










Serviços Personalizados
Journal
Artigo
 Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkRevista mexicana de ciencias geológicas
versão On-line ISSN 2007-2902versão impressa ISSN 1026-8774
Rev. mex. cienc. geol vol.29 no.3 Ciudad de México Dez. 2012
The first report of "chaetetids" from the Cretaceous of North America and their palaeoecological implications
Primer reporte de "chaetetidos" del Cretácico de Norte América y sus implicaciones paleoecológicas
Francisco Sánchez-Beristain1*, Pedro García-Barrera1, and José Ramón Torres-Hernández2
1 Museo de Paleontología, Facultad de Ciencias, Universidad Nacional Autónoma de México, Circuito Exterior s/n, Coyoacán 04510, México, D.F., México. * sanchez@ciencias.unam.mx.
2 Instituto de Geología, Universidad Autónoma de San Luis Potosí, Álvaro Obregón 68, 78000, Centro Histórico, San Luis Potosí, SLP, México.
Manuscript received: March 23, 2012
Corrected manuscript received: June 27, 2012
Manuscript accepted: June 29, 2012
ABSTRACT
The recently discovered Upper Cretaceous locality El Gorrión (SI48, San Luis Potosí, central Mexico) yielded three specimens of chaetetids. All specimens can be attributed to the genus Blastochaetetes Dietrich due to the discontinuities in the calicle walls. In addition, one of them could be classified as Blastochaetetes flabellum (Michelin) based on its distinctive internal dimensions. The remaining two specimens were assigned to an undetermined species taxon in open nomenclature, namely Blastochaetetes sp. indet. 1.
Three growth forms were identified for the chaetetids of El Gorrión. Blastochaetetes flabellum has a laminar growth form, which may be an adaptation to survive high-energy conditions, depending on whether the chaetetid was fixed to its substrate or not. The two remaining specimens show either a smooth, non-enveloping or a ragged, high-domical morphotypes, which respond to different environmental conditions.
The material is strongly silicified and therefore poorly preserved. The specimens lack spicules, so it is not possible to place them into any of the known sponge groups. Nevertheless, these fossils, as chaetetids, are clearly sponges.
Thisfinding is of great importance since such organisms have been neither reported for the Mesozoic in Mexico, nor for the Cretaceous in North America. Furthermore, it constitutes the second report of chaetetids for the Mesozoic of both North and Latin America.
Key words: chaetetids, Porifera, Blastochaetetes, Mexico, Cretaceous.
RESUMEN
Se encontraron tres especímenes de quetétidos en la recientemente descubierta localidad del Cretácico Superior "El Gorrión" (SI48, San Luis Potosí, centro de México). Todos ellos pueden ser asignados al género Blastochaetetes Dietrich con base en las discontinuidades en las paredes caliculares. Uno de ellos pudo ser clasificado como Blastochaetetes flabellum (Michelin) gracias a sus dimensiones internas distintivas. Los dos especímenes restantes fueron asignados a un taxón indeterminado a nivel específico en nomenclatura abierta: Blastochaetetes sp. indet. 1.
Tres formas de crecimiento fueron identificadas para los quetétidos de El Gorrión. Blastochaetetes flabellum posee una forma laminar, que puede ser considerada como una adaptación a condiciones de alta energía, dependiendo si el quetétido estuviese o no anclado al sustrato. Los dos especímenes restantes muestran dos morfotipos; uno liso no-envolvente y el otro dómico, alto y mellado. Ambos morfotipos responden a condiciones ambientales diferentes.
El material se encuentra en un pobre estado de conservación debido a la silicificación. Los especímenes carecen de espículas, por lo que no es posible asignarlos a algún grupo conocido de esponjas. No obstante, estos fósiles, como quetétidos, son claramente esponjas.
Este hallazgo es de gran importancia debido a que este tipo de organismos no han sido reportados para el Mesozoico de México o para el Cretácico de Norteamérica. Asimismo, representa el segundo reporte de quetétidos mesozoicos tanto para Norteamérica como para Latinoamérica.
Palabras clave: quetétidos, Porifera, Blastochaetetes, México, Cretácico.
INTRODUCTION
Chaetetids are marine organisms possessing a skeleton composed of tubules, now considered by most workers (e.g., Reitner, 1992; Flügel, 2004) to be sponges. They have been important reef builders in the geological past (Riedel, 1990). They present an external shape, which ranges from thin laminar to bulbous ones. The tubules of the calcareous skeleton are named calicles, which have shapes ranging from polygonal to irregular in cross section. This variability in shape can be attributed to environmental perturbations. Occasionally, calicles may display structures named pseu-dosepta within their walls. These septa may disappear as a consequence of calicle growth (West and Clark, 1983).
Their primary skeleton is often composed of siliceous spicules, whereas their basal, secondary calcareous skeleton is formed by abundant tubules, which are independent and narrow (Reitner, 1991, 1992; Cremer, 1994). The soft, living body of the sponge settles on top of the calcareous skeleton (Hartman and Goreau, 1972; Reitner, 1992). Modern representatives of the chaetetid sponges lose their spicules during their lifetime (Hartman and Goreau, 1970). This could be the reason why many fossil chaetetid sponges lack spicules, thus their classification as members of the Phylum Porifera has been problematic. However, numerous findings of spicules within the calcareous skeletons of some chaetetids have been previously reported (e.g., Kazmierczak,1979; Gray, 1980; Reitner, 1992; Cremer, 1994,). This fact supports their classification within the Porifera. Moreover, the taxonomic situation of some chaetetid organisms has been redefined by the recent interpretation of the type-species of the genus Solenopora (formerly an alga) as a sponge with a "chaetetid" bauplan (Riding, 2004). The term "chaetetid" is therefore taxonomically no longer valid and should only be used for morphological approaches, taking into account that in fact, all fossil chaetetids could indeed be sponges (Flügel, 2004).
The importance of this paper relies on the fact that no chaetetid sponges have been reported for the Mesozoic of Mexico. In addition, it represents the first Cretaceous report for North America and barely the second Mesozoic finding for both North America (Jansa et al., 1982), and Latin America (Wells, 1944). Furthermore, this work intends to contribute to the knowledge of the overall poor fossil record of chaetetids and their palaeoecology.
Finally, we want to remark that this work refers geographically to Mexico as a part of North America (Ángeles-Villarreal, 2010).
GEOLOGICAL SETTING
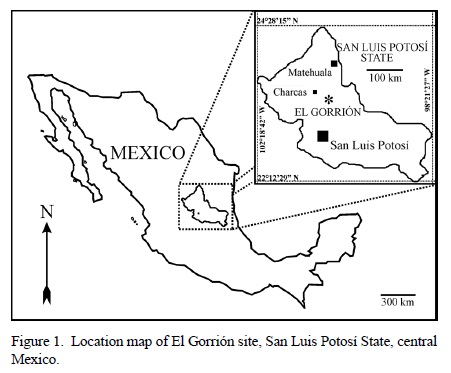
The fossil locality El Gorrión (San Luis Potosí State, central Mexico, Figure 1) is geologically part of the Valles-San Luis Potosí platform. Although this recently discovered locality (Sánchez-Beristain, 2005) yields a rich fossil assemblage (mainly remains of rudists and undetermined corals), this paper focuses on the remarkable presence of chaetetids.
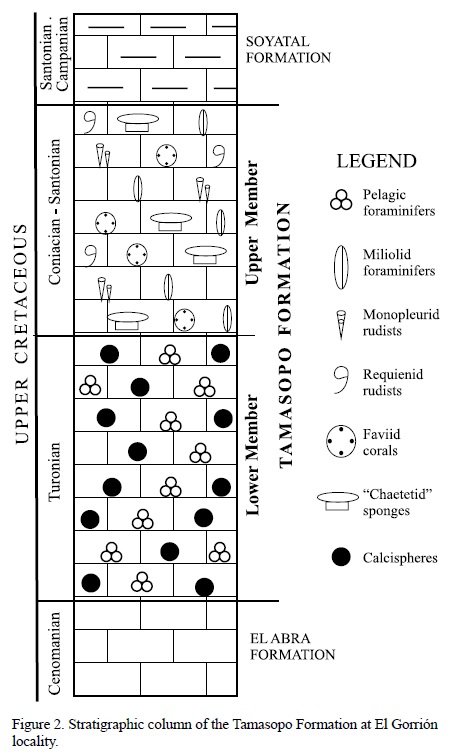
The Tamasopo Formation, which crops out at this locality, is comprised of a Lower and an Upper Member (Carrillo-Bravo, 1971; Basáñez-Loyola et al., 1993; Figure 2). The Lower Member of the unit is Turonian in age and consists of gray wackestone with calcispheres and pelagic foraminifers from lagoonal and open platform facies. It concordantly overlies the El Abra Formation deposits along with the Agua Nueva Formation. The Upper Member under- lies the Santonian-Campanian Soyatal Formation and also consists of gray limestone from lagoonal and open platform facies. It contains monopleurid and requienid rudists, faviid corals, miliolid foraminifers, as well as the studied chaetetid sponges. This stratigraphic unit was concordantly deposited over the Lower Member during the Coniacian and Santonian times, which can be verified by the presence of both Dicarinella sp. and Marginotruncata angusticarinata (Basáñez Loyola et al., 1993).
MATERIAL AND METHODS
Three specimens were collected at El Gorrión. All of them are housed in the Museo de Paleontología at the Facultad de Ciencias, Universidad Nacional Autónoma de México (UNAM), Mexico City (FCMP) with the inventory numbers FCMP SI 48-2004 (FCMP SI 48-2004-447/2, FCMP SI 48-2004-447/3 and FCMP SI 48-2004-447/5). Five thin sections with format 27 x 46 mm and ca. 70 µm thick were made from these specimens; two for both specimens FCMP SI 48-2004-447/2 and FCMP SI 48-2004447/5, and one for specimen FCMP SI 48-2004-447/3.
The description of the morphology of the secondary skeleton follows the terminology of Fischer (1970) and West and Clark (1983). The material was collected at the open platform facies of the middle level of the Upper Member of the Tamasopo Formation (Basáñez-Loyola et al., 1993). Nomenclature and internal morphological descriptions follow Cremer (1994, 1995), Fischer (1970) and West and Clark (1983). Growth form descriptions follow Kershaw and West (1991).
It should be taken into consideration that chaetetids are rarely easily extracted from the rocks that bear them, and therefore one faces the perspective of a two-dimensional organism rather than a three-dimensional one (Kershaw and West, 1991). This would obviously affect the identification of their growth form. However and fortunately, this is not the case for El Gorrión chaetetids, since they are easily extracted from their substrate.
SYSTEMATIC PALAEONTOLOGY
Phylum ?Porifera Grant, 1836
Class Demospongiae Sollas, 1885
Orden Axinellida Levi, 1955
Family ?Stromatoaxinellidae Wood and Reitner, 1988
Genus Blastochaetetes Dietrich, 1919
(Figures 3-12) (4, 5, 6, 7, 8, 9, 10, 11)






Blastochaetetes gen. nov. Dietrich 1919, p. 210, fig. 2
Blastochaetetes Fischer, 1970, p. 30
Stromatoaxinella gen. nov. Wood and Reitner, 1988, p. 215, figs. 4-7
?Blastochaetetes Cremer, 1994, p. 94-95, pl.1, fig.1
Type species. Chaetetes capilliformis Michelin, 1845.
Diagnosis. Chaetetids with discontinuous calicle walls. Calicle multiplication by intraparietal gemmation or by fissiparous division. Growth zones more or less clearly recognizable along the calicles.
Occurrence. Upper Triassic (Carnian) to Upper Cretaceous (Coniacian-Santonian).
Discussion. Due to the discontinuous walls, the three specimens in this work could be assigned to one of the following genera: Ptychochaetetes or Blastochaetetes. However, the former can only be found from the Upper Triassic to the Early Cretaceous (Fischer, 1970; Cremer, 1995), whereas the latter can be found from the Jurassic to the Upper Cretaceous (Fischer, 1970; Wood and Reitner, 1988). In addition, the possible relicts of a fibro-radial microstructure confirm this assessment, since Ptychochaetetes displays only a granular microstructure, whereas Blastochaetetes possesses always a fibro-radial (clinogonal) microstructure.
Wood and Reitner (1988) and Cremer (1994) mentioned a notorious problem related to the systematics of the latter genus. According to them, it should not be defined only by the presence of discontinuous calicles. Since spic-ule pseudomorphs were found in three specimens formerly conferred to the taxon Blastochaetetes irregularis, a new genus (Stromatoaxinella) was erected by Wood and Reitner (1988). Cremer (1994) proposed polyphyly for the species enclosed in this genus based on the finding of Wood and Reitner (1988) and urged to deepen its study.
In this work we classify the three specimens in the genus Blastochaetetes Dietrich based solely on their internal morphology an on their stratigraphical distribution, since no spicules/spicule pseudomorphs were found in any of them.
Blastochaetetes sp. indet. 1
Description. Specimen FCMP SI 48-2004-447/2 (in the following, 447/2).
External characters. The specimen consists of a secondary skeleton measuring 4.5 cm in length and 3.5 cm in width.
Height is 2.5 cm. (Figures 3, 4). It presents a smooth, non-enveloping, high-domical growth form. Calicle discontinuities can be observed on the upper surface. It is strongly recrystallized.
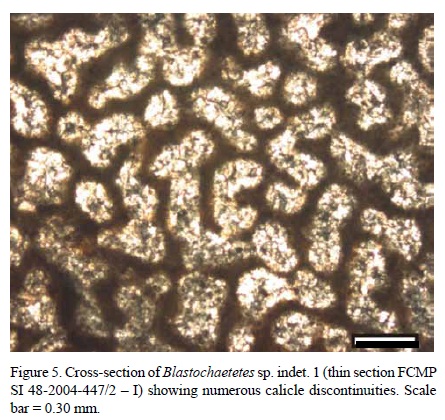
Internal morphological characters. Calicles are mostly round to elliptical. However, in most cases there is evidence of fissiparous division processes which do not allow the designation of a delimited tube shape (Figures 5, 6).
The mean value of tubes/mm 2 is 9, although this value can even reach 12. Although calicle centre to centre diameter measures 0.40 to 0.50 mm, most values approach the highest value.
Calicle diameter lies between 0.20 and 0.50 mm, although most of them measure around 0.3 mm. Double walls are 0.09 to 0.15 mm in thickness, a striking measure among chaetetids due to the high values (Cuif and Fischer, 1974; Fischer, 1970). Mean value is 0.12 mm.
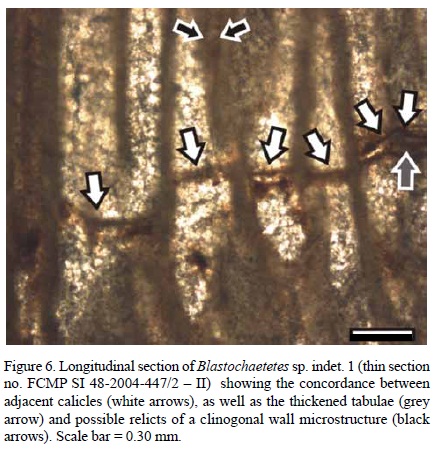
Tabulae thickness measures 0.06 to 0.13 mm, with a mean value of 0.1 mm. Tabulae are more or less concordant between adjacent calicles (Figure 6). However, they are difficult to find. Intertabular spaces are no less than 1.5 mm. Sánchez-Beristain (2005) misinterpreted thick tabulae as interrupted growth zones, which are actually absent (Figure 6).
Numerous discontinuities are observed along calicle walls, which could correspond to fissiparous division processes.
There is no clear evidence of the nature of the microstructure of the skeleton. However, it may have been fibro-radial (clinogonal), based on some poorly preserved angular structures (needles?, Figure 6).
Description. Specimen FCMP SI 48-2004-447/3 (in the following, 447/3)
External characters. This specimen also consists of a secondary skeleton. It displays a high-domical, partly ragged growth pattern, having thus a fungiform external appearance (Figures 7, 8). This form resembles the "ragged" growth form of James and Bourque (1992). The material is also strongly recrystallized. It measures 2.5 cm in lenght and 1.5 cm in width. Height is 2.5 cm (Figures 7, 8). Calicle discontinuities can also be observed on the upper surface.
Internal morphological characters. Few differences are observed in comparison with specimen 447/2. Calicles are mostly round to elliptical, with several division processes. The mean value of tubes/mm 2 is ca. 9, although this value can even reach 11 to 12. Calicle centre to centre diameter measures 0.40 to 0.50 mm, but most values approach the highest value, as for specimen 447/2. Calicle diameter lies between 0.20 and 0.50 mm, although most of them measure around 0.35 mm.
Measurements of double walls, tabulae thickness and intertabular spaces render the same values as for specimen 447/2. Tabulae are equally more or less concordant between adjacent calicles.
In this specimen there is neither clear evidence of the nature of the microstructure of the skeleton.
Examined material. FCMP SI 48-2004-447/2 and two thin sections obtained from it, labeled: FCMP SI 48-2004-447-2 - I and FCMP SI 48-2004-447-2 - II. FCMP SI 48-2004447/3 and two thin section obtained from it, labeled: FCMP SI 48-2004- 447-3 - I and FCMP SI 48-2004-447-3 - II
Occurrence. El Gorrión Limestone, part of the middle Upper Member of the Tamasopo Formation of Coniacian-Santonian age, San Luis Potosí state, Mexico.
Discussion. Due to the scarcity of specimens and to the lack of sufficient diagnostic characters to erect a new species epithet, we decided to let the taxonomical classification for this species with an open nomenclature. However, some affinities exist for certain characters between these specimens and some other species of the genus Blastochaetetes, which will be discussed next, as well as the impossibility to name a new species based solely on these affinities.
Upper Cretaceous species of the genus Blastochaetetes differ from these two specimens in particular with regard to some of the internal morphological dimensions and external shape. Three species of the genus plus one rede-scribed taxon, Stromatoaxinella irregularis, are used here for comparison. This redescribed taxon has been classified undoubtedly as an axinellid sponge due to the presence of spicule pseudomorphs (Wood and Reitner 1988). In this paper, this name is considered valid, taking into account, however, that the taxon bearing it previously belonged to the genus Blastochaetetes (Fischer, 1970).
Blastochaetetes flabellum [(Michelin), Senonian of d'Alais, France], Blastochaetetes petri [(Zuffardi-Commerci), Turonian of Presso Teghirinna, Italy], Stromatoaxinella irregularis [(Michelin), Lower "Senonian" of Martigues, France] and Blastochaetetes coquandi [(Michelin), Senonian of Mazaugues; France] do not differ much between each other in calicle density/ mm2 and calicle diameter (Table 1). These characters also display almost no variation in specimens 447/2 or 447/3, and cannot be useful in the determination of a new species.
Centre to centre calicle diameter is neither a good criterium to establish a new species. Only Blastochaetetes flabellum has distinctively small tubes (mean value: 0.28 mm), while dimensions from all other mentioned species or the specimens 447/2 or 447/3 are larger (from 0.37 to 0.60 mm, Table 1).
Double wall dimension is an important character, since it is distinctively similar between two groups of species. In B. flabellum and B.coquandi, its mean value does not exceed 0.07 mm in average, while in B. petri, Stromatoaxinella ir-regularis and in Blastochaetetes n. sp. A, the average value is at least 0.1 mm. In specimens 447/2 and 447/3, this character has, however, the highest minimum value (0.09 mm).
Tabular thickness is much more developed in specimens 447/2 or 447/3 than in most species of the genus. B. flabellum has the thinnest tabulae with 0.02 mm as the highest value, which is similar to B. petri. B. coquandi lies in the middle, reaching from 0.05 to 0.08 mm.
Some similarities exist between B. irregularis and both specimens 447/2 and 447/3 (Table 1). Nevertheless, the minimum value for Stromatoaxinella irregularis is 0.02 mm, being thus much lower than that of both 447/2 and 447/3 (0.07 mm). Maximum values also show differences between the two species.
Regarding the intertabular space, there are close similarities between B. flabellum and B. petri, in which this character has the lowest values (Table 1). In Stromatoaxinella irregularis, its value is higher, followed by B.coquandi. In both cases, its minimum is no more than 0.2 mm. However, although it reaches even more than 2 mm, in 447/2 and 447/3, this is the minimum value (Table 1). Nevertheless, this feature can seldom be seen due to the poor preservation.
Its growth form would help distinguish specimens 447/2 or 447/3 from species with similar internal morphological dimensions, like Stromatoaxinella irregularis and B. coquandi (Table 1). For S. irregularis, irregular growth forms can be seen. These include tuberose, branched and digitate forms (Fischer, 1970) as well as encrusting forms (Wood and Reitner, 1989). B.coquandi displays rounded forms (high-domical to bulbous, according to West and Clark, 1983), which can reach up to 30 cm in width and height and has a reduced flat fixation area (Fischer, 1970). On the other hand, specimens 447/2 and FCMP SI 447/3 measure only a few centimeters in height and width and can develop smooth and high-domical growth forms (Figures 7, 8, 11, 12). Nevertheless, growth form should not be considered to determine new species in most benthic organisms, since this character is prone to ecological variations (James and Bourque, 1992; Flügel, 2004). Furthermore, differences in the growth forms between specimens 447/2 and 447/3 should not be used as a valid character for distinguishing them as members of different species, since this character can be influenced by a vast array of ecological factors (James and Bourque, 1992).
It is widely accepted that spicules constitute a diagnostic character to identify sponges (Reitner, 1992). Many fossil chaetetids have been found to bear spicule pseudo-morphs (Dieci et al., 1977; Gray, 1980; Cremer 1994, 1995). However, neither 447/2 nor 447/3 bear any of them. There are even polychaete worms with a chaetetid growth form in the fossil record (Fischer et al., 1989). Nevertheless, as many spicules have been found in chaetetids of the genus Blastochaetetes (Dieci et al., 1977; Cremer, 1995), the taxonomical status of specimens 447/2 and 447/3 could eventually be solved and, thus, they could be recognised as poriferans.
Blastochaetetes flabellum (Michelin, 1845)
Chaetetes flabellum Michelin, 1845, p. 306, pl. 72, fig. 9 a, b; Félix, 1914, p. 249.
Polytrema flabellum d'Orbigny, 1850, p. 209, étage 21, no. 348.
Reptomulticava flabellum d'Orbigny, 1854, p. 1039, pl. 793, fig.1, 2; Gregory, 1909, p. 148 (partim).
Blastochaetetes flabellum (Michelin) Fischer, 1970, p. 182-184, fig. 23 a, b, pl. C, fig. 1-5.
Description. Specimen FCMP SI 48-2004-447/5
External characters. One chaetetid specimen was collected (FCMP SI 48-2004-447/5). It consists of a secondary skeleton measuring 7 cm in length and 5 cm in width. Height is 1 to 2 cm (Figures 9, 10). The chaetetid displays a pattern of concentric bands (common in calcified sponges and corals), which are located on the lower surface of the skeleton. These bands are parallel to each other and to the most external edge of the skeleton (Figure 9). Growth form is laminar, smooth and non-enveloping.
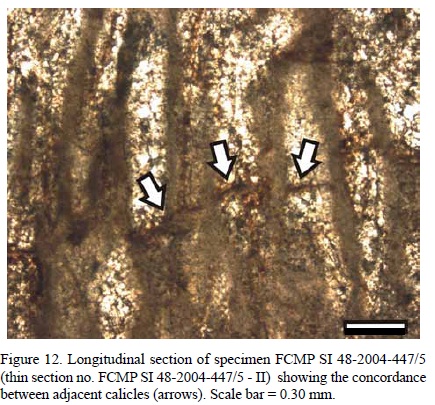
Internal morphological characters. Calicles are more or less rounded to oval (Figure 11) and can be found in average from 10 to 14 per mm2. Calicle diameter from center to center is ca. 300 µm. Internal calicle diameter ranges from 0.15 to 0.30 mm. Thickness of double walls varies from 0.06 to 0.15 mm, ca. 0.08 mm on average. No remains of the wall microstructure can be observed. Tabulae are arranged sub-horizontally. They measure 0.04 to 0.05 mm on average and are more or less concordant between adjacent calicles (Figure 12). The space between tabulae along the same calicle varies slightly, from 0.50 to 0.60 mm. There is neither evidence of any kind of calicle multiplication, nor the presence of spicule pseudomorphs.
Examined material: FCMP SI 48-2004-447/5 and two thin sections obtained from it, namely FCMP SI 48-2004-447/51 and FCMP SI 48-2004-447/5-2
Occurrence. "Senonian" of Alais, France; Coniacian-Santonian of San Luis Potosí, Mexico.
Discussion and conclusion. This material was assigned to the genus Blastochaetetes due to the distinctive and diagnostic discontinuities observed along the calicle walls. These discontinuities may be due to fissiparous division (Fischer 1970), although no true pseudoseptum could be found on transverse sections.
Species determination was difficult, since there are no visible hints of any microstructural wall characteristics and not all described characters are unique to any particular species of Blastochaetetes. Although most internal dimensions are similar to those of Blastochaetetes flabellum (Michelin), some variations exist and will be discussed next. Internal calicle diameter (0.15 to 0.30 mm) is slightly lower than the one indicated in the diagnosis of this species (0.20 to 0.35 mm from Fischer, 1970). In addition, more remarkable differences exist between the dimensions of tabulae thickness. Tabulae in the specimen analysed here range between 0.04 and 0.05 mm, being thus significantly thicker than those described by Felix (1914) and Fischer (1970).
Nevertheless, since the remaining diagnostic morphological dimensions coincide with those from the redescription of Fischer (1970), this specimen can be conferred to the species Blastochaetetes flabellum. This is reinforced by the presence of concentric bands on the upper surface of the colony, unique to this species.
PALAEOECOLOGY OF THE CHAETETID SPONGES FROM EL GORRIÓN: GROWTH MORPHOTYPES
It is not easy to perform inferences on the palaoecology of chaetetids due to the difficulty of extracting them from their frameworks. As this is not the case, it is therefore easy to determine the causes of the skeletons form from El Gorrión.
The growth record of colonial skeletons has a great potential to reveal the depositional dynamics in a short term of ancient marine benthonic environments. Growth form has been used for a long time as an indicator for environmental constraints (James and Bourque, 1992). In chaetetids, the laminar growth form can be interpreted as the early growth form of the fossil, which may develop into other growth forms as the individual grew; nevertheless, some chaetetids retain a laminar form throughout their lives. This form can be applied to other organisms, such as stromatoporoids, corals and bryozoans, because of the similarity in their skeletal development. However, the study of the ecology of fossil chaetetids remains scarce. They were most abundant in the Upper Carboniferous, where they inhabited biohermal limestones of shallow-depth waters (West and Clark, 1983). Apart from the Paleozoic, their fossil record is meager. Nowadays, some "chaetetiform" sponges preferably live in crevices and shadowy zones at a depth between 0 and 250 m (Rosenheim et al., 2004). Fossil chaetetids were influenced by a vast array of factors, including water depth, sedimentation and turbulence. These factors determined their external morphology and the size of their skeletons. Chaetetids can therefore be modelled as stacked laminar units, where from calicles can take different patterns which give form to the fully developed skeleton (Kershaw and West, 1991).
Kershaw and West (1991) interpret laminar growth form for chaetetids in two ways: If it has an enveloping nature, the colonies did not undergo considerable deposi-tional events in their lifetime, although they could have died during an abrupt depositional episode. On the other hand, if the laminar growth form is not enveloping, it could be interpreted as a basal stage of growth of the colony. Growth would focus on the apical part of the colony, which in the end would develop a domical form. Parallel to this, a slow and continuous deposition of the sediment would prevent growth at the sides of the colony. Blastochaetetesflabellum has a smooth non-enveloping growth form, so the second premise would seem true as a preliminary growth stage of a still unknown morphotype for this species. Laminar growth forms have been interpreted as resistant to water currents when the environment indicates a high-energy setting (James and Bourque, 1992; Flügel, 2004). Unfortunately, this cannot be defined, since it has not been determined if the chaetetids were fixed to the substrate. Therefore, no particular energy setting has been inferred.
The difference between the two observed morphotypes of Blastochaetetes sp. may have the following explanation: Although the two samples have a non-enveloping margin, the specimen 447/3 displays a ragged margin. Ragged margins, on the other hand, can be explained as an adaptation to high-energy environments, in which growth rate is very similar to sedimentation rate (James and Bourque, 1992; Flügel, 2004) and in which these ragged margins are a consequence of episodic sedimentation. On the other hand, some workers have considered this feature not to be reliable (Kershaw et al., 2006)
The high domical form of specimen 447/2, though non-enveloping, can be interpreted as an adaptation to a constant sedimentation rate environment (Kershaw and West, 1991) and/or due to the presence of an irregularity on the substrate (West and Kershaw, 1991).
FOSSIL RECORD OF CHAETETIDS IN THE AMERICAN CONTINENT
To date, only two chaetetid species have been described for Mesozoic in the American continent. The first species discovered was Blastochaetetes venezuelensis Wells, a chaetetid from the perireef limestones of the Barranquín Formation of Aptian/Albian age at Las Cinco Ceibas, Venezuela (Wells, 1944). The second one was Chaetetopsis krimholzi, a chaetetid sponge from the shelf-margin reef environments of the Abenaki Formation of Tithonian age at Nova Scotia, Canada (Jansa et al., 1982). The finding of both Blastochaetetes flabellum and the two specimens of Blastochaetetes sp. in El Gorrión therefore constitutes the first record of Mesozoic chaetetids for Mexico as well as the second one for both North America and Latin America, respectively.
Chaetetids were important and abundant reef builders during the Paleozoic (Suchy and West, 2001; May, 2008). This finding is thus of special importance, considering the scarceness of these kind of Mesozoic fossils.
ACKNOWLEDGEMENTS
This work is part of the M.Sc. project of the first author (2003-2005). FSB would like to thank Prof. Dr. Andrzej Pisera (Warszawa) for several valuable and helpful discussions and for his hospitality during a research stay at the Institute of Palaeontology of the Polish Academy of Sciences at the beginning of2004. The authors are thankful to Prof. Pisera for having reviewed two previous versions of this manuscript. We would also like to thank Dipl.-Geow. Vanessa Roden (Göttingen) and B.P. Carmen Sánchez-Beristain (Mexico City) for English language corrections. M.Sc. Miguel Ángel Basáñez-Loyola (Mexico City) provided valuable literature, for which we are beholden. M.Sc. Laura López-Esquivel Kranksith (Mexico City) is highly acknowledged for valuable discussions. The comments and suggestions of Dr. Steve Kershaw (Brunel) and Dr. Felix Schlagintweit (München) contributed to improve this manuscript substantially. We are much obliged to them. Dr. Schlagintweit also reviewed a previous version of this work, for which we are thankful. The present work was supported by both the Consejo Nacional de Ciencia y Tecnología (CONACYT, Grant No. 181707) and the Posgrado en Ciencias Biológicas de la Universidad Nacional Autónoma de México (UNAM). Both institutions funded the M.Sc. project of the first author.
REFERENCES
Ángeles-Villarreal, M., 2010, Mexico's free trade agreements: Washington, D. C., U.S.A., Congressional Research Service, reporte técnico, 22 pp. [ Links ]
Basáñez-Loyola, M.A., Fernández-Turner, R., Rosales-Domínguez, M.C., 1993, Cretaceous Platform of Valles-San Luis Potosí, Northeastern Central Mexico, in Simo, J.A., Masse, J.P., Scott, R.W. (eds.), Cretaceous Carbonate Platforms: American Association of Petroleum Geologists Memoir, 56, 51-59. [ Links ]
Carrillo-Bravo, J., 1971, La Platforma de Valles-San Luis Potosí: Boletín de la Asociación Mexicana de Geólogos Petroleros, 23 (1-6), 1-101. [ Links ]
Cremer, H., 1994, Zwei neue chaetetide Schwämme aus der Obertrias (Nor) von Südanatolien: Abhandlungen der Geologischen Bundesanstalt Wien, 50, 89-96. [ Links ]
Cremer, H., 1995, Spicule pseudomorphs in Upper Triassic (Norian) chaetetid sponges from the Western Taurids (Antalya-Region, SW Turkey): Geobios, 28, 163-174. [ Links ]
Cuif, J.P., Fischer, J.C., 1974, Etude systematique sur les Chaetetida du Trias de Turquie: Annales de Paleontologie, 60 (1), 1-14, pls. I-IV. [ Links ]
D'Orbigny, M.A., 1849-1854, Paléontologie francaise. Terrains Crétaces. V, Bryozoaires: Paris, Victor Masson, 1192 p. [ Links ]
D'Orbigny, M.A., 1850, Prodrome de Paléontologie Stratigraphique Universelle des Animaux Mollusques et Rayonnés, Vol. I: Paris, Victor Masson, 394 pp. [ Links ]
Dieci, G., Russo, A., Russo, F., Marchi, M.S., 1977, Occurrence of Spicules in Triassic Chaetetids and Ceratoporellids: Bolletino della Societá Paleontologica Italiana, 16(2), 229-238, pls 1-3. [ Links ]
Dietrich, W.O., 1919, Ueber sogennante Tabulaten des Jura und der Kreide, insbesondere die Gattung Acantharia Qu: Zentralblatt für Mineralogie, Geologie und Paläontologie, 1919, 208-218. [ Links ]
Felix, J., 1914, Fossilium Catalogus. I. Animalia. Pars 5-7. Anthozoa cretacea: Berlin, W. Junk, 273 pp. [ Links ]
Fischer, J.C., 1970, Révision et Essaie de Classification des Choetetida (Cnidaria) Post-Paleozoïques: Annales de Paléontologie, 56(2), 151-220, pls. A-F. [ Links ]
Fischer, R., Galli-Oliver, C., Reitner, J., 1989, Skeletal structure, growth and paleoecology of the patch reef-building worm Diplochaetetes mexicanus Wilson,1986 from the Oligocene of Baja California, Mexico: Geobios, 22(5), 761-775. [ Links ]
Flügel, E., 2004, Microfacies of carbonate rocks. Analysis, interpretation and application : Berlin- Heidelberg- New York, Springer Verlag, 984 pp. [ Links ]
Grant, R.E., 1836, Animal Kingdom, in Todd, R.B. (ed), The Cyclopaedia of Anatomy and Physiology. Volume 1, A-Dea: London, Longman, Brown, Green, Longman and Roberts, 107-118. [ Links ]
Gray, D.I., 1980, Spicule Pseudomorphs in a new Palaeozoic Chaetetid, and its Sclerosponge Affinities: Palaeontology, 23(4), 803-820. [ Links ]
Gregory, J. W. W., 1909, Catalogue ofthe fossil Bryozoa in the Department of Geology, British Museum (Natural History). The Cretaceous Bryozoa, Vol. II: London, The Trustees of the British Museum, 346 pp. [ Links ]
Hartman, W., Goreau, T.F., 1970, A new Pacific Sponge: homeomorph or descendent of the tabulate "corals"?: Geological Society of America, Abstracts with Programs, 2(7), 570. [ Links ]
Hartman, W., Goreau, T.F., 1972, Ceratoporella (Porifera: Sclerospongiae) and the "chaetetid" corals: Transactions of the Connecticut Academy of Arts and Sciences, 44, 133-148. [ Links ]
James, N.P., Bourque, P.A., 1992, Reefs and Mounds, in Walker, R.G., James, N.P. (eds.), Facies models- response to sea level change: St. John's, Geological Association of Canada, 323-347. [ Links ]
Jansa, L.F., Termier, G., Termier, H., 1982, Les biohermes à algues, spongiaires et coraux des series carbonateés de la flexure bordiere du "paleoshelf" au large du Canada oriental: Revue de Micropaleontologie, 25(3), 181-219. [ Links ]
Kazmierczak, J., 1979, Sclerosponge nature of chaetetids evidenced by speculated Chaetetopsis favrei (Deninger 1906) from the Barremian of Crimea: Neues Jahrbuch für Geologie und Paläontologie Monatshefte, 2, 97-108 [ Links ]
Kershaw, S., West, R.R., 1991, Chaetetid growth form and its controlling factors: Lethaia, 24, 333-346. [ Links ]
Kershaw, S., Wood, R., Guo, L., 2006, Stromatoporoid response to muddy substrates in Silurian limestones: GFF - Geologiska Föreningens i Stockholm Förhandlingar, 128, 131-138. [ Links ]
Levi, C., 1955, Les Clavaxinellides, Démosponges Tétractinomorphes: Archives de Zoologie Expérimentale et Générale, 93, 1-181. [ Links ]
May, A., 2008, Corals (Anthozoa, Tabulata and Rugosa) and chaetetids (Porifera) from the Devonian of the Semara area (Morocco) at the Museo Geominero (Madrid, Spain) and their biogeographic significance: Bulletin dell'Institut Scientifique Rabat, Section Sciences de la Terre, 30, 1-12 [ Links ]
Michelin, H., 1840-47, Iconographie zoophytologique. Description par localités et terrains des Polypiers fossils de France et des pays environnants: Paris, Bertrand, 348 pp. [ Links ]
Reitner, J., 1991, Phylogenetic Aspects and new Descriptions of Spicule-Bearing Hadromerid Sponges with a Secondary Calcareous Skeleton (Tetractinomorpha, Demospongiae), in Reitner, J., Keupp, H. (eds.), Fossil and recent sponges: Berlin, Springer Verlag, 179-211. [ Links ]
Reitner, J., 1992, "Coralline Spongien" Der Versuch einer phylogenetisch-taxonomischen Analyse: Berliner Geowissenschaftliche Abhandlungen, E1, 1-352, pl- 1-62. [ Links ]
Riding, R., 2004, Solenopora is a Chaetetid Sponge, Not an Alga: Palaeontology, 47, 117-122. [ Links ]
Riedel, P., 1990, Riffbiotope im Karn und Nor (Obertrias) der Tethys: Entwicklung, Einschnitte und Diversitätsmuster: Erlangen, Germany, Friedrich-Alexander-Universität Erlangen-Nürnberg, Geozentrum Nordbayern, tesis doctoral, 90 pp., pls. 1-15. [ Links ]
Rosenheim, B.E., Swart, P.K., Thorrold, S.R., Willenz, P., Berry, L., Latkoczy, C., 2004, High-resolution Sr/Ca records in sclerosponges calibrated to temperature in situ: Geology, 32(2), 145-148. [ Links ]
Sánchez-Beristain, J.F., 2005, Sistemática e Implicaciones Paleoecológicas de Esponjas y Espongiomorfos del Cretácico y Paleoceno de México: Ciudad de México, México, Universidad Nacional Autónoma de México, Facultad de Ciencias, tesis de maestría, 85 pp., pls. 1-3. [ Links ]
Sollas, W.J., 1885, A classification of the sponges: Annals and magazine of Natural History, 16(95) 395. [ Links ]
Suchy, D.R., West, R.R., 2001, Chaetetid buildups in a Westphalian (Desmoinesian) cyclothem in Southeastern Kansas: Palaios, 16(5), 425-443. [ Links ]
Wells, J.W., 1944, Cretaceous, Tertiary, and recent corals, a sponge, and an alga from Venezuela: Journal of Paleontology, 18(5), 429-447. [ Links ]
West, R.R., Clark, G.R.II, 1983, Chaetetids, in Broadhead, T.W. (ed.), Sponges and Spongiomorphs. Notes for a Short Course organized by J.K. Rigby and C.W. Stearn: Knoxville, Tennessee, University of Tennessee, Department of Geological Sciences, Studies in Geology, 7, 130-140. [ Links ]
West, R.R., Kershaw, S., 1991, Chaetetid habitats, in Reitner, J., Keupp, H. (eds.), Fossil and Recent Sponges: Berlin, Springer Verlag: 445-455. [ Links ]
Wood, R., Reitner, J., 1988, The "chaetetid" demosponge Stromatoaxinella irregularis (Michelin) and its systematic implications: Neues Jahrbuch für Geologie und Paläontologie Abhandlungen, 177, 213-224. [ Links ]

