










Serviços Personalizados
Journal
Artigo
 Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkRevista mexicana de ciencias geológicas
versão On-line ISSN 2007-2902versão impressa ISSN 1026-8774
Rev. mex. cienc. geol vol.29 no.2 Ciudad de México Ago. 2012
Lacustrine ostracodes from the Chihuahuan Desert of Mexico and inferred Late Quaternary paleoecological conditions
Ostrácodos lacustres del Desierto de Chihuahua, México y condiciones paleoecológicas inferidas del Cuaternario tardío
Claudia M. Chávez–Lara1, Priyadarsi D. Roy1*, Margarita M. Caballero2, Ana Luisa Carreño1, and Chokkalingam Lakshumanan3
1 Instituto de Geología, Universidad Nacional Autónoma de México, Ciudad Universitaria, 04510, México D.F., México. *roy@geologia.unam.mx
2 Instituto de Geofísica, Universidad Nacional Autónoma de México, Ciudad Universitaria, 04510, México D.F., México.
3 Centre for Remote Sensing, Bharathidasan University, Tiruchirapalli, 620023, Tamil Nadu, India.
Manuscript received: February 7, 2012.
Corrected manuscript received: April 17, 2012.
Manuscript accepted: April 23, 2012.
ABSTRACT
The abundance and distribution of ostracodes in the upper 865 cm of a sediment core recovered from Babícorapaleolake allows reconstruction of paleoecological conditions in the western Chihuahuan Desert by inferring variations in temperature and salinity of the lake water over the last ca. 72 cal kyr BP. The ostracode assemblage consists of five species (Candona patzcuaro Tressler, Cypridopsis vidua Brady, Limnocythere bradburyi Forester, L. platyforma Delorme andL. Ceriotuberosa Delorme), and its abundance varies between 0 and 1441 valves/g. Low ostracode abundance suggests the presence of dilute lake water (salinity>100 ppm) with abundant fresh water inflow into the basin prior to ca. 57 cal kyr BP. During ca. 57–49 cal kyr BP salinity remained low (>100ppm), but the water column was characterized by cold temperatures (>4 °C). Sediments of this zone have higher organic carbon content. Conditions were optimal for ostracode productivity (abundance of44–1441 valves/g) during ca. 49–29 cal kyr BP, and this zone indicates fluctuating temperature (5–32 °C) and salinity (100–100,000 ppm). A warm water column (>13 °C) and salinity of >500 ppm are inferred during ca. 29–12 cal kyr BP. Sediments of the last 12 cal kyr BP lack preserved ostracodes, and precipitation of authigenic calcite indicates periodic desiccation of the lacustrine basin during the Holocene. The inferred paleoecological conditions in the western Chihuahuan Desert are synchronous with other proxy–based late Quaternary paleohydrological and paleoclimatic information obtained from southwestern USA and northern Mexico.
Key words: ostracode, paleoecology, Late Quaternary, Chihuahuan Desert, Mexico.
RESUMEN
La abundancia y distribución de ostrácodos en los 865 cm superiores de un núcleo sedimentario recuperado del paleolago de Babícora, permitió inferir variaciones de salinidad y temperatura del cuerpo de agua en los últimos 72 ka cal AP y, por tanto, las condiciones paleoecológicas de la parte occidental del Desierto de Chihuahua. El grupo faunístico de ostrácodos se compone de cinco especies (Candona patzcuaro Tressler, Cypridopsis vidua Brady, Limnocythere bradburyi Forester, L. platyforma Delorme y L. ceriotuberosa Delorme) y su abundancia varía entre 0 y 1441 valvas/g. Durante >57 ka cal AP, los ostrácodos son escasos sugiriendo un lago diluido (salinidad>100 ppm) debido al flujo abundante de agua hacia la cuenca. Se infieren condiciones frías (>4 °C) con salinidad de >100 ppm durante ca. 57–49 ka cal AP, mostrando una alta productividad de materia orgánica. Condiciones óptimas para los ostrácodos (abundancia de 44–1441 valvas/g) se observan durante el intervalo de ca. 49–29 ka cal AP, donde se infieren fluctuaciones de la temperatura (5–32 °C) y la salinidad (100–100,000 ppm) del agua. Durante ca. 29–12 ka cal AP, la asociación de ostrácodos sugiere condiciones cálidas (>13 °C) con salinidades de >500 ppm. Los sedimentos de los últimos 12 ka cal AP no contienen ostrácodos mientras que la precipitación de calcita autigénica indica diferentes períodos de desecación de la cuenca durante el Holoceno. Las observaciones paleoecológicas del Cuaternario tardío inferidas de la parte occidental del Desierto de Chihuahua son comparables con la información paleohidrológica y paleoclimática obtenidas del suroeste de EUA y norte de México.
Palabras claves: Ostracoda, paleoecología, Cuaternario tardío, Desierto de Chihuahua, México.
INTRODUCTION
In recent times, understanding the influence of climate change and its effect on ecosystems has become a challenge. The limitation of instrumental records, reaching up to the past century, has led to the growth in interest in generating proxy–based records from geological deposits such as lacustrine and marine sediments in order to document long–term changes in the climatic system. Ostracodes are one of the most important biological proxies to infer late Quaternary paleoecological conditions. They are small crustaceans (with length up to 3 mm); their soft body is covered by two valves composed of low–Mg calcite joined at the dorsal part and forming the shell or carapace (Turpen and Angell, 1971). Diversification and adaptation to different environmental conditions are the reasons behind their presence in a wide variety of water bodies such as lakes, estuaries, swamps, lagoons, as well as oceans.
The ostracode valves preserved in lacustrine sediments are useful in inferring limnological conditions as their occurrence and assemblage are controlled by water chemistry, salinity and seasonal variation in temperature (Forester, 1986). Palacios–Fest and Dettman (2001) observed a linear correlation between Mg/Ca in valves and water temperature of live–collected Cypridopsis vidua from El Yeso lake in northern Mexico. Similarly, Bridgwater et al. (1999) observed an influence of salinity and host water chemistry on the trace–element composition of ostracode valves in short sediment cores from Pátzcuaro lake (central Mexico). Turbidity, substrate texture, water depth, bottom water oxygen concentration, food supply and predators are some among other factors affecting the productivity of ostracodes in lacustrine environments (Forester, 1986; Delorme, 1989; Holmes, 1992).
In the last several decades, the total abundance of ostracode populations, species diversity, geochemistry, and the structures of ostracode valves have been used as effective tools in reconstruction of past environmental conditions (De Deckker, 1988; Carreño, 1990; Forester, 1991; Palacios–Fest et al., 1993, 2002). For example, Chivas et al. (1993) studied both the elemental and stable isotope (carbon and oxygen) composition of ostracode valves in Holocene sediments from Keilambete lake (Australia) in order to understand varying salinity and lake–level history. Curtis and Hodell (1993) inferred millennial scale climatic variation by studying the geochemistry of ostracode valves preserved in sediments of the last 10.5 cal kyr BP from Miragoane lake (Haiti). In an application to geoarcheology, Palacios–Fest (1997) used Mg/Ca and Sr/Ca in valves of Limnocythere staplini from the Hohokam canal to reconstruct temperature and salinity for the last 1400 years in central Arizona, and suggested that the canals were mainly used during the late winter and pre–monsoon seasons.
In this paper, we discuss the abundance and distribution of ostracodes preserved in sediments of Babícora paleolake in order to extend the existing ostracode record from the Chihuahuan Desert and reconstruct late Quaternary paleoecological conditions of drylands of northern Mexico. In a previous study from the same lacustrine basin, Palacios–Fest et al. (2002) inferred the salinity and temperature of the lake water for the last 33 cal kyr BP by studying the ostracode assemblage. The present study extends the record up to 72 cal kyr BP. Additionally, we compared the distribution of ostracodes with the concentrations of total organic carbon (TOC) and inorganic carbon (TIC) in the bulk sediments, and also with earlier published pollen and diatom data (Metcalfe et al., 2002) in order to strengthen the Quaternary paleoclimatic information for the western Chihuahuan Desert. A regional perspective of late Quaternary paleohydrological and paleoclimatic conditions is possible by comparison with proxy records obtained from Baldwin lake, Estancia basin, and the Guadalupe mountains in southwestern USA, and from San Felipe paleolake and the Palomas basin in northwestern Mexico.
REGIONAL SETTING
The Chihuahuan Desert of Mexico includes the states of San Luis Potosí, Zacatecas, Durango, Coahuila, and Chihuahua. The Babícora paleolake (29°15'–29°30' N, 107°40'–108° W and ca. 2160 m asl) is located in a NW–SE oriented endorheic tectonic basin on the western margin of the Chihuahuan Desert (Figure 1) This lacustrine basin covers an area of ca. 1900 km2 and is surrounded by ca. 2200 to 3100 m high hills comprising Tertiary and Quaternary volcanic (basalt, rhyolite and ignimbrite) and sedimentary rocks (sandstone, siltstone and conglomerate). The meteorological station at Gómez Farías (eastern margin of the basin) registers mean annual precipitation of 550 mm; the western watersheds (ca. 1000 mm) receives higher precipitation compared to the eastern part (Metcalfe et al., 2002). The dominant summer rainfall (>70%) occurs as a result of monsoonal circulation and tropical cyclones bringing moisture from the Gulf of Mexico and Pacific Ocean (Schmidt, 1986). The winter rainfall derives from the middle latitudes of the Pacific (Metcalfe et al., 1997). The mean annual temperature varies between 4 °C (January) and 20 °C (July). According to the classification of Köppen, the region represents semi–cold climate (C(E)(w1)(x')(b')) (García, 1973).
The surrounding vegetation is altitude–dependent: conifer forests are found at elevations higher than 2400 m, juniper–oak woodland is located at 2400–2200 m, and tropical grassland lives below 2200 m (Ortega–Ramírez, 1995). The vegetation community consists of Pinus arizonica, Pinus chihuahensis, Pinus engelmanii, Quercus grisea and Quercus reticulata. The late Quaternary paleoecology and paleoclimatic information for this arid region comes mainly from micropaleontological assemblages in the lacustrine deposits of the Babícora basin. In addition, there are few studies of the sedimentology, mineral magnetism and geochemistry of this area (Urrutia–Fucugauchi et al., 1997; Ortega–Ramírez et al., 1998; Metcalfe et al., 2002). Metcalfe et al. (2002) inferred lake level variations between 65 and 10 cal kyr BP from diatom assemblages and reconstructed the paleovegetation during ca. 65–24 cal kyr BP from pollen preserved in the sediments. Similarly, Palacios–Fest et al. (2002) reported the paleoecological conditions of the last 33 cal kyr BP from the ostracode faunal association and Mg/Ca in valves of Limnocythere ceriotuberosa and Limnocythere platyforma.
MATERIAL AND METHODS
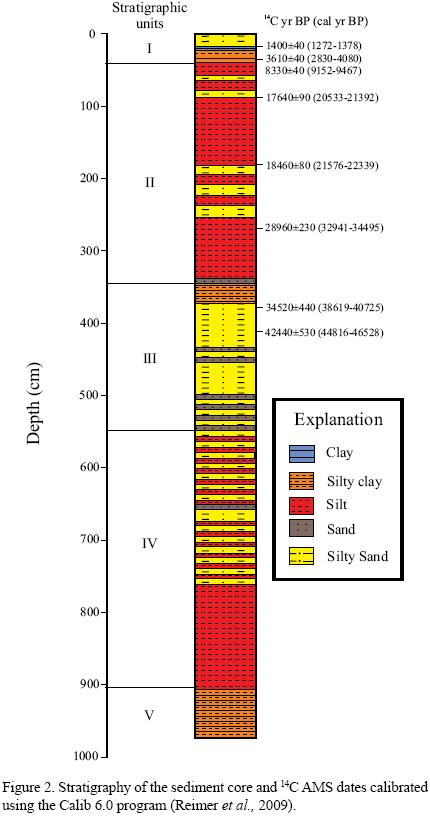
A 980 cm long sediment core (GF) was collected from the central eastern part (29°21'48.6" N, 107°48'09.0" W, 2143 m asl) of the basin using an Eijelkamp soil corer and transparent polycarbonate tubes (Figure 2). Chronology of the sedimentary sequence is based on eight AMS 14C dates on organic matter present in the bulk sediments. The age model was created after calibrating the 14C dates in the Calib 6.0 program of Reimer et al. (2009). The reservoir effect is regarded as negligible as 14C analysis was performed on bulk organic matter, and the lake watershed lacks carbonate rocks. Ages of samples taken between 17 and 409 cm depth were estimated by linear interpolation between two consecutive dates. It was not possible to date the sediments deposited before 45.6 cal kyr BP by AMS 14C analysis, so we made a linear extrapolation to assign an age to sediments below 409 cm.
For the ostracode analysis, a total of 87 samples were selected at intervals of ca. 10 cm between 0 to 865 cm depth. Due to the lack of ostracode valves, the lower part (980–865 cm) of the sediment sequence was not included in the study. The samples were oven dried at 50 °C and weighed to determine their dry weights. Sediments were disintegrated by soaking in water for 2–3 days and washed through two sieves of 0.063 mm and 0.425 mm to collect sediment with ostracode valves and eliminate the finer sediments. The residues were oven dried at 50 °C and examined under an Olympus SZX–FOF stereoscopic microscope to identify the species and calculate their abundance according to the descriptions provided by Van Morkhoven (1963), Delorme (1971 a, b, c), Forester (1985) and Carreño (1990). The total ostracode abundance in each sample was calculated by adding the number of adult and juvenile valves of all the observed species. The number of adult valves of each species was counted in 10 g of dry sediments and used for distribution calculations in percentage. The adult valves of each species were separated and placed in Plummer–type microfossil slides.
The concentrations of total organic and inorganic carbon in the bulk sediments were measured in 87 samples in a Thermo Scientific HiperTOC solid analyzer after oven drying (40 °C) and powdering the sediments using an agate mortar and pestle. Concentration of total carbon (TC) was measured by heating the sample to 980 °C, and inorganic carbon (TIC) was measured by acidifying the sediment with 10 % H3PO4. Accuracy and precision of both analyses are less than 5%. Total organic carbon (TOC) content was estimated by subtracting the inorganic carbon (TIC) from total carbon (TC).
RESULTS
Stratigraphy and chronology
The 980 cm long sediment core was divided into five successive stratigraphic units (Figure 2). We present here the stratigraphy of the upper 865 cm of the sediments. Unit IV (865–560 cm) contains centimeter–scale, brown to green silty sand and silt intercalations. Silt is dominant between 865 and 760 cm, and silty sand is abundant between 760 and 560 cm. Sediments of unit III (560–330 cm) comprise dark grey to brown silty sand and fine sand intercalations capped by a massive layer of brownish green silty clay. Layers of silty sand are found at depths of 515–505 cm, 445–438 cm, and 435–365 cm. Layers of fine sand are present at depths of 560–500 cm and 460–435 cm. The overlying unit II (330–40 cm) dominantly consists of silt along with silty sand. Intercalations of massive brown silt and silty sand are present between 330 and 180 cm, whereas weakly laminated brown silt is present between 180 and 40 cm. The uppermost unit I (40–0 cm) is composed of interbedded silty clay and clay (40–15 cm), and organic–rich, dark grey silty sand (15–0 cm). Vertical desiccation fissures filled with silty sand are found at depths of 20 to 15 cm. This unit also contains abundant plant roots.
Based on linear extrapolation of the AMS 14C dates, we constrain the entire sediment sequence (980–0 cm) between ca. 79 cal kyr BP and present day (Figure 2). The upper 865 cm of the deposit represents the last ca. 72 cal kyr BP. However, this is a rough estimate as linear extrapolation was used to assign dates to sediments deposited before 45.6 cal kyr BP. Sediments of unit IV (865–560 cm) were deposited during ca. 72–55 cal kyr BP, unit III (560–330 cm) represent ca. 55–37 cal kyr BP, and unit II (330–40 cm) is constrained between ca. 37 and 5 cal kyr BP. Similarly, the sediments of unit I (40–0 cm) represent paleoecological conditions of the last 5 cal kyr BP.
Abundance and distribution of ostracodes
Five different ostracode species were identified in the sediments (Figure 3): three of them belong to the genus Limnocythere (L. bradburyi, L. ceriotuberosa and L. platyforma) and the other two are Candona patzcuaro and Cypridopsis vidua. Most of the shells are well preserved. The juvenile shells (up to 1220 valves/g) are more abundant than the adult shells (up to 502 valves/g). Taphonomic features (i.e. ratios of carapaces/valves and juveniles/adults) were used to establish that the ostracode shells are autochthonous (Delorme, 1969, 1989; Forester, 1988). The total ostracode abundance varies between 0 and 1441 valves/g along the core, and we divided the profile into five different zones based on the variation in total ostracode abundance. Figure 4 summarizes the total ostracode abundance and distribution of adult shells of each species relative to depth. C. patzcuaro (0–453 valves/g) is the most abundant species throughout the record. It is followed by L. platyforma (0–171 valves/g), L. bradburyi (0–158 valves/g), L. ceriotu–berosa (0–85 valves/g) and finally C. vidua (0–48 valves/g).
The sediments of zone 5 (865–605 cm, ca. 72–57 cal kyr BP) have the lowest total ostracode abundance (0–27 valves/g), and most of these are juveniles (0–24 valves/g). This zone is also characterized by an absence of L. brad–buryi, C. vidua and L. ceriotuberosa. Adult shells of C. patzcuaro are found at depths of 835, 805, 755 and 645 cm. Only one adult shell of L. platyforma was observed at a depth of 705 cm. Zone 4 (605–455 cm, ca. 57–49 cal kyr BP) has an ostracode population of 7 to 419 valves/g. In this zone, L. platyforma shows its highest abundance (12–100 %) and is present at depths of 575 cm and 545–505 cm. C. patzcuaro (15–100 %) is present at depths of 595, 565, 525 and 485 cm. C. vidua (16–100 %) is observed at depths of 605, 555, 525 and 465 cm and L. ceriotuberosa (<20 %) is present at depths of 595, 575 and 515 cm. L. bradburyi (10 %) is only found at 525 cm depth.
Zone 3 comprises the depth interval 455–235 cm (ca. 49–29 cal kyr BP) and shows the highest ostracode abundance (44–1441 valves/g). In this zone, C. patzcuaro has its lowest abundance and comprises up to 90 % of the assemblage only at 255 cm. Both L. platyforma and L. bradburyi have comparable abundances and consist of 80 % and 50 % of the total abundance, respectively. C. vidua (up to 50 %) and L. ceriotuberosa (up to 40 %) are present in lower abundances. The lower part of this zone (455–395 cm, ca. 49–43 cal kyr BP) shows lower L. bradburyi and higher L. platyforma and C. vidua compared to the upper part. Zone 2 consists of sediments between 235 and 55 cm (ca. 29–12 cal kyr BP) with a total ostracode abundance of 2 to 398 valves/g. In this zone, C. patzcuaro (up to 100 %) and L. bradburyi (up to 100 %) are the most abundant species. L. platyforma (0–90 %) and C. vidua (0–90 %) have comparable abundances, and L. ceriotuberosa consists of 0 to 70 % of the total ostracode assemblage. The sediments from 55 to 0 cm depth comprise zone 1 (last 12 cal kyr BP), and do not contain ostracode valves.
Total inorganic carbon (TIC) and total organic carbon (TOC)
Figure 4 presents the distribution of TIC and TOC in the bulk sediments relative to depth. The concentration of TIC varies between 0.4 % and 1.7 % with depth. The highest values are observed at depths of 517 cm (1.2 %), 437 cm (1.6 %) and 275 cm (1.7 %). In the ostracode–free sediments, higher concentration of TIC is observed at depths of 57 cm (1.1 %), 37 cm (1.4 %) and 17 cm (1.4 %), respectively.
The TOC concentration in the bulk sediments varies between 0.2 % and 4.4 %. It shows a gradually increasing trend from 865 cm to 677 cm (0.2–3 %, ca. 72–61 cal kyr BP). Sediments at depths 677–400 cm (1.5–4.4 %, ca. 61–44 cal kyr BP) and 37–27 cm (1.7–2.8 %, ca. 2.5–3.5 cal kyr BP) show higher TOC values. In the rest of the sediments, it remains in the range of 0.4 to 1.6 % (400–37 cm, ca. 44–3.5 cal kyr BP).
DISCUSSION
The occurrence and association of ostracodes in a water body are controlled by temperature, salinity, dissolved oxygen content, pH and solute composition of the water as ostracode shells are composed of low–Mg calcite (Delorme, 1989). Each ostracode species is tolerant to a range of water salinity (total dissolved solids) and temperature, and their association indicates the variation in hydrochemical characteristics of the water column (Forester, 1986). For example, Candona patzcuaro and Cypridopsis vidua inhabit water bodies that are warm (13–32 °C) and have low salinity (100–5000 ppm), whereas Limnocythere platyforma inhabits cold (4–15 °C) and dilute (100–1000 ppm) waters. The presence of both Limnocythere bradburyi and Limnocythere ceriotuberosa indicates water bodies having a wide variation in temperature (4–32 °C). However, Limnocythere bradburyi (1000–100000 ppm) inhabits more saline waters compared to Limnocythere ceriotuberosa (500–10000 ppm; Delorme, 1989). Based on the ostracode association in the five different zones, we infer the water salinity and temperature of Babícora paleolake over the last ca. 72 cal kyr BP, and we support those interpretations with the concentrations of TIC and TOC (see below).
The solute composition of inflowing water is controlled by lithology. The watershed of Babícora paleolake is dominated by sandstone, siltstone, rhyolite, and basalt; and the weathering of these rocks produces waters rich in HCO3, Ca, Mg and Na (Eugster and Hardie, 1978). Due to the absence of carbonate rocks in the basin watershed, we consider the TIC values as the amount of authigenic carbonate precipitated from the saline water column (Cohen, 2003) as well as the abundance of ostracode shells. The TOC concentration is interpreted as the amount of organic matter preserved after sedimentation (Meyers and Teranes, 2001), and is considered as an indicator of productivity in the basin (Cohen, 2003). Next, we compare our observations with late Quaternary paleoclimatic records from other sites in southwestern USA and northern Mexico, in order to obtain a regional perspective.
Zone 5 (865–605 cm, ca. 72–57 cal kyr BP)
The total ostracode abundance of 0 to 27 valves/g can represent a water body with pH < 7. Carbonate is the principal constituent of ostracode shells, and it dissolves under low alkaline conditions (Delorme, 1989). Ostracodes are also rarely preserved in anoxic environments, and the gradually increasing TOC concentrations (0.2–3.8 %) in this zone possibly means decreasing oxygen availability in the water column. Dominance of the fresh water species (C. patzcuaro) and absence of species tolerant to saline water (C. vidua and L. ceriotuberosa) suggests the presence of a dilute water body with abundant fresh water inflow. This ostracode association inhabits a warm water body with temperature of 13–32 °C and salinity of >100 ppm. In a recent study on diatoms and pollen, Metcalfe et al. (2002) observed dominant Stephanodiscus niagarae, Juniperus and Quercus in the sediments representing ca. 65–57 cal kyr BP from the same basin. The diatom assemblage indicates the presence of a deep, fresh water lake, and the pollen association suggests expansion of the open oak and pine woodland to lower elevations. Broken and crumpled pollen grains along with other geochemical and magnetic proxies reflect the higher catchment erosion and deposition in a high energy environment (Metcalfe et al., 2002). However, the juvenile–dominant autochthonous ostracode shells do not provide any information about the energy conditions of deposition. The presence of other ostracode species during brief intervals in this zone suggests variation in the lake water salinity. For example, the occurrence of L. ceriotuberosa at 695 cm indicates increased lake water salinity (>500 ppm) and the presence of L. platyforma at depths of 705 cm represents dilute water (<1000 ppm). Variations in salinity of the lake water column occurred during ca. 63–62 cal kyr BP.
Zone 4 (605–155 cm, ca. 57–49 cal kyr BP)
The increasing ostracode abundance (7–419 valves/g) suggests presence of alkaline lake water (pH>7). Relatively higher TOC (1.6–3.4 %) possibly indicates higher organic productivity. L. platyforma shows its highest abundance in this zone, and suggests the presence of cold (4–15 °C) water compared to zone 5 with salinity of 100–1000 ppm. The presence of L. platyforma valves may also be related to lower total dissolved solids and low calcium concentrations (Keyser, 2005). Both C. vidua and L. ceriotuberosa are present in higher abundance compared to zone 5 and suggest that the lake water salinity increased (>500 ppm). C. vidua is a eurythermic species and prefers to live in warm (10–25 °C) wetlands (Palacios–Fest et al., 1993). This suggests that the water temperature was possibly higher than 10 °C during these periods. Based on the diatom association (Fragilaria construens and Cyclotella meneghiniana) and precipitation of authigenic calcite, Metcalfe et al. (2002) also inferred the presence of a shallow and chemically concentrated lake after 57 cal kyr BP.
Zone 3 (455–235 cm, ca. 49–29 cal kyr BP)
The highest abundance of ostracode shells (44–1441 valves/g) in this interval suggests that conditions were optimal for ostracode productivity. Abundances of L. platyforma and L. ceriotuberosa are lower compared to zone 4. However, abundance of L. bradburyi is higher. This indicates water with fluctuating temperature (5–32 °C) and salinity (100–100000 ppm) during ca. 49–29 cal kyr BP. The abundance of L. bradburyi is relatively lower in the lower part of the zone (455–395 cm) compared to the upper part (395–235 cm). Higher abundances of C. vidua and L. platyforma from 455 to 395 cm suggests a relatively dilute water body (salinity <1000 ppm) during ca. 49–43 cal kyr BP. Higher abundance of L. bradburyi at depths of 395 to 235 cm suggests a highly saline water body (>1000 ppm) during ca. 43–29 cal kyr BP. The gradually decreasing TOC content indicates that organic productivity possibly decreased with increasing lake water salinity. Fluctuating salinity is supported by the precipitation of authigenic calcite during evaporative phases, and the presence of Stephanodiscus niagarae during phases of deep water conditions (Metcalfe et al., 2002). Similarly, varying temperature is suggested by dominance of Quercus indicating warm conditions prior to 34 cal kyr BP, and abundant Pinus and Picea indicating cold conditions after 34 cal kyr BP (Metcalfe et al., 2002).
Zone 2 (235–55 cm, ca. 29–12 cal kyr BP)
The ostracode abundance decreases (2–398 valves/g) compared to zone 3, and the concentration of organic carbon (TOC: 0.3–1.4 %) is lower compared to zones 4 and 5. L. ceriotuberosa shows its highest abundance in this zone and its association with C. vidua suggests warm water (>13 °C) with salinity in the range of 500–4000 ppm. The relatively higher abundance ofL. bradburyi at depths of 185, 105 and 75 cm suggests that the salinity increased (>1000 ppm) at ca. 22 cal kyr BP and ca. 17 cal kyr BP. The diatom assemblage (Nitzschia inconspicua, N.frustulum and N. cuspidata) also suggests a shallow, saline, and turbid lake with oscillating lake levels. The presence of oxidized corroded pollen grains indicates periodic drying of the lake basin after ca. 24 cal kyr BP (Metcalfe et al., 2002).
Zone 1 (55–0 cm, last 12 cal kyr BP)
This zone is characterized by the absence of ostracode valves. Higher TIC values (1.1–1.4 %) at 57, 37, and 17 cm mirrors the precipitation of authigenic calcite from the lake water as a result of increasing salinity and temperature at ca. 12, ca. 3.5, and ca. 1 cal kyr BP. The vertical desiccation fissures (mud cracks) filled with silty sand at 20–15 cm depth represent drying of the basin for an extensive period of ca. 1 cal kyr BP. The absence of diatoms during the last 11 cal kyr BP, and pollen in the sediments of ca. 12–1.5 cal kyr BP also reflect the decrease in effective moisture and periodic desiccation of the lake during the Holocene (Metcalfe et al., 2002).
Comparison with the existing ostracode record
In a previous study of the same lacustrine basin, Palacios–Fest et al. (2002) identified seven different species in 230 to 298 cm short cores collected from the western, central and southern parts of the basin. They reconstructed the temperature of the water column by measuring Mg/Ca in valves of L. ceriotuberosa and L. platyforma present in sediments of the last 33 cal kyr BP. Contrary to the existing record, we did not observe the presence of either Candona caudata or Limnocythere sappaenis, and the Holocene sediments in the present register lack preservation of ostracode shells. As the sampling resolution in the present study is better compared to the previous study, we relate the absence of these two species in the late Pleistocene sediments, and the lack of ostracode shells in the Holocene sediments to the location of the sediment cores within the basin. Palacios–Fest et al. (2002) also did not observe L. ceriotuberosa in the core collected from central part of the basin and L. sappaensis was not present in the core collected from the southern part. The difference between their reconstructed temperature (5.6–9.9 °C) during ca. 33–13 cal kyr BP and that proposed in the present work (>13 °C in zone 2) could be due to different approaches in inferring temperature of the water column. Temperature and salinity inferences for the present study are based only on the association of os–tracode species, whereas the earlier study was based on the geochemistry of ostracode shells. Similarly, Palacios–Fest et al. (2002) also inferred a lake water temperature reaching up to 21.3 °C during ca. 9–4 cal kyr BP. The absence of ostracode shells in sediments of the last 12 cal kyr BP in the present study rules out any inference on the Holocene physico–chemical parameters of the Babícora water column.
Comparison with proxy–based regional paleoclimatic records
The inferred salinity and temperature of the Babícora paleolake water column may be compared with other proxy–based late Quaternary paleoclimatic records obtained from Baldwin lake, Estancia basin, and Guadalupe mountains in southwestern USA, and from San Felipe paleolake and the Palomas basin in northern Mexico (Menking and Anderson, 2003; Polyak et al., 2004; Castiglia and Fawcett, 2006; Kirby et al., 2006; Asmerom et al., 2007; Roy et al., 2010, 2012).
The inferred presence of dilute water prior to ca. 49 cal kyr BP at Babícora is contemporaneous with high lake levels observed at Baldwin lake in the coastal southwestern USA. The gradually increasing salinity and varying temperature inferred for Babícora paleolake after 49 cal kyr BP is comparable to the extreme wet–dry cycles (49–24 cal kyr BP) and shallow saline lake conditions (24–22 cal kyr BP) at Baldwin lake (Kirby et al., 2006). From 48 to 14 cal kyr BP, sediments of San Felipe paleolake in the western Sonoran Desert reflect lower pluvial discharge as a result of weak summer precipitation, and the saline conditions there led to precipitation of Na–sulfate minerals (Roy et al., 2010, 2012). Comparison with orbital–scale variations in winter–summer insolation at 30°N suggests that winter storms were strengthened during periods of lower winter insolation, and the magnitude and spatial extent of the North American monsoon increased during intervals of higher summer insolation (Kirby et al., 2006). During the onset of the last glacial maximum (24–22 cal kyr BP), the unfavourable conditions for precipitation based on insolation variation was diminished by an expanded North American ice sheet.
Zone 1 at Babícora paleolake, with its lack of ostra–code shells, may be compared with speleothem records from the Guadalupe mountains in southern Texas. Based on stalagmite growth rates, Polyak et al. (2004) identified dry conditions during ca. 14–12.5 ka and wet conditions between 12.5 and 10.5 ka. Wet conditions during the Younger Dryas were related to a southward shift in the jet stream and associated winter storms as a result of northern hemisphere cooling. The end of this wet period coincided with the Holocene summer maximum insolation. The oxygen isotope record obtained from another speleothem in the same region indicates that the dry period continued up to 7 ka, and negative excursions in δ18O indicate brief periods of increased precipitation (summer and winter) at ca. 6.2 ka, 3.3 ka, and 2.7 ka (Asmerom et al., 2007). Castiglia and Fawcett (2006) identified lake highstands and wet conditions during ca. 9.6–9, ca. 7.2–6.7, and ca. 4.9–4 cal kyr BP at the Palomas basin in northern Mexico. The synchronicity between Holocene wet conditions in the Palomas basin and cooler sea–ice drift events in the Northern Atlantic suggests that the Holocene lake highstands may have been influenced by periods of increasing frequency of El Niño modulated winter precipitation along with colder temperature and reduced evaporation rates. In the Estancia basin of central New Mexico, Menking and Anderson (2003) identified two distinct mid–Holocene droughts during ca. 7–5 cal kyr BP, and related them to enhanced La Niña.
CONCLUSIONS
In the absence of longer and continuous paleoclimatic records, the late Quaternary paleohydrological evolution of the arid regions in northern Mexico is not yet understood. In the present study, the total ostracode abundance and association of five different species identified in sediments from Babícora paleolake provide a qualitative estimation of lake water salinity and temperature over the last ca. 72 cal kyr BP. The presence of a dilute water column (salinity >100 ppm) before ca. 49 cal kyr BP was contemporaneous with high lake levels in the southwestern USA attributed to increased frequency of both summer as well as winter precipitation. A regime of fluctuating temperature (>4 to >13°C) and gradually increasing salinity (>100 to >1000 ppm) between ca. 49 and 12 cal kyr BP was contemporaneous with regional records suggesting weak summer precipitation in northwestern Mexico and wet–dry cycles in the southwestern USA. The absence of ostracode shells in sediments of the last 12 cal kyr BP at Babícora paleolake limits our understanding of Holocene paleohydrological changes.
ACKNOWLEDGEMENTS
The data presented in this paper were generated with the financial support from CONACyT Ciencias Básicas project 83800 and Papiit–UNAM project IN104610. CMCL acknowledges the scholarship from CONACyT for her postgraduate studies. The coring equipment was provided by Dr. Socorro Lozano, and technical assistance was provided during different stages from M.P. Jonathan, Rosalinda Hernández, Alejandro Rodríguez and Violeta Amparo Romeo. We are thankful to Steffen Mischke, Gabriela Cusminsky and Thomas M. Lehman for their critical comments and observations. This is a partial contribution from Earth System Science Group, India.
REFERENCES
Asmerom, Y., Polyak, V., Burns, S., Rassmussen, J., 2007, Solar forcing of Holocene climate: New insights from a speleothem record, southwestern United States: Geology, 35(1), 1–4. [ Links ]
Bridgwater, N., Holmes, J.A., O'Hara, S., 1999, Complex controls on the trace–element chemistry of non–marine ostracods: an example from Lake Pátzcuaro, central Mexico: Palaeogeography, Palaeoclimatology, Palaeoecology, 148, 117–131. [ Links ]
Carreño, A.L., 1990, Ostrácodos lacustres del paleolago de Texcoco: Revista de la Sociedad Mexicana de Paleontología, 3(1), 117–135. [ Links ]
Castiglia, P.J., Fawcett, P.J., 2006, Large Holocene lakes and climatic change in the Chihuahua Desert: Geology 34(2), 113–116. [ Links ]
Chivas, A., De Deckker, P., Cali, J., Chapman, A., Kiss, E., and Shelley, M., 1993, Coupled stable–isotope and trace–element measurements of lacustrine carbonates as paleoclimatic indicators, in Swart, P.K., Lohmann, K.C., McKenzie, J., Savin, S. (eds.), Climate Change in Continental Isotopic Record: Washington D.C., Geophysical Monograph, 78, 113–121. [ Links ]
Cohen, A.S., 2003, Paleolimnology: the history and evolution of lake systems: New York, Oxford University Press, 500 pp. [ Links ]
Curtis, J., Hodell, D., 1993, An isotopic and trace element study of ostracods from Lake Miragoane, Haiti: A 10,500 year record of paleosalinity and paleotemperature changes in the Caribbean, in Swart, P.K., Lohmann, K.C., McKenzie, J., Savin, S. (eds.), Climate Change in Continental Isotopic Record: Washington D.C., Geophysical Monograph, 78, 135–152. [ Links ]
De Deckker, P., 1988, An account of the techniques using ostracodes in paleolimnology in Australia: Palaeogeography, Palaeoclimatology, Palaeoecology, 62, 463–475. [ Links ]
Delorme, D.L., 1969, Ostracodes as Quaternary paleoecological indicators: Canadian Journal of Earth Sciences, 6, 1471–1476. [ Links ]
Delorme, D.L., 1971a, Freshwater ostracodes of Canada. Part II. Subfamily Cypridopsinae and Herpetocypridinae, and family Cyclocyprididae: Canadian Journal of Zoology, 48, 253–266. [ Links ]
Delorme, D.L., 1971b, Freshwater ostracodes of Canada. Part III. Family Candonidae: Canadian Journal of Zoology, 48, 1099–1127. [ Links ]
Delorme, D.L., 1971c, Freshwater ostracodes of Canada. Part V. Families Limnocytheridae, Loxoconchidae: Canadian Journal of Zoology, 49, 43–64. [ Links ]
Delorme, L.D., 1989, Methods in Quaternary ecology # 7. Fresh water ostracodes: Geoscience Canada, 16(2), 85–90. [ Links ]
Eugster, H.P., Hardie, L.A., 1978, Saline Lakes, in Lerman, A. (ed.), Lakes–chemistry, geology, physics: New York, Springer–Verlag, 237–294. [ Links ]
Forester, R.M., 1985, Limnocythere bradburyi n. sp; a modern ostracode from Central México and possible Quaternary paleoclimatic indicator: Journal of Paleontology, 59(1), 8–20. [ Links ]
Forester, R.M., 1986, Determination of the dissolved anion composition of ancient lakes from fossil ostracodes: Geology, 14, 796–798. [ Links ]
Forester, R.M., 1988, Nonmarine calcareous microfossil sample preparation and data acquisition procedures: U.S. Geological Survey Technical Procedure, HP–78, R1, 1–9. [ Links ]
Forester, R.M., 1991, Ostracode assemblages from springs in the western United States: lmplications for paleohydrology: Memories ofthe Entomological Society of Canada, 155, 181–201. [ Links ]
García, E., 1973, Modificaciones al sistema de clasificación climática de Köppen (para adaptarlo a las condiciones de la República Mexicana): Mexico, D.F., Boletín del Instituto de Geografia, 246 pp. [ Links ]
Holmes, J.P., 1992, Nonmarine ostracodes as Quaternary paleoenvironmental indicators: Progress in Physical Geography, 16, 425–431. [ Links ]
Keyser, D., 2005, Histological peculiarities of the noding process in Cyprideis torosa (Jones) (Crustacea, Ostracoda): Hydrobiologia, 538, 95–106. [ Links ]
Kirby, M.E., Lund, S.P., Bird, B.W., 2006, Mid–Wisconsin sediment record from Baldwin Lake reveals hemispheric climate dynamics (Southern CA, USA): Palaeogeography, Palaeoclimatology, Palaeoecology, 241, 267–283. [ Links ]
Menking, K.M., Anderson, R.Y., 2003, Contributions of La Niña and El Niño to middle Holocene drought and late Holocene moisture in the American southwest: Geology, 31(11), 937–940. [ Links ]
Metcalfe, S.E., Bimpson, A., Courtice, A.J., O'Hara, S.L., Taylor, D.M., 1997, Climate change at the Monsoon/Westerly boundary in Northern Mexico: Journal of Paleolimnology, 17, 155–171. [ Links ]
Metcalfe, S., Say, A., Black, S., McCulloch, R., O'Hara, S., 2002, Wet conditions during the Last Glaciation in the Chihuahuan Desert, Alta Babicora Basin, Mexico: Quaternary Research, 57, 91–101. [ Links ]
Meyers, P.A., Teranes, J.L., 2001, Sediment organic matter, in Last, W.M., Smol, J.P. (eds.), Tracking Environmental Change Using Lake Sediments. Volume 2: Physical and Geochemical Techniques: Dordrecht, Kluwer Academic Publishers, 239–269. [ Links ]
Ortega–Ramírez, J., 1995, Correlación estratigráfica de los depósitos cuaternarios de la Laguna de Babícora, Chihuahua, México: Geofísica Internacional, 34 (1), 117–129. [ Links ]
Ortega–Ramírez, J. R., Valiente–Banuet, A., Urrutia–Fucugauchi, J., Mortera–Gutiérrez, C.A., Alvarado–Valdez, G., 1998, Paleoclimatic changes during the Late Pleistocene–Holocene in Laguna de Babícora, near the Chihuahuan Desert, México: Canadian Journal of Earth Sciences, 35, 1168–1179. [ Links ]
Palacios–Fest, M.R., 1997, Paleoenvironmental reconstruction of human activity in Central Arizona using shell chemistry of Hohokam Canal Ostracodes: Geoarchaeology: An International Journal, 12(3), 211–226. [ Links ]
Palacios–Fest, M.R., Dettman, D.L., 2001, Temperature controls monthly variation in Ostracode valve Mg/Ca: Cypridopsis vidua from a small lake in Sonora, México: Geochimica et Cosmochimica Acta, 65(15), 2499–2507. [ Links ]
Palacios–Fest, M.R., Cohen, A.S., Ruíz, J., Blank, B., 1993, Comparative paleoclimatic interpretations from nonmarine ostracodes using faunal assemblages, trace elements shell chemistry and stable isotope data, in Swart, P.K., Lohmann, K.C., McKenzie, J., Savin, S. (eds.), Climate Change in Continental Isotopic Record: Washington, D.C., Geophysical Monograph, 78, 179–190. [ Links ]
Palacios–Fest, M.R., Carreño, A.L., Ortega–Ramirez, J.R., Alvarado–Valdez, G., 2002, A paleoenvironmental reconstruction of Laguna Babicora, Chihuahua, Mexico based on ostracode paleoecology and trace element shell chemistry: Journal of Paleolimnology, 27, 185–206. [ Links ]
Polyak, V.J., Rasmussen, J.B.T., Asmerom, Y., 2004, Prolonged wet period in the southwestern United States through the Younger Dryas: Geology, 32(1), 5–8. [ Links ]
Reimer, P.J., Baillie, M.G.L., Bard, E., Bayliss, A., Beck, J.W., Blackwell, P.G., Ramsey, C.B., Buck, C.E., Burr, G.S., Edwards, R.L., Friedrich, M., Grootes, P.M., Guilderson, T.P., Hajdas, I., Heaton, T.J., Hogg, A.G., Hughen, K.A., Kaiser, K.F., Kromer, B., McCormac, F.G., Manning, S.W., Reimer, R.W., Richards, D.A., Southon, J.R.,Talamo, S.R., Turney, C.S.M., Van Der Plicht, J., Weyhenmeyer C.E., 2009, IntCal09 and Marine09 radiocarbon age calibration curves, 0 – 50,000 years cal BP: Radiocarbon, 51, 1111–1150. [ Links ]
Roy, P.D., Caballero, M., Lozano, R., Ortega, B., Lozano, S., Pi, T., Israde, I., Morton, O., 2010, Geochemical record of Late Quaternary paleoclimate from lacustrine sediments of paleo–lake San Felipe, western Sonora Desert, Mexico: Journal of South American Earth Sciences, 29, 586–596. [ Links ]
Roy, P.D., Caballero, M., Lozano, S., Morton, O., Lozano, R., Jonathan, M.P., Sánchez, J.L., Macías, M.C., 2012, Provenance of sediments deposited at paleolake San Felipe, western Sonora Desert: Implications to regimes of summer and winter precipitation during last 50 cal kyr BP: Journal of Arid Environments, 81, 47–58. [ Links ]
Schmidt, R.H.Jr., 1986, Chihuahuan climate, in Barlow, J.C., Powell, A.M., Timmerman, B.N. (eds.), Second Symposium on Resources of the Chihuahuan Desert Region U.S. and Mexico: Chihuahuan Desert Research Institute, 40–63. [ Links ]
Turpen, J.B., Angell, R.W., 1971, Aspects of molting and calcification in the ostracod Heterocypris: Biological Bulletin, 140, 331–338. [ Links ]
Urrutia–Fucugauchi, J., Ortega–Ramírez, J., Cruz–Gatica, R., 1997, Rock–magnetic study of Late Pleistocene–Holocene sediments from the Babícora lacustrine basin, Chihuahua, northern Mexico: Geofísica Internacional, 36, 77–86. [ Links ]
Van Morkhoven, F.P.C.M., 1963, Post–Paleozoic Ostracoda. Their morphology, taxonomy and economic use, Vol. II: New York, EIsevier Publishing Company, 478 pp. [ Links ]

