










Servicios Personalizados
Revista
Articulo
 Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista mexicana de ciencias geológicas
versión On-line ISSN 2007-2902versión impresa ISSN 1026-8774
Rev. mex. cienc. geol vol.30 no.1 Ciudad de México abr. 2013
Clasificación del área foliar de las gimnospermas fósiles de la zona norte de Oaxaca, México
Foliar area classification of fossil gymnosperms from northern Oaxaca, Mexico
Erika Lourdes Ortiz-Martínez1*, María Patricia Velasco de León1, Isaías Salgado-Ugarte1 y Alicia Silva-Pineda2
1 Facultad de Estudios Superiores Zaragoza, Universidad Nacional Autónoma de México, Av. Guelatao 66, Col. Ejército de Oriente, Iztapalapa, 09230 México D.F. *elom111176@hotmail.com
2 Instituto de Geología, Universidad Nacional Autónoma de México, Circuito Exterior, Ciudad Universitaria, 04510 México D.F.
Manuscrito recibido: Septiembre 15, 2011
Manuscrito corregido recibido: Septiembre 28, 2012
Manuscrito aceptado: Octubre 2, 2012
RESUMEN
Se da a conocer la clasificación del área foliar de diferentes géneros de gimnospermas fósiles de siete localidades jurásicas de México, con el fin de que pueda ser utilizada en la inferencia de las condiciones ambientales que favorecieron el establecimiento y dominancia de este grupo durante Jurásico Inferior y Medio del terreno Mixteco. Después de obtener el área foliar de 186 ejemplares completos y analizar los datos obtenidos utilizando estimadores de densidad por Kernel se proponen siete categorías del área foliar. En la zona de estudio las hojas microfilas son las más abundantes (87.08%). Sin embargo, la categoría Microfila I (0.08241-1.3660 cm2; 58.03 %) es el tamaño dominante y es la única presente en todas las localidades. Por otro lado, el análisis litológico hecho en la zona de estudio reveló la presencia de estratos alternantes de areniscas de diferente tamaño de grano y capas de carbón de espesor variable en todas las localidades. Para inferir el paleoclima se obtuvo el área foliar de las especies actuales de cicadales que se distribuyen en México. La categoría Microfila II (1.3661-3.4835 cm2) incluye las hojas más pequeñas encontradas en esta muestra; los organismos que las presentan se distribuyen en zonas con climas secos a húmedos. Esta categoría de área foliar es la más cercana a lo encontrado en las hojas fósiles del terreno Mixteco, lo que permite proponer que en el Jurásico Inferior existió un clima más seco que en el Jurásico Medio, con localidades con estrés hídrico al menos durante ciertas épocas del año y áreas con mayor humedad y presencia de pantanos. Sin embargo, las diferencias entre las localidades en los porcentajes de hojas microfilas, en el espesor de los estratos de carbón y en la textura de las rocas sedimentarias indican que las condiciones ambientales durante el Jurásico no fueron homogéneas.
Palabras clave: Paleobotánica, clima, Jurásico, terreno Mixteco, México.
ABSTRACT
The foliar area ofdifferent genera of fossil gymnosperms from seven Jurassic localities in Mexico was classified with the purpose of being used in the inference of the environmental conditions that favored the establishment and domination of this group in the Early andMiddle Jurassic in the Mixteco terrane. After getting the foliar area of 186 complete specimens and analyzing the data obtained using Kernel density estimation (Stata), seven categories of the foliar area were obtained. In the study area, the microphyllleaves were the most plentiful (87.08 %), however theMicrophylaIcategory (0.08241-1.3660 cm2; 58.03 %) is the dominate size and is the only found in all localities. On the other hand, a lithological analysis carried out in the study area revealed alternating strata of sandstone with variable grain size, and coal layers of variable thickness in all the localities. In order to infer the paleoclimate, the foliar area of the current cycada species distributed in Mexico was obtained. The smallest leaves found in this sample are included in the Microphyla II category (1.3661-3.4835 cm2), and come from organisms distributed in zones with dry to humid climate, and scarce rainfall. This foliar area category of the current species is the closest to what is found in the fossil leaves of the Mixteco terrane, which let us propose that the climate in the Early Jurassic was drier than in the Middle Jurassic, with localities subjected to hydric stress at least during some seasons of the year, and more humid swamp areas. The difference between the localities in the percentage of microphyll leaves, the variable thickness of the coal strata and the texture of the sedimentary rocks indicate that the climatic conditions during the Jurassic in the Mixteco terrane were not homogeneous.
Key words: Paleobotany, climate, Jurassic, Mixteco terrane, Mexico.
INTRODUCCIÓN
La distribución de la vegetación en el pasado, al igual que hoy en día, estuvo controlada en gran medida por factores ambientales entre los que destaca el clima. Los factores naturales que afectan a este complejo sistema son la latitud, altitud, continentalidad, corrientes marinas y vientos. Modificaciones en estos parámetros han ocasionado cambios climáticos que se han manifestado en la Tierra a lo largo del tiempo geológico causando extinciones en masa, alteraciones en el hábitat de los organismos y promovido la evolución de los seres vivos. Debido a esto, la relación entre la fisonomía foliar de las plantas y el clima ha despertado el interés de los investigadores desde mediados del siglo XVII (Seward, 1917). El tamaño de la hoja es uno de los caracteres adaptativos al clima mejor estudiado, pues controla la temperatura en la superficie de la lámina. Hojas grandes o con amplias áreas foliares expuestas se localizan en ambientes de climas templados y húmedos, hojas de áreas foliares reducidas tienden a presentarse en ambientes secos o soleados (Givnish, 1979; 1984; Hickey, 1981). Esta última adaptación surge en respuesta a la necesidad de las plantas de disminuir la absorción de radiación solar ante su imposibilidad de deshacerse rápidamente de la energía calórica emitida durante los procesos catabólicos. Un aumento brusco de la temperatura de la hoja provocaría que el metabolismo celular se viera seriamente afectado, poniendo en riesgo la sobrevivencia de estas estructuras.
Se ha demostrado que plantas adaptadas a un clima en particular presentan características morfológicas similares o ciertas formas de crecimiento. Esto ha permitido que disciplinas como la paleobotánica y la paleoecología utilicen la arquitectura foliar para realizar inferencias paleoclimáticas (Wolfe, 1971; Hickey y Wolfe, 1975; Stewart, 2008). Sin embargo, las reconstrucciones paleoecológicas que han utilizado hojas fósiles para inferir el clima se han enfocado principalmente en las angiospermas, dejando de lado las gimnospermas e ignorando los grandes cambios climáticos que se presentaron en la Tierra cuando estas plantas se diversificaron.
Fue al final del siglo pasado que los paleobotánicos comienzan a interesarse en las gimnospermas como una fuente importante de información, para inferir el clima de períodos anteriores al Paleógeno (Hallam, 1984; Ziengler et al., 1993; Rees et al., 2000; 2004; Krassilov, 2003). Para ello han utilizado listados de géneros presentes en localidades de Asía, y asociado su presencia a parámetros ambientales, por ejemplo, registran la existencia de Gingkgofitas en zonas húmedas y de Cheirolepidaceae en ambientes áridos (Krassilov, 1969, 2003). A nivel mundial se han empleado bases de datos para proponer una distribución latitudinal de microconíferas o plantas de hojas aciculares y megaconíferas o gimnospermas de hojas escuamiformes; lo mismo ha sucedido con las bennettitales que han sido clasificadas en microcicadas y macrocicadas, aunque en este grupo la designación de tamaños no es clara ni consistente. Ziengler et al. (1993) proponen cinco clases de tamaño para las bennettitales del Mesozóico temprano, 46% de las cuales no se han reportado en México; Rees et al. (2000, 2004), en su propuesta de biomas, consideran 10 clases de tamaño, aunque no explican los criterios utilizados en la generación de las categorías. En México, la flora fósil mejor preservada y más abundante se localiza en rocas jurásicas del estado de Oaxaca. Sin embargo, la aplicación de cualquiera de estos criterios en estudios paleoecológicos sería evidentemente inadecuada. Debido a lo anterior el objetivo de este trabajo es generar una clasificación estadística del área foliar de las gimnospermas fósiles del terreno Mixteco, que pueda ser utilizada en la inferencia de las condiciones ambientales que favorecieron su establecimiento durante el Jurásico Inferior y Medio, inferencias que tendrán que ser apoyadas por la información sedimentológica recabada en los afloramientos fósiles.
ZONA DE ESTUDIO
El terreno Mixteco se ubica al sur de la República Mexicana, aproximadamente a 200 km al sureste de la Ciudad de México. Se localiza en la porción noroeste del estado de Oaxaca cerca de los límites de los estados de Puebla, Guerrero y Oaxaca (Ortega-Guerrero, 1989), abarcando una superficie aproximada de 40,000 km2, entre las coordenadas geográficas 16°35' a 18°00' N y 97° 5' a 97°45' O (Figura 1). En la zona aflora una cubierta sedimentaria amplia del Jurásico Inferior y Medio que incluye a las formaciones Rosario, Conglomerado Cualac, Otlaltepec, Tecomazúchil y Grupo Tecocoyunca, las cuales en conjunto contienen a los afloramientos fósiles más abundantes y mejor preservados de gimnospermas de esta edad en México.

Geología y estratigrafía
Jurásico Inferior
La secuencia estratigráfica depositada sobre el terreno Mixteco durante el Jurásico inicia con rocas de la Formación Rosario (Caballero-Miranda, 1994, Figura 2) que afloran únicamente en la parte central de la cuenca de Tlaxiaco en las coordenadas geográficas 17°36'-12.9''N y 97°51'-37.1''O. Cubre en discordancia al Complejo Acatlán y tiene un espesor de 80.4 m en la zona de estudio. Jiménez-Rentería (2004) la ha definido como una facie carbonosa, diferente a la Formación Conglomerado Prieto. En la Formación Rosario se intercalan lutitas y areniscas de color café rojizo y amarillento, con horizontes de carbón no mayores a 30 cm de espesor. Las impresiones de cicadales, bennettitales, pteridospermas y filicales se encuentran mal preservadas en areniscas de grano medio y con mayor detalle en lutitas negras. Wieland (1914-1916) llamó a estos estratos "capas con plantas" y señaló un espesor para esta formación de 137 m. Erben (1956) considero un espesor entre 100 y 120 m. La edad asignada a esta unidad estratigráfica es Jurásico Inferior, pues subyace al Conglomerado Prieto (Toarciano; Jiménez-Rentería, 2004) (Figura 2). Las características litológicas de la formación indican un ambiente de depósito de tipo fluvial con llanuras de inundación (Erben, 1956; Morán-Zenteno et al., 1993; Jiménez-Rentería, 2004).
Jurásico Medio
La Formación Conglomerado Cualac aflora al este y noreste del estado de Guerrero en las cercanías del municipio de Cualac. Cubre concordantemente a la Formación Rosario y está cubierta por rocas del Grupo Tecocoyunca (Figura 2). Los sedimentos de la Formación Conglomerado Cualac que afloran en las coordenadas 17°35'52.8" N y 98°44'35" O (localidad Tres Caminos) tienen un espesor de 256 m, y están compuestos de conglomerados cuarcíticos duros, de color gris, con guijarros de cuarzo lechoso blanco y en menor frecuencia de micaesquistos y gneises. Impresiones de plantas se localizan en la columna estratigráfica a partir del metro 219 y hasta el 230; en los estratos inferiores de areniscas de grano fino y lutitas predominan Cheirolepidaceas y filicales; mientras que en los estratos superiores, conformados por areniscas de grano fino dominan las bennettitales bien conservadas. El Conglomerado Cualac se ha interpretado como un depósito continental, en un ambiente subáereo (estratos con concreciones de hierro) con pequeñas zonas pantanosas (presencia de intercalaciones de estratos de carbón que varían de 30 a 40 cm de espesor) (Erben, 1956; Velasco-de León et al, 2011; Velasco-de León y Guerrero-Arévalo, 2011).
La formación Otlaltepec (Figura 2) es una secuencia clástica (no descrita formalmente) que descansa discordantemente sobre la Unidad Piedra Hueca y en algunas ocasiones sobre el Complejo Acatlán y el Tronco de Totoltepec (Figura 2). Aflora en el área de Coyotepec-Tianguistengo, al suroeste del Estado de Puebla en las coordenadas geográficas 18°18'1.19'' N y 97°46'1.57''O (Ortega-Guerrero, 1989), tiene un espesor de 826.5 m y por sus características litológicas particulares se dividió a esta unidad en dos miembros: un miembro inferior con un espesor de 197m y un miembro superior que abarca aproximadamente 629 m (Cruz-Cruz, 2012). Las impresiones y permineralizaciones de plantas son escasas y poco diversas (troncos, ramas, frondas de bennetitales y filicales) y están preservadas en areniscas finas y lodolitas que se localizan en los últimos 40 metros del miembro inferior (Grajeda et al., 2011). Este miembro presenta estratos de carbón y estructuras secundarias como rizaduras, estratificación cruzada, lentes conglomeráticos y marcas de carga. Las características litológicas observadas, indican un ambiente fluvial de alta energía con períodos de máxima precipitación pluvial, lo que originó llanuras de inundación durante el relleno de la cuenca (Cruz-Cruz, 2012).
Cerca del Municipio de Huajuapan de León se han localizado rocas sedimentarias del Jurásico Medio pertenecientes a la Formación Tecomazúchil. Esta secuencia clástica tiene un espesor total de 705m. Se divide en 23 miembros (Grimaldo-Ávalos, 2010). El quinto de ellos, con un espesor aproximado de 9 m, contiene abundantes fósiles, entre los que destacan frutos, conos y frondas de Cycadophytas, así como una gran diversidad de filicales (Rojas-Chávez, 2010). Los miembros nueve y diez presentan bennettitales y Cheirolepidaceae. Esta unidad geológica se caracteriza por presentar litarenita arcósica y arcosa lítica con cantidades variables de limos y gravas de colores amarillentos y rojizos, con estratificación cruzada y laminación alterna poco definida. En las partes bajas de la formación se localiza una secuencia terrígena arrítmica de colores beige a gris claro, donde predominan las capas de arenisca y areniscas conglomeráticas con estratos que varían de 20 a 60 cm. Los rasgos mostrados por esta formación sugieren ser depósitos fluviales con desarrollo de barreras laterales y longitudinales combinadas con el desarrollo de sistemas meándricos, que se intercalan con abanicos aluviales y llanuras de inundación dentro de un graben, acumulados en un clima húmedo y cálido. Su contacto inferior con el Complejo Acatlán (Paleozoico) es discordante (Grimaldo-Ávalos, 2010; Arellano-Gil et al., 2011). Sobreyaciendo a la Formación Tecomazúchil se encuentran dos conjuntos de rocas de origen marino del Jurásico Superior: el primero corresponde a la Formación Chimeco y el segundo a la Formación Mapache (Figura 2), con 600 m de espesor máximo entre ambas formaciones (área de Texcalapa, Puebla), y que se acuña hacia Santiago Chilixtlahuaca, (Grimaldo-Ávalos, 2010; Arellano-Gil et al., 2011).
En la localidad Cañada del Ajo, que se encuentra a las afueras del poblado de Mixquiapan, Puebla, en las coordenadas 18°4'54" N, 98°16'16.8" O y 18°4'58.84" N, 98°16'25.68" O, afloran rocas del Grupo Tecocoyunca. En la localidad estudiada se midió una columna estratigráfica con un espesor total de 224.43 m. La parte basal de esta unidad corresponde a un conglomerado polimíctico con matriz arenosa intercalado con capas gruesas de conglomerado con clastos de cuarzo blanco y esquisto. La parte media está constituida por una secuencia terrígena arrítmica de colores beige a gris claro de capas de areniscas y areniscas conglomeráticas, con cuerpos lenticulares de conglomerados arenoso, estructuras de flute cast y bioturbación. La cima consiste en una intercalación de arenisca fina con conglomerado grueso a medio, con horizontes de lutita carbonosa y carbón, seguido por una alternancia de limolitas verdes y rojas con arenisca conglomerática, con algunas costras de hierro y nódulos de pedernal (Hernández-Vulpes y Rodríguez-Calderón, 2012). Los fósiles se encuentran en dos niveles, el primero, hacia la base de la columna, con bennettitales, filicales, equisetales, conos y coniferales, y el segundo, hacia la cima, con bennettitales y Cheirolepidaceas (Velasco-de León y Guerrero-Arévalo, 2011).
Este sistema presentó periodos de alta energía con la correspondiente acumulación de sedimentos, lo que originó la migración del cauce del río, teniendo como resultado un conjunto de canales abandonados, donde se acumuló materia orgánica vegetal, que dio lugar al carbón.
La última localidad del Grupo Tecocoyunca estudiada es la que aflora cerca de la ciudad de Tlaxiaco, ubicada en la localidad designada con el nombre de Río Ñumi que se ubica en las coordenadas geográficas 17°19'24.16" N y 97°43'3.21" O. Esta localidad forma parte de la Formación Zorrillo-Taberna indiferenciadas. Con este nombre, Carrasco-Ramírez (1981) conjunta las formaciones Zorrillo y Taberna, ya que presentan un notable parecido en litologías, y describe su relación transicional que hace difícil su reconocimiento fuera de las localidades tipo. Corro-Ortiz y Ruiz-González (2010) concuerdan con esta definición. Erben (1956) le asigna una edad Bajociana-Batoniana. La columna estratigráfica en la zona de estudio fue medida y arrojó un espesor de 303 m. Está constituida por areniscas intercaladas con lutitas que presentan una variación en el contenido de matriz, presentando litarenitas y grauvacas. Los líticos están constituidos por gneises y esquistos. El tamaño de los clastos, en su mayoría son subangulosos, varía entre arenas finas y gruesas. Conforman estratos tabulares e irregulares con espesores que van de los 30 cm a los 3 m. Las lutitas se encuentran en estratos delgados que están entre los 10 y 30 cm, aunque en algunos casos alcanzan espesores de hasta 1.5 m, y algunos contienen carbón, el cual aumenta hacia la parte superior, que es donde se localizan abundantes impresiones de corteza, frondas de bennetita-les, cicadales y filicales. La parte basal de la Formación Zorrillo-Taberna indiferenciadas fue depositada en un ambiente fluvial meándrico, mientras que la parte superior se depositó en un ambiente de llanura deltaica en su parte proximal, con presencia de zonas pantanosas ocasionales, que dieron origen a los depósitos de carbón.
MÉTODO
El trabajo paleontológico en el terreno Mixteco fue propuesto por la Facultad de Estudios Superiores Zaragoza (FES Zaragoza) y el Instituto de Geología de la Universidad Nacional Autónoma de México (UNAM) en el año 2010. El presente estudio se realizó con 186 hojas completas colectadas hasta ese momento en las diferentes localidades que conforman la cubierta sedimentaria del Jurásico de Oaxaca: Rosario Nuevo (Formación. Rosario); Tres Caminos (Formación Conglomerado Cualac); Tianguistengo (formación Otlaltepec); Ayuquila, Chilixtlahuaca, Partideño (Formación Tecomazúchil); Cañada del Ajo y Río Ñumi (Grupo Tecocoyunca).
Los fósiles de plantas fueron depositados en la colección paleontológica de la FES Zaragoza, UNAM, bajo los acrónimos CFZ R, CFZCC, CFZTe, CFZTz y CFZOt. De cada formación se limpió y midió un promedio de 31 ejemplares de gimnospermas (pinas en el caso de cicadales y bennettitales y hojas de coníferas). Para calcular el área foliar de cada ejemplar, los datos de largo y ancho fueron multiplicados por un factor de corrección de 0.66 (Welles y Norman, 1991; Ollat et al., 1998). A diferencia de la longitud, el área foliar controla la intensidad del intercambio gaseoso así como la actividad fotosintética de la planta (Welles y Norman, 1991), por lo que es un indicador de la radiación interceptada, de la cual depende el potencial fotosintético y productivo de las plantas (Ollat et al., 1998). Por esta razón, el área foliar refleja mejor las condiciones bajo las cuales vivieron los organismos.
Debido al pronunciado sesgo positivo de los valores de área foliar, los datos fueron transformados a logaritmos naturales, que facilitaron el manejo de la información evitando el sesgo de los datos. El análisis estadístico de la distribución del área foliar se realizó con ayuda del programa estadístico Stata 12, utilizando estimadores de densidad por Kernel (Salgado-Ugarte et al., 1997, 2005). La distribución multimodal resultante permitió el establecimiento de intervalos de tamaño que se utilizaron como indicadores de las categorías de área foliar de la muestra trabajada. Los límites de cada intervalo se basaron en la estimación de máximos (modas) y mínimos (antimodas) de frecuencia (Sanvicente-Añorve et al., 2003; Salgado-Ugarte et al., 1997, 2002, 2005), transformando posteriormente los valores logarítmicos a su escala original (cm2).
Por último se revisaron y midieron 10 pinas de diferentes frondas de 43 especies actuales de cicadales (22 de ellas corresponden al género Ceratozamia, 12 a Dioon y 9 a Zamia) depositadas en el Herbario Nacional (UNAM) y en el Herbario del Instituto de Ecología de Xalapa, Veracruz (XAL). Con los datos obtenidos se obtuvo el tamaño promedio para cada especie (largo y ancho de las pinas), para posteriormente calcular, con el mismo método utilizado para los organismos fósiles, el área foliar de cada especie actual de cicadales. Por último se ubicó el área foliar de estos ejemplares y de los 186 fósiles de gimnospermas del terreno Mixteco en las categorías generadas, con el fin de observar la representatividad de tamaños en la muestra fósil y comparar estos datos con el tamaño y las condiciones ambientales en las que viven las especies actuales de cicadales con áreas foliares cercanas a las encontradas en las hojas fósiles del terreno Mixteco. Se espera que esta información facilite, con ayuda de las características litológicas, la inferencia de las condiciones bajo las cuales vivieron las plantas de la mixteca oaxaqueña durante el Jurásico.
RESULTADOS
El análisis de densidad (Figura 3) muestra la generación de seis modas en donde cada una de ellas corresponde al punto medio de una categoría. El valor óptimo de Silverman fue de 0.4537, mientras que el valor sobresuavizado fue de 0.5767 (Tabla 1). La diferencia mínima entre estos valores evidencia la falta de sesgo al manejar el logaritmo natural de los valores originales. El ancho de la ventana se fijó en 0.26, según el criterio empírico que afirma que la mejor forma de fijar este valor es obteniendo la media del valor óptimo (Salgado-Ugarte, comunicación personal).


El análisis de antimodas permitió conocer los límites de cada una de las categorías (Tabla 2, Figura 4), encontrando que el área foliar más pequeña de las gimnospermas analizadas corresponde a valores menores a 0.01142 cm2, categoría representada por el género Brachyphyllum Brongiart, presente en las localidades de Ayuquila, Partideño, Tianguistengo, Tres Caminos y Río Ñumi, mientras que el grupo que presenta mayor superficie expuesta a la radiación es el que ostenta un área foliar mayor a 57.74 cm2. El 58.03 % del total de los ejemplares de la región norte del terreno Mixteco tiene una área foliar de 0.08241 a 1.3660 cm2 (bennettitales) coincidiendo con la categoría número tres (Microfila I). Las categorías menos representada en la muestra de hojas fósiles es la dos (Nanofila II) con apenas el 1.67% y la siete (Mesófila I) con el 0.53 %. El área foliar de estas categorías va de 0.01142 a 0.08240 cm2 (bennettitales) y mayores a 57.7428 cm2, respectivamente (Tabla 3).


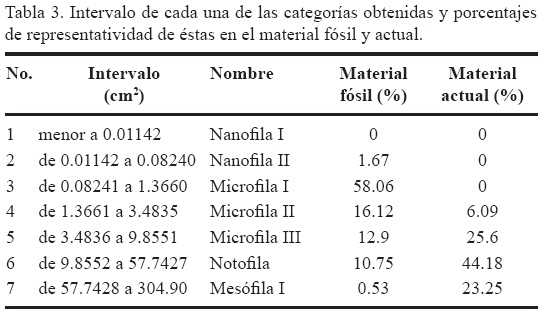
En comparación, las especies actuales de cícadas muestran hojas con mayor superficie foliar expuesta. La categoría Microfila II en esta muestra incluye las hojas de las especies más pequeñas y también las menos comunes (6.09%), siendo las especies Dioon angustifolia Miquel y Dioon caputoi De Luca, Sabato y Vázquez-Torres, las representantes de esta categoría. Actualmente sus miembros se distribuyen en climas secos, formando parte de matorrales xerófilos y selvas bajas caducifolias. Las hojas notofilas (44.18%) constituyen la clase mejor representada entre los organismos actuales, mientras que el género Ceratozamia Brongniart incluye a las hojas más grandes, con áreas foliares superiores a 57.7428 cm2. Estos organismos habitan en climas cálidos, principalmente como parte de la vegetación secundaria de selvas altas perennifolia y bosques mesófilos de montaña.
DISCUSIÓN
El análisis estadístico realizado con las impresiones de hojas fósiles colectadas en la zona norte del terreno Mixteco permitió clasificar las pinas en siete categorías de tamaño, nombradas en orden ascendente utilizando los prefijos de origen griego nano, micro, noto y meso (Tabla 3). En el área de estudio, las hojas microfilas son las más abundantes (87.08%). Sin embargo, sólo la categoría Microfila I está presente en todas las localidades y es el tamaño dominante, excepto en Partideño y Tianguistengo (Tabla 4). Esta clase incluye a las hojas microfilas más pequeñas con áreas foliares de 0.08 a 1.36 cm2. Esto indica que las bennettitales que dominaron el ambiente terrestre durante el Jurásico en el terreno Mixteco se desarrollaron en climas de secos a húmedos con escasas lluvia, marcado por periodos de estrés hídrico al menos durante ciertas épocas del año. Lo anterior se ve reforzado por la litología que muestra estratos alternos de areniscas de diferente tamaño de grano que fueron arrastrados por la corriente de los ríos durante la época de lluvias, y capas de carbón de espesor variable en todas las localidades. Este último tipo de roca se forma en depresiones con escasa circulación y abundante depósito de hojas, las cuales se ven sometidas a condiciones de alta temperatura y humedad, como en Río Ñumi, donde existe evidencia de miles de hojas acumuladas que han dado lugar a la formación de estratos de carbón con espesores de hasta 70 cm. Los ejemplares colectados en la Formación Otlaltepec y en la localidad del Partideño muestran un ligero aumento en el tamaño de las pinas (Microfila II y Microfila III, Tabla 4).
México cuenta con 60 especies actuales de cicadales distribuidas principalmente en climas cálidos. El área foliar de estas especies indica que las pinas de los organismos actuales son más grandes que en el Jurásico (Tabla 5). Por ejemplo, la categoría Microfila I, dominante en el material fósil, no está presente en las especies de cicadales actuales. Las pinas más pequeñas (Microfila II) dentro de las cícadas actuales se presentan en las especies Dioon angustifolia y Dioon caputoi. La primera se localiza únicamente en los estados de Nuevo León y Tamaulipas (Bosque de Quercus, Selva baja caducifolia, Selva baja perennifolia, pinar, Matorral alto espinoso y Bosque de pino), mientras que ejemplares de la especie Dioon caputoi se pueden ubicar en los límites de los estados de Puebla y Oaxaca, en las laderas de los cerros y matorrales xerófilos. Ambas especies sobreviven en ambientes en donde la temperatura media anual presenta un rango de 10 a 26 °C, mientras que la precipitación media anual va de 350 a 2000 mm, dependiendo de la comunidad.
CONCLUSIONES
La apertura del Golfo de México a principios de la era Mesozoica provocada por la separación de las placas tectónicas originó un rift en la región sur de México, el cual dio origen a una topografía accidentada en donde se depositaron los sedimentos de las formaciones que conforman la cubierta jurásica del terreno Mixteco. El análisis de estos sedimentos, lo observado en campo, el registro fósil y su análisis estadístico, permiten concluir que las unidades geológicas de la región norte del terreno Mixteco fueron depositadas en diferentes facies continentales como planicies de inundación, pantanos y lagos, en donde, de acuerdo con lo indicado por los tamaños de las pinas, se establecieron microclimas que permitieron el establecimiento de diferentes comunidades vegetales.
AGRADECIMIENTOS
La primera autora desea expresar su agradecimiento al Consejo Nacional de Ciencia y Tecnología (Conacyt) por el apoyo financiero recibido durante sus estudios de posgrado, becario CVU 164601, y los coautores por el financiamiento recibido al proyecto CONACYT, IN106010, y proyecto 103773 (PAPIIT, DGAPA, UNAM).
REFERENCIAS
Arellano-Gil, J., Velasco-de León, M.P., Grimaldo, J.R., Silva-Pineda, A., Yussim, S., Ortiz-Martínez E.L., 2011, Análisis estratigráfico de la secuencia continental del Jurásico Medio en región de Ayuquila Santiago Chilixtlahuaca, en los límites de los estados de Puebla y Oaxaca, en Congreso Nacional de Paleontología, Puebla, México, p. 19. [ Links ]
Caballero-Miranda, C., 1994, Fabrica magnética de secuencias continentales jurásicas de Oaxaca - Puebla, México: Facultad de Ingeniería Universidad Nacional Autónoma de México, tesis doctoral, 95 pp. [ Links ]
Carrasco-Ramírez, R., 1981. Geología del área de Tlaxiaco, Mixteca Alta, Oaxaca: Universidad Nacional Autónoma de México, Facultad de Ciencias, tesis de maestría, 105 pp. [ Links ]
Corro-Ortiz, G., Ruiz-González F., 2011, Análisis estratigráfico de las secuencias jurásicas de la región de Tlaxiaco, México: Universidad Nacional Autónoma de México, Facultad de Ingeniería, tesis de licenciatura, 75 pp. [ Links ]
Cruz-Cruz, M.A., 2012, Análisis estratigráfico de la secuencia Jurásica de la región de Santo Domingo Tianguistengo, Oaxaca, Santa Cruz Nuevo Puebla, México: Universidad Nacional Autónoma de México, Facultad de Ingeniería, tesis de licenciatura, 59 pp. [ Links ]
Erben, H.K., 1956, El Jurásico Medio y el Calloviano de México, en XX Congreso Geológico Internacional: Universidad Nacional Autónoma de México, Instituto de Geología, Monografía, 140 pp. [ Links ]
Givnish, T.J., 1979, On the adaptative significance of leaf from, en Solbrig, O.T., Jain, S., Johnson, G.B., Raven, P.H. (eds.), Tropics in plant population biology: New York, Columbia University Press, 375-407. [ Links ]
Givnish, T.J., 1984, Leaf and canopy adaptations in tropical forest, en Medina, E., Mooney, H.A., Vazquez-Yañez, C. (eds.), Phisiological Ecology of Plants of the Wet Tropics: La Haya, Dr. W. Junk, 51-84. [ Links ]
Grajeda, C.L., Velasco-de León, M.P., Cruz-Cruz, M.A., Arellano, J., 2011, Gimnospermas y ambiente de la formación Otlaltepec del Jurásico Medio, en los límites de Puebla y Oaxaca (resumen), en Sour-Tovar, F., Martin M. L., Reynoso R. V.H. (eds.), XII Congreso Nacional de Paleontología, Puebla de los Ángeles, México, p. 70. [ Links ]
Grimaldo-Ávalos, J.R., 2010, Análisis estratigráfico de las secuencias jurásicas de La región de Ayuquila-Santiago Chilixtlahuaca, Oaxaca. México: Universidad Nacional Autónoma de México, Facultad de Ingeniería, tesis de licenciatura, 103 pp. [ Links ]
Hallam, A., 1984, Continental humid and arid zones during the Jurassic and Cretaceous: Palaeogeograhy, Palaeoclimatology, Palaeoecology 47, 195-223. [ Links ]
Hernández-Vulpes, R.M., Rodríguez-Calderón, C.M., 2012, Análisis estratigráfico de la secuencia jurásica de la región de Tecocoyunca Tecomatlan, Puebla, México: Universidad Nacional Autónoma de México, Facultad de Ingeniería, tesis de licenciatura, 120 pp. [ Links ]
Hickey, L., 1981, Land plant evidence compatible with gradual, not catastrophic change at the end of the Cretaceous: Nature, 292, 529-531. [ Links ]
Hickey, L., Wolfe, J.A., 1975, The bases of angiosperm phylogeny vegetative morphology: Annals Missouri Botanical Garden, 62 pp. [ Links ]
Jiménez-Rentería, J., 2004, Estudio palinoestratigrafico de los grupos Consuelo y Tecocoyunca (Triásico-Jurásico) en la barranca Rosario Nuevo, Región de Tezoatlan al noroeste del estado de Oaxaca, México: Universidad Nacional Autónoma de México, Facultad de Ingeniería, tesis de licenciatura, 102 pp. [ Links ]
Krassilov, V.A., 1969, On reconstruction of extinct plants: Paleontology Ziurn, 1, 3-12 [ Links ]
Krassilov, V.A., 2003, Terrestrial Paleoecology and global change: Moscow, Pensoft, 463 pp. [ Links ]
Morán-Zenteno, D.J, Caballero-Miranda, C, I, Silva-Romo, G, Ortega-Guerrero, B, González-Torres, E, 1993, Jurassic-Cretaceous paleogeographic evolution of the northern Mixteca terrane southern México: Geofísica Internacional 32, 453-473. [ Links ]
Ollat, N., Fermaud, M., Tandonnet, J.P., Neveux, M., 1998, Evaluation of an indirect method for leaf area index determination in the vineyard: Combined effects of cultivar, yeard and training system: Vitis, 37(2), 73-78. [ Links ]
Ortega-Guerrero, B., 1989, Paleomagnetismo y geología de las unidades clásticas mesozoicas del área de Totoltepec-Ixcaquixtla, Estados de Puebla y Oaxaca, México: Universidad Nacional Autónoma de México, Facultad de Ciencias, tesis de maestría, 155 pp. [ Links ]
Ortega-Gutiérrez, F., Elías-Herrera, M., Reyes-Salas, M., Macías-Romo, C., López, R., 1999, Late Ordovician-Early Silurian continental collisional orogeny in Southern Mexico and its bearing on Gondwana-Laurentia connections: Geology, 27(8), 719-722. [ Links ]
Rees, P.M., Ziengler, A.M., Valdes, P., 2000, Jurassic Phytogeography and climates; New data and model comparisons, en Huber, B.T. Macleod, K.G. Wing, S.L. (eds), Warm Climates in Earth History: Cambridge University Press, 297-318. [ Links ]
Rees, P.M., Noto, C.R., Parris, J.M., 2004, Late Jurassic climates, vegetation and dinosaur distributions: The Journal of Geology, 112, 643-653. [ Links ]
Rojas-Chávez, C., 2010, Taxonomía de Dicksoniaceae, Gleicheniaceae y Matoniaceae (Filicales) del Jurásico en la región Mixteca: México, Universidad Nacional Autónoma de México, Facultad de Estudios Superiores Zaragoza, tesis de licenciatura, 165 pp. [ Links ]
Salgado-Ugarte, I.H., Shimizu, M., Taniuchi, T., 1997, Nonparametric assessment of multimodality for univariate data: Stata Technical Bulletin, 38, 27-35. [ Links ]
Salgado-Ugarte, I.H., Shimizu, M., Taniuchi, T., Matsushita, K., 2002, Nonparametric assessment of multimodality for size frequency distributions: Asian Fisheries Science, 15, 295-303 [ Links ]
Salgado-Ugarte, I.H., Gómez-Márquez, J.L., Peña-Mendoza, B., 2005, Métodos actualizados para análisis de datos biológicos-pesqueros: Universidad Nacional Autónoma de México, Facultad de Estudios Superiores Zaragoza, 240 pp. [ Links ]
Sanvicente-Añorve, L., Salgado-Ugarte, I.H., Castillo-Rivera, M., 2003, The use of kernel density estimators to analyze length-frequency distributions of fish larvae, en Browman, I.H., Skiftesvik A.B. (eds.), The Big Fish Bang: Proceeding of the 26th Annual Larval Fish Conference: Bergen, Noruega, Intitute of Marine Research, 419-430. [ Links ]
Seward, A.C., 1917, Fossil plants, v. 3, Pteridospermeae, Cycadofilices, Cordaitales, Cycadophyta: Cambridge University Press, v. 3, 656 pp. [ Links ]
Stewart, W.N., 2008, Paleobotany and the evolution of plants: London, Cambridge University Press, 521 pp. [ Links ]
Velasco-de León, M.P., Guerrero Arévalo, I.D., 2011, Análisis Tafonómico y sedimentológico de Barranca de la mina, Tecomatlán, Puebla, en Reunión anual de la Unión Geofísica Mexicana, Pto. Vallarta, Jalisco, México p. 23-24. [ Links ]
Velasco-de León, M.P., Domínguez, A., Mercado-Millán, H.A., Arcos-Hernández, J.L., 2011, Paleontología y estratigrafía de dos localidades en Guerrero de la Formación Cualac, en Congreso Nacional de Paleontología, Puebla, México, p. 32. [ Links ]
Welles, J.M., Norman J.M., 1991, Instrument for indirect measurement of canopy architecture: Agronomy Journal, 83, 818-825. [ Links ]
Wieland, G.R. 1914-1916, La flora liásica de la Mixteca Alta: México, Secretaría de Fomento, Instituto Geológico de México, Boletín 31, 165 pp. + 50 láminas. [ Links ]
Wolfe, J., 1971, Tertiary climatic fluctuations and methods of analysis of Tertiary floras: Palaeogeography, Palaeoclimatology, Palaeoecology 9, 27-57. [ Links ]
Wolfe, J., 1985, Distribution of major vegetational types during the Tertiary, en Sundquist E.T., Broecker, W.S., The carbon Cycle and Atmospheric CO2: Natural variations Archean to Present: American Geophysical Union, Monograph 32, 357-375. [ Links ]
Ziengler, A.M., Parris, J., Ping, E.D., Gyllenhaal, D.B., Rowlwy, J.T., Parrish, N., Shangyou, A., Beker, L., Hulver, M., 1993, Early Mesozoic Phytogeography and Climate. Philosophical Transactions of the Royal Society: Biological Sciences, 341, 297-305. [ Links ]

