










Serviços Personalizados
Journal
Artigo
 Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkRevista mexicana de ciencias geológicas
versão On-line ISSN 2007-2902versão impressa ISSN 1026-8774
Rev. mex. cienc. geol vol.29 no.2 Ciudad de México Ago. 2012
Taphonomy, sedimentology and chronology of a fossiliferous outcrop from the continental Pleistocene of Uruguay
Tafonomía, sedimentología y cronología de un yacimiento fosilífero del Pleistoceno continental de Uruguay
Andrea Corona*, Daniel Perea, Pablo Toriño, and César Goso
Departamento de Evolución de Cuencas, Facultad de Ciencias, Universidad de la República, Iguá 4225, 11400. Montevideo, Uruguay. *acorona@fcien.edu.uy
Manuscript received: January 28, 2012.
Corrected manuscript received: May 4, 2012.
Manuscript accepted: May 8, 2012.
RESUMEN
La fauna local La Paz (34°44' S, 56"17' W, elevación 31 m) constituye una asociación fosilífera con abundantes restos de mamíferos típicos del Pleistoceno del sur de Sudamérica. Esta asociación se encuentra conformando un banco osario de características únicas en Uruguay, el cual contiene una gran concentración de restos óseos en su mayoría completamente desarticulados y fragmentados, aunque también otros, que están muy bien conservados. Los principales objetivos de este trabajo son contribuir al entendimiento de los procesos sedimentarios y tafonómicos actuantes y aportar nuevos datos a la cronología del depósito. Los fragmentos óseos estudiados se presentan sin conexión anatómica, en su mayoría sumamente fracturados, y acumulados conformando un banco osario multidominante. El número de elementos identificados (NISP) es de 130. El 50% de las piezas estudiadas no presentó signos de meteorización; el 70 % mostró algún tipo de fractura, en la mayoría de los casos (40%) paralelas al eje principal; un escaso número de piezas mostró señales de abrasión (5%) y la evidencia directa indica que no existió selección (ni por forma ni por tamaño) por parte del agente de transporte. Los aspectos faciológicos, como la ausencia de estructuras sedimentarias–depósito dominantemente masivo–, la muy pobre grano–selección (clasificación), el elevado grado de angulosidad de los clastos mayores y las asociaciones de facies presentes permiten proponer que elfenómeno responsable del depósitofinal de los restos fue del tipo flujo de barro (mudflow) no canalizado. Las características tafonómicas también son compatibles con un proceso de este tipo. Los sedimentos portadores del banco osario son atribuibles a la Formación Libertad, asignada al Pleistoceno inferior–medio en la literatura precedente. Adicionalmente, se discute la validez de las dos únicas dataciones radiocarbónicas exitosas en el contexto de numerosos análisis fallidos. Los mamíferos de la fauna local La Paz y la presencia de ciertos minerales arcillosos permiten inferir, en el contexto de este depósito, un ambiente árido a semiárido.
Palabras clave: Cuaternario, banco osario, tafonomía, Formación Libertad, Uruguay.
ABSTRACT
The La Paz local fauna (34°44' S, 56°17' W, elevation 31 m) is a fossiliferous assemblage with abundant remains of typical southern South American Pleistocene mammals. The assemblage consists of a bonebed, containing a high concentration of skeletal remains, most of them completely disarticulated and fragmented, although very well preserved, and is unique in Uruguay as most remains have been found as single isolated finds until now. The goals of this study are to determine the sedimentary and taphonomic processes that produced this regionally unique concentration of bones and determine the age of the deposit. The bone fragments studied are not in their original anatomical position and consist of mostly, very fractured specimens accumulated in a multidominant bonebed. The number of identified specimens (NISP) is 130. Within the recovered sample, 50% of the studied specimens showed no signs of weathering; 70% showed some kind of fracture or breakage, many of the specimens (40%) parallel to the major axis; and a small number of specimens showed signs of abrasion (5%). Based on the sample there does not appear to have been selection (either by form or by size) by transport. The dominantly massive deposit lacks sedimentary structures, has poor sorting and a high degree of angularity of the largest pebbles suggesting that the final deposition of the remains resulted from a mudflow and were not channeled deposited. The taphonomic features observed on the bones are also compatible with this depositional process. The sediments in which the bonebed is located are attributed to the Libertad Formation, currently assigned to the lower–middle Pleistocene by other workers. We discuss the validity of the two available radiocarbon ages in the context of many failed essays. The mammals of the La Paz local fauna and the presence ofparticular clay mineral allow us to infer, in the context of this deposit, an arid to semi–arid environment.
Key words: Quaternary, bonebed, taphonomy, Libertad Formation, Uruguay.
INTRODUCTION
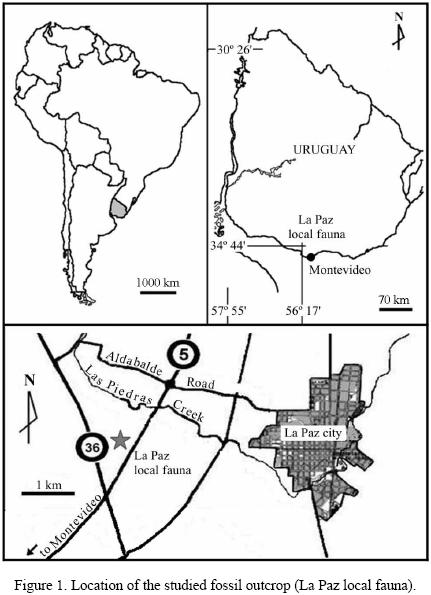
The La Paz local fauna (34°44' S, 56°17' W, elevation 31 m, Figure 1) is a fossiliferous assemblage with abundant remains of mammals typical of the Pleistocene in southern South America. This fauna was obtained from a preserved bonebed in a diamictite facies of the Libertad Formation (Pleistocene sensu lato) stratigraphically above a weathered granular layer of the La Paz Granite (Cambrian) located in a quarry.
Preliminary studies of the faunal composition of the association, its chronology, paleoenvironments and processes related to the genesis of the deposit have been previously discussed (Perea et al, 2001; Marchesano et al, 2002), whereas a detailed study of the site was initiated in 2005. This site is unique in Uruguay, as it contains a high concentration of skeletal remains, most of them completely disarticulated and fragmented, although other part is very well preserved. The fauna includes characteristic taxa of the Pleistocene of southern South America: Glyptodon sp., Doedicurus sp., Panochthus sp., Neosclerocalyptus sp., Lestodon sp., Macrauchenia cf. M. patachonica, Toxodon sp., Antifer sp., Stegomastodon waringi, among others. However, our analysis of the taphonomy of the deposit suggests that the constituent elements of the assemblage do not represent a single community.
Based on the taphonomic and sedimentological evidence (Perea et al., 2001; Marchesano et al, 2002; Corona et al., 2007; Corona et al, 2008a) the bonebed appears to have formed as a result of transport and deposition of the faunal remains during a high–energy event, like a mudflow. In addition, the presence of sedimentary facies that indicates remobilization and aeolic reworking in part of the sedimentary succession, show the occurrence of successive cycles of transport of the bones and sedimentation in a continental environment.
Based on geological mapping of the area, it was initially proposed that the lithostratigraphic unit that includes the bonebed was the Dolores Formation (Late Pleistocene) (Perea et al., 2001; Marchesano et al., 2002). However, Corona et al. (2008a) considered that the bonebed, most likely, is located in the Libertad Formation taking into account that the studied section occupies upper topographic positions, the slightly undulating morphology, and its location on the 1:100000 scale geological map of the area in which the bonebed occurs (Coronel et al, 1988). In addition, Gutiérrez et al. (2005) obtained a radiocarbon age of 17620 ± 100 yrs BP for this local fauna, based on dental enamel of Stegomastodon waringi, while here we present new chronological data.
The main objectives of this study are to contribute to the understanding of taphonomical and sedimentary processes that produced this unique concentration of bones never before documented in Uruguay, as well as to provide new data regarding the age of the deposit. Specifically, we made a comprehensive analysis of the taphocenosis through its main parameters. We investigated the sedimentology of the section in order to identify various lithofacies and analyzed the mineralogy of the clay fraction. Additionally, we analyze and discuss some chronological aspects.
METHODS
Taphonomy
Following the procedures described in Beherensmeyer (1978), Lyman (1994) and Cladera et al. (2004), we examined the following taphonomical parameters of the bones: orientation, degree of disarticulation, weathering stages, amount of abrasion, fracturing and degree of selection of the remains. The first two parameters were studied in situ, while the others were determined in the laboratory following the removal of the bones from the bonebed.
Orientation
It means the position of the long axis of bones to determine if there is any preferential direction.
Disarticulation
It refers to the degree to which the original anatomical connections of skeletal parts were preserved. We distinguished: elements retaining their original anatomical connection and elements without anatomical connections.
Weathering
It involves the loss of material from bone surface as result of physical and chemical agents. Beherensmeyer (1978) recognized six states of the weathering process, but for our study we simplified this to three categories: 1 (intact bone, corresponds to the state 0 of Beherensmeyer); 2 (loss of superficial material, corresponding to Beherensmeyer's 1 and 2 stages) and 3 (loss of deep bone material, Beherensmeyer's stages from 3 to 5).
Fracturing
It refers to both the number and disposition of fractures present in the remains. It was established using the following categories for both, the number and kind of fracture: 0 (no fracture), 1 (one or more fractures across the maximum dimension), 2 (one or more oblique fractures to the maximum dimension); >2 (one or more fractures longitudinal to the maximum dimension).
Abrasion
It measures the softness of edges and loss of roughness by transport. The categories established were: 1 (intact bone), 2 (rounded bone), 3 (polished bone).
Selection
First was established the shape of the elements following the Zingg's criterion for gravel (Suguio, 2003), which can be applied considering fossils as bioclasts. According to the relations among the three axes, Zingg established four basic forms: discoid, spherical, cylindrical and ellipsoidal. The three axes of each specimen (a, b, c) perpendicular to each other, being a > b > c were measured and the data plotted similar to Zingg's diagram.
Simultaneously, we identified taxonomically and anatomically each specimen and established the age of the individuals (young, young adult, adult and senile) based on the stages of tooth eruption or wear of the dentition or for the post–cranial skeleton through size or degree of ossification–of other elements. Some of the most important specimens in terms of better preservation are described in "Systematic Paleontology".
The studied fossil materials (Table 1) are housed in the Colección Paleontológica de la Facultad de Ciencias, Montevideo, Uruguay (FC–DPV) and in the Colección Particular Luis Castiglioni (LRC).

Sedimentology
For the studied outcrop we built a detailed stratigraphic profile in which textures, sedimentary structures, geometry and fossiliferous content were indicated. Representative samples of each lithofacies were collected in order to carry out mineralogical studies. Samples were subjected to physical and chemical breakdown for separation by screening the clay fraction. Clay components were treated with conventional methods for dispersion (sodium hexametaphosphate) in order to analyze by X–ray diffraction. These tests were carried out at the Instituto de Física, Facultad de Ciencias, with a diffractometer PW3710 (Anode: copper, wavelength = 1.54056 Â). The sedimentological study was complemented with textural and compositional observation of the coarser fraction (sand and silt) through a binocular magnifier and petrographic microscope.
Chronology
Some of the better preserved specimens were selected to obtain radiocarbon ages. These analyses were performed in the Accelerator Mass Spectrometry Laboratory, University of Arizona and in LATYR, Universidad Nacional de La Plata.
SYSTEMATIC PALEONTOLOGY
Included here are brief descriptions of some of the most relevant materials in terms of better preservation (Figure 2).
Mammalia Linnaeus, 1758
Xenarthra Cope, 1889
Folivora Delsuc et al., 2001
Mylodontidae Gill, 1872
Lestodon Gervais, 1855
Lestodon sp.
Materials. LRC 621, right femur. LRC 838, almost complete skull with damaged anterior dorsal region.
Measurements. LRC 621. Length: 74 cm; width: 24 cm.
Comments. Both specimens are very large representatives of the genus.
Notoungulata Roth, 1903
Toxodontidae Gervais, 1847
Toxodon Owen, 1837
Toxodon sp.
Material. LRC 469. Almost complete skull lacking incisors.
Measurements. Length: 53.5 cm; width: 35.8 cm.
Comments. It seems to be a young specimen because of the small size and the porous texture of some bones.
Carnivora Bowdich, 1821
Ursidae Gray, 1825
Tremarctinae Merriam y Stock,1925
Arctotherium Bravard, 1857
Arctotherium bonariense Gervais, 1852
Material. LRC 740, right mandibular fragment with p4, damaged m1 and m2.
Measurements. Teeth row length: 9.8 cm; mandible depth between m1 and m2: 7.3 cm; p4: 1.9/2.2 cm; m1: 2.6/4.5 cm; m2: 2.3/3.6 cm.
Comments. The teeth morphology allows us to recognize the species, being this specimen a very large representative of it.
RESULTS
Taphonomy
The studied bone fragments that constitutes the single bonebed are disarticulated from their original anatomical positions, most are highly fractured, and accumulated in a chaotic way with no preferred orientation and densely packed. Overall, this bonebed, which shows lobular geometry, has a lateral continuity of about 50 m and a maximum thickness of 1.0 m. The highest density of specimens is found in the central area, decreasing significantly towards the marginal areas, from where the best preserved fossils (skulls, jaws and the most complete long bones) were exhumed (Figure 3).
According to Rogers et al. (2007) this is a macrofossil bonebed (most bioclasts are more than 5 cm in maximum dimension). Also, it is a high diversity bonebed (10 or more taxa), and multidominant, since more than two taxa contribute to the total number of elements identified (NISP). Assuming that the probability of association among the elements is zero due to the high degree of disarticulation observed and the possible physical and/or temporary mixture, it is possible to obtain the number of individuals from the NISP (sensu Rogers et al., 2007). We obtained a minimum of 130 individuals, mostly representing herbivorous and omnivorous forms. The carnivores are represented by the short–faced bear Arctotherium bonariense (Corona et al., 2008b) and a phorusracid bird (Alvarenga et al., 2010). It should be noted that, due to the very fragmentary nature of the specimens, many could not be anatomical and/or taxonomically identified, therefore they were not included in the quantitative taphonomic analysis.
We identified two taphonomic modes (sensu Beherensmeyer and Hook, 1992) within the bonebed: the first and most frequent consists of highly fragmented and fragile bones with a characteristic yellowish white color, usually with dark spots on the surface resulting from mineral precipitation during diagenesis. The second is formed by very consolidated, black, small bones with rounded vertices and edges, indicating longer transport time and possible selection by size. Many of the bone specimens have gravel–size clasts adhering to its surface, with calcium carbonate either in the form of crusts (concretions) or as cement. The crusts and carbonate nodules may be formed after deposition during burial, mainly during early diagenetic time. It looks like randomly dispersed over the section, but they are mainly concentrated or nucleated around the bonebed. This indicates the early time diagenetic dissolution and precipitation of calcite around fossils. In addition, many of the fossils provide evidence of relatively long periods of exposure to erosive agents, while others have intact surfaces.
We describe below the main results of the taphonomic analysis (Figure 4):
Weathering
Fifty percent of the specimens did not show weathering effects (stage 1), while 40% showed loss of superficial bone (stage 2), but not affected the entire bone surface evenly. Only 10% of the materials were more affected, with loss of part of spongy bone (stage 3).
Fracturing
Seventy percent of the specimens had some type of fracture. In 40% of the cases the fractures are parallel to the main axis. Transverse and oblique fractures occurred in less proportion, 20% and 10% respectively.
Abrasion
The majority of fossils (95%) showed no evidence of abrasion. A very small percentage (5%) showed a marked erosion of reliefs and edges, attributed to category 2. There were no materials attributable to category 3 (polished bone).
Selection
There was no sorting during transport, as included in the mud debris deposit are both small (about 3 cm, e. g. Phorusracidae tarsometatarsus) and large (80 cm, e. g. Lestodon femur). As shown in Figure 4d, all the studied elements were distributed randomly in the Zingg diagram, which implies that there was no selection by shape.
Sedimentology
The complete stratigraphic profile (Figure 3) allowed the recognition of distinct depositional events. The studied section shows three lithofacies.
Lithofacies 1
Deposits of diamictite and mud with relatively well–defined, thin horizontal lamination, rest unconforrmably on the La Paz Granite (Cambrian). These sediments are dark grey–brown (10YR 4/2), 2 m thick, have a tabular geometry and consist of silty claystone with black spots, with very scattered angular clasts from the granite below, that varies in size from gravel to fine–coarse sand made of quartz, feldspar, and biotite. Associated with this level are rizholiths, concretions and thick crusts of calcium carbonate. Toward the top are interspersed lenses of thin, green clay.
Lithofacies 2
In abrupt contact above the previous lithofacies is a white (10 YR 8/1), matrix–supported diamictite, with relatively tabular geometry, and variable thickness between 2.0 and 2.50 m. Very angular megaclasts made of quartz, are abundant; they vary in size between gravel and pebble. The matrix is predominantly clayey containing a few sheets of biotite. At the bottom of this lithofacies the bonebed is found and consists of abundant remains of Pleistocene mammals, which show a relatively large lateral distribution (tens of meters). Above the bonebed appears a level of hard crust composed of calcium carbonate.
Lithofacies 3
The contact with the underlying unit is abrupt. It consists of a brown (7.5YR 5/4), approximately 1.50 m thick, sludge (predominantly silt clayey fraction) with tabular geometry, massive to slightly laminated structure and some oxide spots. This sludge includes some well–sorted, fine to medium sand composed of rounded to subrounded quartz.
The results of X–ray diffraction of samples of the three lithofacies show a predominance of chlorite and smectite over sepiolite. In particular, the lithofacies 2 (where is the bonebed) contains chlorite and sepiolite.
Chronology
The number of skeletal elements of this association is high, but the fragmentary nature of most of them do not allow a determination to a specific level (Appendix). However, the bonebed does not consist of taxa with stratigraphic or chronologic range restricted to a single Quaternary chronostratigraphic unit (sensu Cione y Tonni, 2005). Stegomastodon waringi andArctotherium bonariense are restricted to the Bonaerian–Lujanian stages (late middle Pleistocene–early Holocene). It is noteworthy that from dental enamel of Toxodon a new age was obtained for this local fauna: 5917 ± 65 years BP. This one and that performed by Gutiérrez et al. (2005) were the only data obtained in the context of a lot of failed analysis and cannot be considered indicative of the true age of the bonebed (Table 2).
DISCUSSION
Taphonomy
The taphonomic features of the bonebed are indicative of a deposit generated by a high energy process, such as a mudflow. The degree of weathering observed in the fossils (only 50% of materials analyzed) indicates that exposure prior to burial was long enough to allow complete disarticulation and dispersal of skeletal components, but not long enough to produce deep weathering signs.
However, as shown above, note that most of the fossils presented fractures parallel to the main axis of the specimens. According to Haynes (1980) and Alcalá (1994) fractures of this type are due to processes such as contraction and expansion of the bones during biostratinomic stage, prior to final burial. Therefore, we could infer only from this result that the subaerial exposure period was quite long, which is not consistent with the previous observation on the lack of deep weathering of the bone surface. According to Merino–Cristóbal (2000), fractures that occur prior to burial show cracks filled of sediment or some kind of cement, which was observed in many of the fossils from the La Paz local fauna. Transverse and oblique fractures occurred in a smaller number of specimens. The first fracturing event, presumably, occurred during diagenesis (Fernández–López, 2000), while there is no agreement among researchers on the factors responsible for the second event: action of carnivores, fluvial transport or trampling have been proposed (Cladera et al., 2004).
The observation and analysis of degree of weathering and type of fractures are mutually inconsistent results: the first parameter indicates a short time of subaerial exposure; the second suggests otherwise. However, the effects of weathering of the bone can be mitigated if the remains are under conditions of high humidity and protected by abundant vegetation (Beherensmeyer, 1978) which, in the current context cannot be excluded. Moreover, the presence of various stages of weathering may also indicate the occurrence of more than one taphonomic history, either by accumulation over long periods or concentration under different conditions after the initial weathering (Beherensmeyer, 1978; Rogers et al., 2007).
The small number of specimens that exhibit abrasion (5%) indicates that the impact on these materials, or other debris making up the load, was low. This observation, together with direct evidence that there was no selection (either by shape or by size) during transport, is compatible with the hypothesis of the occurrence of a high energy event responsible for the deposition. Furthermore, the characteristics of the bonebed in terms of massive and chaotic arrangement of its components and the presence of fossils from different backgrounds are consistent with a high energy gravity flow. The mixture of fossils with different taphonomic histories indicates that this assemblage is parautochtonous (remains reworked in the depositional site) or allochtonous (formed by remains transported from other areas) or a combination of both.
Sedimentology
The characteristics of the diamictite like an internal massive bedding or absence of sedimentary structures, poor sorting, erosive contact at the base, high angularity of the largest clasts, the bioclast supported texture, the tabular extent geometry and, most important, the facies associations and the continental environment involved, allow to suggest that the final deposition occurred by non–channelized, subaerial mudflow processes. The lithology and facies arrangement are consistent with that described by Hauser (1985) and Lopes Da Silva et al. (1997). This type of process permits to gravel clasts (or in this case bio–clasts or bones) to move in a cohesive, very thick matrix of clay, silt and water. These flows result in the generation of massive deposits, without structures and with poor sorted clasts, scattered throughout and supported by a pelitic matrix (Major, 2003). At present this type of deposits are typically generated by mudflows in alluvial fans and alluvial environments, especially when the discharge is significant, due to abundant rainfall (Eberth et al., 2006).
Lithostratigraphy
With regard to the designation of the bonebed to a distinct lithostratigraphic unit, it was recently proposed that the sediment containing the La Paz local fauna is part of the Libertad Formation (Corona et al., 2008a). However, this assignment has varied depending on the author. Marchesano et al. (2002) proposed that the sedimentary strata which includes this local fauna belongs to Dolores Formation. Subsequently, we used a more informal name, designating these deposits as "diamictites sediment of Libertad–Dolores(?) Formation" (Corona et al., 2007).
More generally, different authors have had difficulty assigning a formal name to the lithostratigraphic unit (Dolores Formation or Libertad Formation) for the Quaternary fossiliferous deposits in the southern part of Uruguay. The Dolores Formation (sensu Goso, 1972) is characterized by brown with gray–green local shades, silty clay, silt, clay, sand and gravel, associated with plains (Martínez and Ubilla, 2004). The Libertad Formation (sensu Goso, 1965) in contrast, consists of mudstones, sandy loess and reddish brown with calcium carbonate multiforms and undulate geomorphological features. This general lithologic similarity between both units is the cause of the problems to differentiate these two formations. In order to apply a consistent, formal designation, it essentially requires to apply geomorphologic criteria to distinguish them.
X–ray diffraction analysis of the clay fraction of the Quaternary continental deposits, confirmed the presence of smectite and illite in sediments of the Dolores Formation. On the other hand the Libertad Formation also contains these clays and, in addition, kaolinite (Goso and Ledesma, 2002; Goso et al., 2002). The sediments containing the La Paz local fauna consist of smectite, chlorite, and sepiolite (Corona et al., 2008a), which demonstrates that this methodology is not conclusive in the absence of major mineralogical differences between these two lithostratigraphic units. Therefore, although this technique contributes to the lithological characterization, for the moment does not distinguish between the two formations.
Chronology
Traditionally it has been assumed that the Libertad Formation was lower middle Pleistocene in age, although most of the previous works reaching this conclusion have not been stratigraphically controlled (Martínez y Ubilla, 2004). Although in the La Paz local fauna there are no taxa biostratigraphically restricted to a single Quaternary chronostratigraphic unit (sensu Cione y Tonni, 2005), the presence of Stegomastodon waringi and Arctotherium bonariense suggest a correlation with Bonaerian–Lujanian stages (late middle Pleistocene–early Holocene). Recently, substantial changes to the systematics of South American gomphotheres have been proposed by Mothé et al. (2011): Stegomastodon is a junior synonym of Notiomastodon, which includes all South American gomphotheres with a single species: Notiomastodon platensis (early Pleistocene–early Holocene). We agree with the conclusions of Alberdi and Prado (1995), Prado et al. (2005) and Alberdi et al. (2011), considering that Stegomastodon waringi a valid species which was distributed during middle to late Pleistocene in Ecuador, Brazil and Uruguay.
Unfortunately the sedimentary and genetic characteristics of this deposit does not allow the use of alternative dating methods such as Optically Stimulated Luminescence or Thermoluminescence which had been widely used to date other Quaternary units. Therefore, one of the goals of our work was to obtain samples that could permit obtaining new radiocarbon ages. Collagen (organic matter present in a ratio of 18% in the bone) is rapidly degraded, often making it very difficult to obtain enough material for accurate dating (Gupta and Polach, 1985). Another major difficulty related to the dating of bones by the method of 14C is the potential presence of exogenous carbon contamination, either through organic molecules mobilized from the surrounding sediment or through metabolic products of microorganisms. Contamination is very difficult to detect (especially if diagenesis is severe and the quality of preservation of the materials is not good) and result in younger age than the true one (Hedges and Van Klinken, 1992; Arslanov and Svezhentsev, 1993). All samples from the La Paz local fauna, except one, did not preserve collagen (probably because it suffered extensive diagenesis), so they could not be dated. Following strict protocols, a new sample from the same specimen of Stegomastodon waringi, which was dated as 17620 ± 100 years BP (Gutiérrez et al., 2005), was removed. The mandibular bone was perforated and a sample of 5 g of enamel was extracted from a not yet erupted tooth, superficially appeared unaltered and without fractures. However, the analysis determined that the sample lacked collagen, which contradicts the previous result.
An unexpected date of 5917 ± 65 years BP from dental enamel of Toxodon, a large difference with previously published age 17620 ± 100 years BP (Gutiérrez et al, 2005) and the absence of collagen in the remaining samples (including a specimen previously dated), suggest that the validity of both absolute ages is highly questionable.
Paleoenvironment and paleoclimate
There is no agreement among authors regarding the type of climatic conditions that existed at the time of deposition of Libertad Formation. The ranges of suggested climatic conditions that have been proposed include wet periods, torrential conditions and glacial to semi–arid episodes (Martínez and Ubilla, 2004). Some of the clay minerals, allow us to infer the local environmental conditions in the context of this deposit. In particular, the genesis of sepio–lite has been linked to highly alkaline environments (pH> 8), caused by groundwater with a high relative abundance of Mg indicative of semi–arid climatic conditions (Galán, 2006). Moreover, the mammals of the La Paz local fauna also allow reconstructing this environmental conditions since, in general, have been postulated as indicators of open savannah grasslands or arid to semi–arid environments (Tonni et al., 1999; Ortiz–Jaureguizar and Cladera, 2006).
CONCLUSIONS
The bonebed in which the La Paz local fauna is included is, until now, unique in Uruguay in terms of number and concentration of skeletal remains. The taphonomic parameters of the bonebed (disarticulation, orientation, weathering, abrasion, fracturing and selection) are indicative of a deposit generated by a high energy process, such as a mudflow. The mixture of fossils with different taphonomic histories indicates that this assemblage is parautochtonous or allochtonous or a combination of both.
The sedimentological features (massive deposit, absence of sedimentary structures, poor sorting, high degree of angularity of the largest clasts and facies associations) are also consistent with a mudflow but not in a channel. This phenomenon was the final agent responsible for the deposition of the bonebed.
The La Paz local fauna is included in sediments of the Libertad Formation. This fact represents a late middle Pleistocene–early Holocene age for that formation, based on its fauna. Regarding the absolute age of the deposit, the available data (5917 and 17620 yrs BP) cannot be considered indicative of its real age, but the sedimentary features and the intense diagenesis did not allow to obtain more data to be compared with the previous ones.
The mammals of the La Paz local fauna and the presence of particular clay minerals allow us to infer, in the context of this deposit, an arid to semi–arid environment.
ACKNOWLEDGEMENTS
The authors wish to thank the following people and institutions: Comisión Sectorial de Investigación Científica for providing partial financial support for the research. Alberto Silva for allowing access to his quarry. Luis Castiglioni by the provision and support. Leopoldo Soibelzon and Sergio Martínez for their comments and literature provided. Valeria Mesa, Andrés Batista, Felipe Montenegro and Edurné Cawen for their assistance in field and laboratory works. Gabriel Machado and Ernesto Goso collaborated on the mineralogical analysis. Martin Ubilla for useful suggestions made to the first version of this manuscript. H. Gregory McDonald helped us with language and provided useful comments. Edison Oliveira and another anonymous reviewer made comments that allowed improving this work.
REFERENCES
Alberdi, M.T., Prado, J.L., 1995, Los mastodontes de América del Sur, in Alberdi, M.T., Leone, G., Tonni, E.P. (eds.), Evolución biológica y climática de la Región Pampeana durante los últimos 5 millones de años; un ensayo de correlación con el Mediterráneo occidental: España, Museo Nacional de Ciencias Naturales, Monografías, 12, 277–292. [ Links ]
Alberdi, M.T., Prado, J.L., Ortiz–Jaureguizar, E., Posadas, P., Donato, M., 2011, Paleobiogeography of trilophodont gomphotheres (Mammalia: Proboscidea); a reconstruction applying DIVA (Dispersion–Vicariance Analysis): Revista Mexicana de Ciencias Geológicas, 28(2), 235–244. [ Links ]
Alcalá, L., 1994, Macromamíferos neógenos de la fosa de Alfambra–Teruel: Teruel, España, Museo Nacional de Ciencias Naturales, 554 p. [ Links ]
Alfaro, M., Perea, D., 2003, Nuevos registros fósiles de aves en Uruguay (Ciconiiformes, Ciconiidae y Gruiformes, Phorusrhacidae), en VII Jornadas de Zoología del Uruguay, Montevideo, Uruguay, Actas, 177 p. [ Links ]
Alvarenga, H.T., Jones, W., Rinderknecht, A., 2010, The youngest record of phorusrhacid birds (Aves, Phorusrhacidae) from the late Pleistocene of Uruguay: Neues Jahrbuch für Geologie und Palaontologie, Abhandlungen, 256(2), 229–234. [ Links ]
Arslanov, Kh.A., Svezhentsev, Y.S., 1993, An improved method for radiocarbon dating fossil bones: Radiocarbon, 35(3), 387–391. [ Links ]
Beherensmeyer, A.K., 1978, Taphonomic and ecologic information from bone weathering: Paleobiology, 4, 150–162. [ Links ]
Behrensmeyer, A.K., Hook, R.W., 1992, Paleoenvironmental contexts and taphonomic modes, in Behrensmeyer, A., Damuth, J., DiMichele, W., Potts, R., Sues, H.D., Wings, S. (eds.), Terrestrial ecosystems through time: Evolutionary paleoecology of terrestrial plants and animals: Chicago, Chicago University Press, 15–136. [ Links ]
Bowdich, T.E., 1821, An analysis of the natural classifications of mammalia for the use of students and travelers: Paris, J. Smith, 115 p. [ Links ]
Bravard, A., 1857, Observaciones geológicas sobre diferentes terrenos de transporte en la hoya del Plata: Buenos Aires, Imprenta y Litografía J.A. Bernheim, 80 p. [ Links ]
Cione, A., Tonni, E.P., 2005, Bioestratigrafía basada en mamíferos del Cenozoico Superior de la provincia de Buenos Aires, Argentina, in de Barrio, R.E., Etcheverry, R.O., Caballé, M.F., Llambias, E. (eds.), Geología y Recursos Minerales de la Provincia de Buenos Aires: La Plata, Argentina, Relatorio del XVI Congreso Geológico Argentino, XI, 183–200. [ Links ]
Cladera, G., Ruigomez, E., Ortiz–Jaureguizagar, E., Bond, M., López, G., 2004, Tafonomía de La Gran Hondonada (Formación Sarmiento, Edad–mamífero Mustersense, Eoceno Medio) Chubut, Argentina: Ameghiniana, 41(3), 315–330. [ Links ]
Cope, E.D., 1889, The Edentata of North America: American Naturalist, 23, 657–664. [ Links ]
Corona, A., Cawen, E., Toriño, P., Perea, D., Goso, C., 2005, Nuevos aportes al conocimiento de un yacimiento fosilífero del Pleistoceno de Uruguay, in VIII Jornadas de Zoología del Uruguay, Montevideo, 54. [ Links ]
Corona, A., Perea, D., Goso, C., Toriño, P., Castiglioni, L., 2007, Tafonomía de la "Fauna Local La Paz" (Pleistoceno Tardío) y mineralogía de sedimentos continentales asociados, Montevideo, Uruguay (cd): Actas del Quinto Congreso Uruguayo de Geología, Montevideo. [ Links ]
Corona, A., Goso Aguilar, C., Perea, D., 2008a, Sedimentología y mineralogía de los depósitos de la Formación Libertad (Pleistoceno Tardío) asociados a la Fauna Local La Paz (Montevideo, Uruguay): Revista de la Sociedad Uruguaya de Geología, 14, 11–17. [ Links ]
Corona, A. Perea, D., Ubilla, M., Castiglioni, L., 2008b, Primer registro de carnívoros (Ursidae, Tremarctinae) en la Fauna Local La Paz, Formación Libertad (Pleistoceno, Uruguay), in IX Jornadas de Zoología del Uruguay, Montevideo, 45. [ Links ]
Coronel, N., Veroslavsky, G., Goso, C., 1988, Carta Geológica del Uruguay hoja "Los Cerrillos–La Barra", Escala 1:100000: Dirección Nacional de Minería y Geología. [ Links ]
Delsuc, F., Catzeflis, F.M., Stanhope, M.J., Douzery, E.J.P. 2001, The evolution of armadillos, anteaters and sloths, depicted by nuclear and mitochondrial phylogenies: implications for the status of the enigmatic fossil Eurotamandua: Proceedings ofthe Royal Society of London B, 268, 1605–1615. [ Links ]
Eberth, D., Britt, B.B., Scheetz,R., Stadtman, K.L., Brinkman, D.B., 2006, Dalton Wells: Geology and significance of debris–flow–hosted dinosaur bonebeds in the Cedar Mountain Formation (Lower Cretaceous) of eastern Utah, USA: Palaeogeography, Palaeoclimatology, Palaeoecology, 236, 217–245. [ Links ]
Fernández–López, S., 2000, Temas de tafonomía: Universidad Complutense de Madrid, 167 p. [ Links ]
Galán, E., 2006, Genesis of clay minerals, en Bergaya, F., Theng, B.K., Lagaly, G., (eds): Developments in clay science, 1. Handbook of clay science, 1129–1162. [ Links ]
Gervais, P., 1847, Observations sur le mammifères fossiles du midi de la France: Annales des Sciences Naturelles, Zoologie 3(8), 203–224. [ Links ]
Gervais, P., 1852, Zoologie et Paléontologie françaises (animaux vertebrés) ou nouvelles recherches sur les animaux vivantes et fossiles de la France: Paris, A. Bertrand, 271p. [ Links ]
Gervais, P., 1855, Recherches sur les mammifères fossiles propres à l'Amérique méridionale: Comptes rendus des séances de l'Académie des Sciences, 40(20), 1112–1114. [ Links ]
Gill, T., 1872, Arrangement of the families of mammals with analytical tables: Smihsonian Miscellaneous Collections, 11(1), 1–98. [ Links ]
Goso, H., 1965, El Cenozoico en el Uruguay: Montevideo, Instituto Geológico de Uruguay, Informe interno, 36 p. [ Links ]
Goso, H., 1972, Cuaternario. Programa de Estudio y Levantamiento de Suelos: Montevideo, 12 p. [ Links ]
Goso, E., Ledesma, J., 2002, Formación Libertad: aportes para la caracterización como materia prima en la fabricación de cerámica roja, en II Jornadas Uruguayas del Cenozoico, Montevideo, 26–30. [ Links ]
Goso, E., Oyhantcabal, P., Spoturno, J., 2002, Granulometría y mineralogía de los depósitos Cenozoicos del este del Departamento de Montevideo, en II Jornadas Uruguayas del Cenozoico, Montevideo, 31–36. [ Links ]
Gupta, S.F., Polach, H.A., 1985, Radiocarbon dating practice at ANU. Radiocarbon Laboratory Research: Canberra, School of Pacific Studies ANU, 175 p. [ Links ]
Gutiérrez, M., Alberdi, M.T., Prado, J.L., Perea, D., 2005, Late Pleistocene Stegomastodon (Mammalia, Proboscidea) from Uruguay: Neues Jahrbuch für Geologie und Palaontologie, 11, 641–662. [ Links ]
Gray, J.E. 1825, An outline of an attempt at the disposition of Mammalia into Tribes and Families, with a list of genera apparently appertaining to each Tribe: Annals of Philosophy new series, 10, 337–344. [ Links ]
Hauser, A., 1985, Flujos de barro en la zona preandina de la región metropolitana; características, causas, efectos y medidas preventivas: Revista Geológica de Chile, número especial, 24, 75–92. [ Links ]
Haynes, G., 1980, Evidence of carnivore gnawing on Pleistocene and Recent mammalian bones: Paleobiology, 6, 341–351. [ Links ]
Hedges, R.E., Van Klinken, G.J., 1992, Review of current approaches in the pretreatment of bone for radiocarbon dating by AMS: Radiocarbon, 34(3), 279–291. [ Links ]
Linnaeus, C., 1758, Systema naturae per regna tria naturae, secundum classes, ordines, genera, species, cum characteribus, differentiis, synonymis, locis: Stockholm, Laurentii Salvii, Editio Decima, reformata, I, 824 p. [ Links ]
Lyman, R.L. 1994. Vertebrate taphonomy: Cambridge, Cambridge Manuals in Archaeology, 524 p. [ Links ]
Lopes Da Silva, E., Gama Junior, E., Goso, C., 1997, Fluxos gravitacionais de sedimentos: mecanismos de suporte, transformações de fluxos e procesos deposicionais: Geociencias, 16(2), 565–580. [ Links ]
Major, J.J., 2003, Debris flow in Middleton, G.V. (ed.), Encyclopedia of Sediments and Sedimentary Rocks, Dordrecht, 186–188. [ Links ]
Marchesano, M., Rovira, M., Castiglioni, L., Perea, D., 2002, Lista taxonómica actualizada y comentarios sobre un nuevo yacimiento con mamíferos pleistocénicos del departamento de Montevideo: II Jornadas Uruguayas del Cenozoico, Montevideo, 63–67. [ Links ]
Martínez, S., Ubilla, M., 2004, El Cuaternario en Uruguay, in Veroslovsky, G., Ubilla, M., Martínez, S. (eds.): Cuencas sedimentarias del Uruguay: geología, paleontología y recursos naturales –Cenozoico, Montevideo, 195–227. [ Links ]
Merino–Cristóbal, L., 2000, Mineralogía y geoquímica del esqueleto de mamíferos del Neógeno Español: Madrid, Consejo Superior de Investigaciones Científicas, Museo Nacional de Ciencias Naturales, 245 p. [ Links ]
Merriam, J.C., Stock, C., 1925, Relationships and structure of the short–faced bear, Arctotherium, from the Pleistocene of California: Contributions to Palaeontology from the Carnegie Institution of Washington, Papers Concerning the Palaeontology of the Pleistocene of California and the Tertiary of Oregon, Part 1, 35 p. [ Links ]
Mothé, D., Avilla, L.S., Cozzuol, M., Winck, G.R., 2011, Taxonomic revision of the quaternary gomphotheres (Mammalia: Proboscidea: Gomphotheriidae) from the South American lowlands: Quaternary International, in press. [ Links ]
Ortiz–Jaureguizar, E., Cladera, G.A., 2006, Paleoenvironmental evolution of Southern South America during the Cenozoic: Journal of Arid Environments, 66, 498–532. [ Links ]
Owen, R., 1837, A description of the cranium of the Toxodon platensis, a gigantic extinct mammiferous species, referrible by its dentition to the Rodentia, but with affinities to the Pachydermata and the herbivorous Cetacea: Proceeding of the Geological Society of London, 2, 541–542. [ Links ]
Perea, D., Lorenzo, N., Marchesano, M., Rovira, M., 2001, Un nuevo yacimiento con mamíferos cuaternarios del Uruguay: Revista Geológica Uruguaya, 1(1), 38–42. [ Links ]
Prado, J.L., Alberdi, M.T., Azanza, B., Sánchez, B., Frassinetti, D., 2005, The Pleistocene Gomphotheriidae (Proboscidea) from South America: Quaternary International, (126–128), 21–30. [ Links ]
Rogers, R.R., Eberth, D., Fiorillo, A., 2007, Bonebeds genesis, analysis, and paleobiological significance: Chicago and London, The University of Chicago Press, 499 p. [ Links ]
Roth, S., 1903, Los ungulados sudamericanos: Anales del Museo de La Plata (Sección Paleontología), 5, 1–36. [ Links ]
Suguio, K., 2003, Geologia sedimentar: São Paulo, Edgard Blucher Ltda., 400 p. [ Links ]
Tonni, E.P., Cione, A.L., Figini, A.J., 1999, Predominance of arid climates indicated by mammals in the pampas of Argentina during the Late Pleistocene and Holocene: Palaeogeography, Palaeoclimatology, Palaeoecology, 147, 257–281. [ Links ]

