus columbi (Falconer, 1857), had a mixed diet, while E. conversidens and another unknown species of Equus had a more restricted C4 diet.
As for the horse remains from other Mexican late Pleistocene localities included in the present study, the applied names are well related to the recognized species at the Cedral locality. At Valsequillo, three horse species are assigned to Equus mexicanus, Equus conversidens, and Equus cedralensis. The scarce remains from Arroyo Cedazo (Aguascalientes, Mexico), housed at the Geology Institute at UNAM, confirmed that they pertained to a single species, E. conversidens; some large and small sizes were also present, the medium-sized is the dominant form, while at Cedral this form is more scarce. Finally, as for the important deposits at Tequixquiac, remains of at least the same three species occur, without bias to any one species.
CONCLUSION
We propose the presence of three distinct horse species in the region near Cedral, San Luis Potosí, Mexico. Those species show morphological characters quite similar among them, mostly on teeth morphology with a remarkable size difference in measures and body mass.
The large and robust species assigned to Equus mexicanus, a medium-sized animal associated to the name E. conversidens, and a small-sized but robust species to which the name E.cedralensis sp. nov. proposed to honor the town where Cedral is.
During the study of the large series of Mexican specimens, morphological similarity among the entire American horses from the late Pleistocene, mostly in regard to dental morphology was detected. Also, all of the horses were found at Rancholabrean deposits could be assigned to a time range between 50,000 to 10,500 years BP, the latest surviving period for the horses all over the Americas.
ACKNOWLEDGMENTS
We thank Dr. María del Carmen Perrillat for allowing us study specimens under her care at the Colección de Paleontología, Insitituto de Geología, Universidad Nacional Autónoma de México. Also, Antropologist Victor Hugo Valencia allowed studying those materials from Valsequillo housed at Centro INAH Puebla and Luis Via and Sebastian Calzada for the specimens from Tequixquiac stored at Museo Geológico del Seminario de Barcelona. We are grateful to Margarito Mora and Eric Scott for providing data on specimens housed in United States of America collections. Alejandro López-Jiménez provided technical assistance for preparation of photograph illustrations and Maria Teresa Olivera compose stratigraphy figure. The comments of Vera Eisenmann and Darrin Pagnac on a previous version of this article are very insightful to improve the text. We also thank Marisol Montellano and two anonymous reviewers and also the editor, their recommendations to improve this manuscript. Dr. Eileen Johnson kindly reviewed the modified English text for the language issues. This study was possible through the support of the Proyectos de Investigación conjunta con Iberoamérica (Bilateral Agreements CONACYT-CSIC 2001-2002, and 2007-2008) and the projects PB97-1250, BTE2001-1684, CGL2004-00400/BTE, CGL2007-60790/BTE and CGL2010-19116/BOS of the DGICYT, Spain.
SUPPLEMENTARY MATERIAL
Appendices 1 and 2 can be found at the journal website <http://rmcg.unam.mx/>, in the table of contents of this issue.
Appendix 1. Measurements of the isolate teeth from the Cedral site: P2/p2, P3-4/p3-4, M1-2/m1-2, and M3/m3.
Appendix 2. Measurements of the different bones from Cedral site. The measurements follow the recommendations of the "Hipparion Conference" New York 1981 (Eisenmann et al., 1988).
REFERENCES
Alberdi, M.T., Arroyo-Cabrales, J., Polaco, O.J., 2003, ¿Cuántas especies de caballo hubo en una sola localidad del Pleistoceno Mexicano?: Revista Española de Paleontología, 18, 205-212. [ Links ]
Alberdi, M.T., Prado, J.L., Ortiz-Jaureguizar, E., 1995, Patterns of body size changes in fossil and living Equini (Perissodactyla): Biological Journal of the Linnean Society, 54, 349-370. [ Links ]
Álvarez, T., Ocaña, A., 1987, Nomenclatura específica de los Equidae del Pleistoceno de México: Anales de la Escuela Nacional de Ciencias Biológicas, México, 31, 125-132. [ Links ]
Álvarez, T., Polaco, O.J., 1981, Anexo 1. Fauna obtenida de las excavaciones realizadas en el sitio Rancho La Amapola - Cedral, S.L.P., in Lorenzo, J.L., Mirambell, L. (eds.), Cedral, S.L.P., México: Un sitio con presencia humana de más de 30,000 AP: X Congreso de la Unión Internacional de Ciencias Prehistóricas y Protohistóricas, México, 123-124. [ Links ]
Álvarez, T., Polaco, O.J., 1982, Restos pleistocénicos de dos especies de Microtus (Rodentia: Muridae), del norte de San Luis Potosí, México: Anales de la Escuela Nacional de Ciencias Biológicas, México, 26, 47-53. [ Links ]
Álvarez, T., Ocaña, A., Arroyo-Cabrales, J., 2012, Restos de mamíferos, in Mirambell, L. (Coord.), Rancho "La Amapola", Cedral, un sitio Arqueológico-Palentológico, Pleistocénico-Holocénico con restos de actividad humana: México: Instituto Nacional de Antropología e Historia, 147-194. [ Links ]
Arroyo-Cabrales, J., 1992, Sinopsis de los Murciélagos fósiles de México: Revista de la Sociedad Mexicana Paleontológica, 5, 1-14. [ Links ]
Arroyo-Cabrales, J., Polaco, O.J., Álvarez, T., Johnson, E., 1996, New records of fossil tapir from northeastern México: Current Research in the Pleistocene, 13, 93-95. [ Links ]
Arroyo-Cabrales, J., Polaco, O.J., Johnson, E., Guzmán, A.F., 2003, The distribution of the genus Mammuthus in Mexico, in Reumer, J.W.F., De Vos, J., Mol, D. (eds.), Advances in mammoth research: Rotterdam, May 16-20 1999, Proceedings of the Second International Mammoth Conference, , Deinsea, 9, 27-39. [ Links ]
Azzaroli, A., 1995, A Synopsis of the Quaternary species of Equus in North America: Bollettino della Società Paleontologica Italiana, 34, 205-221. [ Links ]
Azzaroli, A., 1998, The genus Equus in North America - The Pleistocene species: Palaeontographia Italica, 85, 1-60. [ Links ]
Bennett, D.K., 1980, Stripes do not a Zebra make, Part I: A cladistic analysis of Equus: Systematic Zoology, 29, 272-287. [ Links ]
Bernor, R.L., Tobien, H., Woodburne, M.O.,1990, Patterns of Old World Hipparionine evolutionary diversification and biogeographic extension, in Lindsay, E.H., Fahlbusch, V., Mein, P. (eds.), European Neogene Mammal Chronology: Plenum Press, New York, USA, 263-319. [ Links ]
Carranza-Castañeda, O., Miller, W.E., 1987, Rediscovered type specimens and other important published Pleistocene mammalian fossils from Central Mexico: Journal of Vertebrate Paleontology, 7, 335-341. [ Links ]
Dalquest, W.W., 1977, Equus tau Owen from the Pleistocene of Mitchell County, Texas: The Texas Journal of Science, 29(1-2), 141. [ Links ]
Dalquest, W.W., 1979, The Little Horses (Genus Equus) of the Pleistocene of North America: The American Modland Naturalist, 101(1), 241-244. [ Links ]
Dalquest, W.W., Schultz, G.E., 1992, Ice Age Mammals of Northwestern Texas: Midwestern State University Press, Wichita Falls, Texas, 194-202, 203-219. [ Links ]
De Queiroz, K., 2007, Species concepts and species delimitation: Systematic Biology, 56, 879-886. [ Links ]
Eisenmann, V., Alberdi, M.T., De Giuli,C., Staesche, U., 1988, Collected papers after the "New York International Hipparion Conference, 1981", in Woodburne, M., Sondaar, P. (eds.), Studying fossil horses, 1, Methodology: E. J. Brill, Leiden, 1-72. [ Links ]
Falconer, H., 1857, On the species of mastodon and elephant occurring in the fossil state in England: The Quarterly Journal of the Geological Society of London, 14, 81-84. [ Links ]
Gidley, J.W., 1900, A new species of Pleistocene Horse from the Staked Plains of Texas: Bulletin of the American Museum of Natural History, 13, 111-116. [ Links ]
Gidley, J.W., 1901, Tooth characters and revision of the North American species of the genus Equus: Bulletin of the American Museum of Natural History, 14, 91-141. [ Links ]
Gray, J.E., 1821, On the natural arrangement of vertebrose animals: London Medical Repository Review, 15, 296-310. [ Links ]
Guthrie, R.D., 2003, Rapid body size decline in Alaskan Pleistocene horses before extinction: Nature, 426, 169-171. [ Links ]
Hay, O.P., 1913, Notes on Some Fossil Horses, with Descriptions of Four New Species: Proceedings of the United States National Museum, 44, 569-594. [ Links ]
Hay, O.P., 1915, Contribution to the knowledge of the mammals of the Pleistocene of North America: Proceedings of the United States National Museum, 48(2086), 515-575. [ Links ]
Hay, O.P., 1917, Description of a new species of extinct horse, Equus lambei, from the Pleistocene of Yukon Territory: Proceedings of the Unites States National Museum, 53, 435-443. [ Links ]
Hibbard, C.W., 1955, Pleistocene vertebrates from the upper Becerra (Becerra Superior) Formation, Valley of Tequixquiac, Mexico, with notes on others Pleistocene forms: Museum of Paleontology University of Michigan, Ann Arbor, 12, 47-96. [ Links ]
Hibbard, C.W., Taylor, D.W., 1960, Two late Pleistocene faunas from Southwestern Kansas: University of Michigan, Contributions from the Museum of Paleontology, 16, 1-223. [ Links ]
Hoffstetter, R., 1950, Algunas observaciones sobre los caballos fósiles de América del Sur Amerhippus gen.nov.: Boletín Infomaciones Científicas Nacionales, 3, 426-454. [ Links ]
Kurtén, B., Anderson, E., 1980, Mammalian Faunas of the Blancan and Pleistocene: Columbia University Press, 420 pp. [ Links ]
Leidy, J., 1858, Notice of remains of extinct Vertebrata, from the valley of The Niobrara River, collected during the exploring expedition of 1857, in Nebraska, under the command of Lieut. G.K. Warren, U. S. Top. Eng., by Dr. F.V. Hayden: Proceedings of the Academy of Natural Sciences of Philadelphia, 1858, 20-29. [ Links ]
Leidy, J., 1860, Description of vertebrate fossil. Holme's Post-Pliocene Fossils of South Carolina, 99-102. [ Links ]
Leidy, J., 1865, Bones and teeth of horses from California and Oregon: Proceedings of the Academy of Natural Sciences of Philadelphia, 17, 94. [ Links ]
Leidy, J., 1868, Notice of some remains of Horses: Proceedings of the Academy of Natural Sciences of Philadelphia, 20, 195. [ Links ]
Linnaeus, K., 1758, Systema naturae perregna tria naturae, secundum classes, ordines, genera, species cum characteribus, differentiis, synonymis, locis, Editio decima, reformata: Stockholm, Laurentii Salvii, 824 pp. [ Links ]
Lorenzo, J.L., Mirambell, L.,1986, Preliminary report on archaeological and paleoenvironmental studies in the area of Cedral, San Luis Potosí, México, 1977-1980, in Bryan, A.L. (ed.), New evidence for the Pleistocene peopling of the Americas: Center for the Study of the Early Man, University of Maine, Orono, Peopling of the Americas series,107-113. [ Links ]
Lorenzo, J.L., Mirambell, L., 1999, The inhabitants of México during the Upper Pleistocene, in Bonnichsen, R., Turnmire, K.L. (eds.), Ice age peoples of North America. Environments, origins, and adaptations of the First Americans: Center for the Study of the First Americans, Oregon State University Press, Corvallis, 482-496. [ Links ]
Lundelius, I.L.Jr., 1972, Fossil vertebrates from the late Pleistocene Ingleside Fauna, San Patricio County, Texas: Bureau of Economic Geology, University of Texas, Reports of Investigation, 77, 1-74. [ Links ]
Lundelius, I.L.Jr., Stevens, M., 1970, "Equus francisi" Hay, A small Stilt-Legged Horse, Middle Pleistocene of Texas: Journal of Paleontology, 44, 148-153. [ Links ]
Marcus, L.F., 1990, Traditional morphometrics, in Rohlf, F.J., Bookstein, F.L. (eds.), Proceedings of the Michigan Morphometrics Workshop: University of Michigan, Museum of Zoology, Special Publication 2, 78-122. [ Links ]
MacFadden, B.J., 1992, Fossil Horses. Systematics, Paleobiology, and Evolution of the Family Equidae: Cambridge University Press, 369 pp. [ Links ]
MacFadden, B.J., Hulbert, R.C.Jr., 1988, Explosive speciation at the base of the adaptive radiation of Miocene grazing horses: Nature, 336, 466-468. [ Links ]
Melgarejo-Damián, M.P., Montellano-Ballesteros, M., 2008, Quantitative differentiation of Mexican Pleistocene Horses: Current Research in the Pleistocene, 25, 184-186. [ Links ]
Mirambell, L., 2012, Rancho "La Amapola", Cedral, un sitio Arqueológico-Palentológico, Pleistocénico-Holocénico con restos de actividad humana: México: Instituto Nacional de Antropología e Historia, 320 pp. [ Links ]
Mooser, O., 1958, La fauna "Cedazo" del Pleistoceno en Aguascalientes: Anales del Instituto de Biología Mexicano, 29, 409-452. [ Links ]
Mooser, O., Dalquest, W.W., 1975, Pleistocene mammals from Aguascalientes, Central Mexico: Journal of Mammalogy, 56(4), 781-820. [ Links ]
Nobis, G., 1973, Die Equiden reste aus dem Pleistozän des Valsequillo (Mexico): Proyecto México de la Fundación Alemana para la Investigación Científica, 6, 95-108. [ Links ]
Olivera-Carrasco, M. T., 2012, Moluscos continentalesde Cedral, un sitio del Pleistoceno final de México, in Mirambell, L. (ed.), Rancho "La Amapola", Cedral, un sitio Arqueológico-Palentológico, Pleistocénico-Holocénico con restos de actividad humana: México: Instituto Nacional de Antropología e Historia, 225-283. [ Links ]
Ord, G., 1815, North American Zoology, in Guthrie, W. (ed.), A new geographical, historical, and commercial grammar: Second edition. Philadelphia, Pennsylvania, Lippincott, 2:1600, 291-361. [ Links ]
Osborn, H.F., 1912, Craniometry of the Equidae: Memoirs of the American Museum of Natural History, 1(3), 57-100. [ Links ]
Osborn, H.F., 1918, Equidae of the Oligocene, Miocene, and Pliocene of North America; iconographic type revision: Memoirs of the American Museum of Natural History, 2, 1-330. [ Links ]
Owen, R., 1848, Description of the teeth and portions of jaws of two extinct anthracotheroid quadrupeds discovered by the Marchioness of Hastings in the Eocene deposits of the N.W. coast of the Isle of Wight: with an attempt to develop Cuvier's idea of the classification of Pachyderms by the number of their toes: Quarterly Journal Geological Society of London, 4, 103-141. [ Links ]
Owen, R.,1869, On fossil remains of Equines from Central and South America referable to Equus conversidens Ow., E. tau Ow., and E. arcidens Ow.: Philosophical Transactions of the Royal Society of London, 159, 559-573. [ Links ]
Pérez-Crespo, V.A., Sánchez-Chillón, B., Arroyo-Cabrales, J., Alberdi, M.T., Polaco, O.J., Santos-Moreno, A., Benammi, M., Morales-Fuente, P., Cienfuegos-Alvarado, E., 2009, La dieta y el hábitat del mamut y los caballos del Pleistoceno tardío de Cedral con base en isótopos estables (δ13C, δ18O): Revista Mexicana de Ciencias Geológicas, 26(2), 347-355. [ Links ]
Pichardo, M., 2000, Valsequillo Biostratigraphy III: Equid Ecospecies in Paleoindian Sites: Anthropologisches Anzeiger, 58, 275-298. [ Links ]
Pichardo, M., 2004, Review of Horses in Paleoindian Sites of the Americas: Anthropologisches Anzeiger, 62, 11-35. [ Links ]
Pichardo, M., 2006, Update of Equids in paleoindean time, in Mashkour, M. (ed.), Equids in Time and Space: Papers in honour of Véra Eisenmann: Durham 2002, Proceedings of the 9th ICAZ Conference, Oxbow Books, Oxford, 31-48. [ Links ]
Pichardo, M., 2008, Identification Guide for horses from North American Paleoindian Time: Copyright 2008 Mario Pichardo 27 14th St., W Minneapolis, MN 55403 USA, 41 pp. [ Links ]
Pilsbry, H.A., 1890a, Two new species of U. S. land shells: The Nautilus, 4(1), 3-4, 4(4), 39. [ Links ]
Pilsbry, H.A., 1891(1890b), Note on a southern pupa: Proceedings of the Academy of Natural Sciences of Philadelphia, 1890, 44-45. [ Links ]
Pilsbry, H.A., 1890c (1889), New and little-known american molluscs. No. 1: Proceedings of the Academy of Natural Sciences of Philadelphia, 1889, 81-89. [ Links ]
Quinn, J.H., 1955, Miocene Equidae of the Texas Gulf Coastal Plain: Bureau of Economic Geology, The University of Texas, 5516, 5-101. [ Links ]
Quinn, J.H., 1957, Pleistocene Equidae of Texas: Bureau of Economic Geology. University of Texas, 33, 5-51. [ Links ]
Reyment, R.A., 1991, Multidimensional paleobiology: Pergamon Press, 416 pp. [ Links ]
Reynoso-Rosales, V.H., Montellano-Ballesteros, M., 1994, Revisión de los équidos de la Fauna Cedazo del Pleistoceno de Aguascalientes, México: Revista Mexicana de Ciencias Geológicas, 11(1), 87-105. [ Links ]
Savage, D.E., 1951, Late Cenozoic Vertebrates, San Francisco Bay Region: Bulletin of the Department of Geological Sciences, 28, 215-314. [ Links ]
Schafer, T.S., Dalquest, W.W., 1991, Comparison of Dental Characters of Fossil Horses in two Pleistocene Local Faunas: The Texas Journal of Science, 43, 45-49. [ Links ]
Scott, E., Stafford, T.W.Jr., Graham, R.W., Martin, L.D., 2010, Morphology and Metrics, Isotopes and Dates: Determining the Validity of Equus laurentius Hay, 1913: Journal of Vertebrate Paleontology, 30, 1840-1847. [ Links ]
Simpson, G.G., 1951, Horses: The Story of the horse Family in the Modern World and through Sixty Million Years of History: Oxford University Press, New York, 247 pp. [ Links ]
Skinner, M.F., 1942, The fauna of Papago Springs Cave, Arizona, with a study of Stockoceros: Bulletin of the American Museum of Natural History, 80, 143-220. [ Links ]
Slaughter, B.H., 1966, The Moore Pit Local Fauna; Pleistocene of Texas: Journal of Paleontology, 40, 78-91. [ Links ]
Slaughter, B.H., Crook, W.W.Jr., Harris, R.K., Allen, D.C., Seifert, M., 1962, The Hill-Shuler Local Faunas of the Upper Trinity River, Dallas and Denton Counties, Texas: Bureau of Economic Geology, Report of Investigation, 48, 1-71. [ Links ]
Stock, C., 1953, El Caballo pleistoceno "Equus conversidens leoni" subs. nov. de la cueva de San Josecito, Aramberri, Nuevo León: Memorias del Congreso Científico Mexicano, 3, 170-171. [ Links ]
Troxell, E.L., 1915, The Vertebrate Fossils of Rock Creek, Texas: American Journal Science, 39(234), 613-638. [ Links ]
Winans, M.C., 1985, Revision of North American fossil species of the genus Equus (Mammalia: Perissodactyla: Equidae): University of Texas, Austin Ph.D. Dissertation, 264 pp. [ Links ]
Winans, M.C., 1989, A quantitative study of North American fossil species of the genus Equus, in Prothero, D.R., Schoch, R.M. (eds.), The Evolution of Perissodactyls: Oxford, USA, Oxford University Press, 263-297. [ Links ]

