










Servicios Personalizados
Revista
Articulo
 Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista mexicana de ciencias geológicas
versión On-line ISSN 2007-2902versión impresa ISSN 1026-8774
Rev. mex. cienc. geol vol.29 no.2 Ciudad de México ago. 2012
The trilobite Maurotarion megacephalum sp. nov. (Aulacopleuridae) in the Lower Devonian of Argentina: phylogenetic and paleobiogeographic remarks
El trilobite Maurotarion megacephalum sp. nov. (Aulacopleuridae) del Devónico Inferior de Argentina: comentarios sobre filogenética y paleobiogeografía
Juan José Rustán and N. Emilio Vaccari
Universidad Nacional de La Rioja, M. de la Fuente s/n, CP 5300, La Rioja, Argentina, and Centro de Investigaciones en Ciencias de la Tierra (Universidad Nacional de Córdoba–CONICET), Av. Vélez Sarsfield 299, CP 5000, Córdoba, Argentina. *juanjorustan@gmail.com, jjrustan@conicet.gov
Manuscript received: July 11, 2011.
Corrected manuscript received: September 26, 2011.
Manuscript accepted: September 29, 2011.
ABSTRACT
Maurotarion megacephalum sp. nov., from the Pragian (Lower Devonian) of the Talacasto Formation, Precordillera basin, central–west Argentina, is described and interpreted as the first Malvinokaffric species of the genus outside the endemic subgenus M. (Malvinotarion). Based mainly on librigenal characters, the coevalM. periergum, from the Wenban Limestone, Nevada, U. S. A., is considered the closest allied taxon. Both species are interpreted as those most closely related to M. (Malvinotarion). Evidence suggests faunal exchange between low and high paleolatitudes for the Pragian, a hypothesis previously discarded in light of low sea level conditions interpreted during this time, and linked to a pattern of vicariance and radiation previously recognized from Malvinokaffric Maurotarion species.
Key words: Devonian, phylogeny, paleobiogeography, trilobites, Talacasto, Argentina.
RESUMEN
Maurotarion megacephalum sp. nov., del Pragiano (Devónico Inferior) de la Formación Talacasto, cuenca de Precordillera, centro–oeste de Argentina, es descrito e interpretado como la primera especie malvinocáfrica del género, no perteneciente al subgénero endémico M. (Malvinotarion). Con base en caracteres librigenales, principalmente, M. periergum, de la Caliza Wenban, Nevada, E.U.A., de edad similar, es considerado el taxón más cercanamente relacionado. Ambas especies son interpretadas como las más cercanas al subgénero M. (Malvinotarion). Las evidencias sugieren intercambio faunístico entre paleolatitudes bajas y altas para el Pragiano, una hipótesis previamente descartada en atención a las condiciones de nivel bajo del mar interpretadas para este intervalo, y ligadas a patrones de vicarianza y radiación con base en datos de Maurotarion malvinocáfricos.
Palabras clave: Devónico, filogenia, paleobiogeografa, trilobites, Talacasto, Argentina.
INTRODUCTION
The origin of the Malvinokaffric faunas has been discussed in light of a complex history of contrasting patterns involving migrations from low paleolatitudes and/or concomitant radiations of ancestral groups already present during the Silurian (Eldredge and Ormiston, 1979). Postulated extrinsic controls driving these processes include complex paleogeographical settings (Abe and Lieberman, 2009), latitudinal position (Boucot et al., 1969), oceanographic factors such as paleocurrents (Eldredge and Ormiston, 1979; Isaacson, 2007) and variations of sea level (Rustán and Vaccari, 2010).
Some Malvinokaffric trilobite groups exhibiting paradigmatic radiations, like the calmoniids, have shown to be strongly endemic (Eldredge and Branisa, 1980; Abe and Lieberman, 2009), whereas others such as dalmanitids appear to fit better a dispersal model (Carvalho and Fonseca, 2007; Holloway and Carvalho, 2009, 2010).
In turn, aulacopleurids, once interpreted as immigrants (Adrain and Edgecombe, 1996), have been demonstrated to have deep Silurian origins in cosmopolitan stocks (Rustán and Vaccari, 2010).
Our work is a contribution to these discussions, provided by new evidence on Maurotarion megacephalum sp. nov., from the Talacasto Formation, Argentina, and focused on phylogenetic and paleobiogeografic aspects of Malvinokaffric congenerics.
GEOLOGICAL SETTING
The Talacasto Formation (Lower Lochkovian–Emsian) is widespread and very well exposed in the Central Argentine Precordillera, in San Juan Province, central–west Argentina (Figure 1). It is composed of a marine sequence of intensely bioturbated greenish–grey mudstones with intercalated beds of sandstone. A composite stratigraphic column typically displays dark muddy levels in the lower part grading upwards to sandy levels, and in the uppermost part contains fine–grained levels with fossiliferous nodules (Figure 2). The Talacasto Formation reaches a maximum thickness of 1145 m in the Loma de los Piojos section. According to Astini (1991), this unit corresponds to a muddy shelf depositional system developed during a high–stand. It overlies the mainly Silurian shelf system of Los Espejos Formation. Strata of the deltaic system (Bustos and Astini, 1997) represented by the upper Lower–lower Upper Devonian Punta Negra Formation overlies the Talacasto Formation.
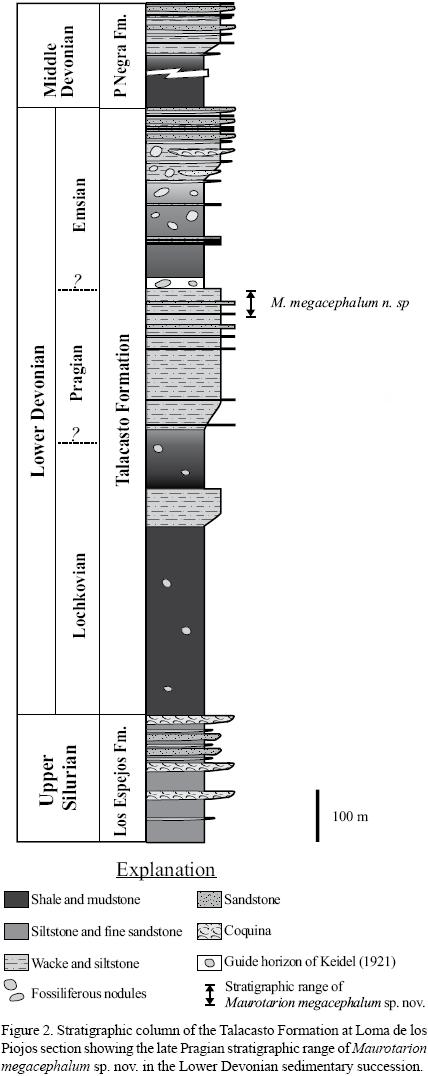
The uppermost levels of the Talacasto Formation at its type locality (Talacasto creek) were referred to the lowermost Emsian (Herrera, 1993, 1995), or latest Pragian (Herrera and Bustos, 2001, p. 369). This stratigraphic position corresponds to a distinctive horizon (Figure 2) referred as a stratigraphic marker by Keidel (1921) and Astini (1991), composed of 10 m thick, ocher laminated mudstone with grey carbonate fossiliferous nodules, which is interpreted as a condensed stratigraphic interval that is laterally continuous along the basin, so that it allows reliable lateral correlation. The strata overlying it, which are mainly recorded in the thicker northernmost sections of the basin and composed of dark greenish nodular mudstones with brownish sandstone levels, are considered early Emsian in age, or younger.
A more complete treatment of the geological aspects of the study area and stratigraphic units, can be found in previous works (Astini, 1991, Edgecombe et al., 1994, Bustos and Astini, 1997, Rustán and Vaccari, 2010).
MATERIAL
Provenance. Study material derives from the upper–middle part of the Talacasto Formation at Loma de los Piojos (Figures 1 and 2), a classical section located 6–7 km, southwest of Jáchal city, San Juan Province.
Trilobites are most abundantly preserved in a 50 m thick greenish–grey, intensely bioturbated interval located immediately below the marker horizon of Keidel (Figure 2). Fossils are preserved either as molds or exhibiting exo–skeleton, usually constituted by silicified, disarticulated sclerites, frequently exhibiting angularity both in respect to each other and to the bedding planes. They are, exceptionally found complete and articulated. The most common trilobites present in the same beds include Burmeisteria notica (Clarke, 1913), B. herschelii (Murchison, 1839), Dalmanitoides boehmi (Knod, 1908), D. drevermanni (Delo, 1935), Tarijactinoides jarcasensis (Suárez–Soruco, 1971), Bainella Rennie, 1930, and indeterminated calmoniids, phacopids and aulacopleurids. The remaining accompanying fauna is composed of brachiopods, bivalves, gastropods, hyolithids, conularids, crinoids, corals and orthoconic nautiloids.
Age. The stratigraphic position of the study seccion is strictly equivalent to the uppermost levels of the Talacasto Formation in its type section of the Talacasto river, and is thus considered latest Pragian in age (Herrera and Bustos, 2001). This age is supported by brachiopod (Herrera, 1993, 1995), and palinomorph (Le Hérissé et al., 1996) data. The presence of the trilobite Tarijactinoides jarcasensis, known only from the lower beds of the Gamoneda Formation bearing Scaphiocoelia in the Tarija area, southern Subandean Ranges from Bolivia, dated as Pragian on the basis of palinomorphs (Grahn, 2005), reinforces the chronological accuracy given for the M. megacephalum bearing strata.
SYSTEMATIC PALEONTOLOGY
Superfamily Aulacopleuroidea Angelin, 1854
Family Aulacopleuridae Angelin, 1854
Subfamily Otarioninae Richter and Richter, 1926
Genus Maurotarion Alberti, 1969
Type species. Harpidella maura Alberti, 1967.
Maurotarion megacephalum sp. nov.
Repository. Illustrated specimens are housed in the paleontological repository of the Centro de Investigaciones Paleobiológicas (CIPAL) numbered with the prefix CEGH–UNC, and Museo de Paleontología numbered with the prefix CORD–PZ, both at Universidad Nacional de Córdoba, Córdoba, Argentina.
The holotype CEGH–UNC 24864 (Fig 3, 1–6) is represented by a nearly articulated specimen, preserved in greenish–grey sandstone, composed of cranidium, left librigena, thorax and pygidium. The paratypes include (Figure 4), CEGH–UNC 24865–24868, 24870, 24882, 24885 Other assigned specimens CORD–PZ 8591, 8598, CEGH–UNC 7082, 7083, 24871–24876, 24878, 24880, 24881, 2488324884. All specimens come from the locality of Loma de Los Piojos (30°17'41.08"S Lat.; 68°46'40.26"W Long.).
Etymology. From latin mega (large), cephalum (head), due to the extremely globose glabella.
Diagnosis. Maurotarion with anterior margin evenly curved; exceedingly vaulted and globose, broad (tr.) and tall glabella; preglabellar field with a more convex small sagittal area, posteriorly located in front of preglabellar furrow, distinguishable in frontal view; deep, rounded apodemal fossula at junction with preglabellar and axial furrows; strong eye ridges, posterolaterally directed; lateral border exceedingly broad (tr.) and strongly inflated; very deep and strongly impressed lateral border furrow; very narrow librigenal field; slot–like apodemal fossula on librigenal posterior furrow near genal angle; genal spine strongly inflated and subcylindrical in section, with broad and strongly inflated base; sets of anastomosed to subparallels sculptural lines on lateral border and genal spine; long (sag., exsag.) occipital ring bearing a median short and conical, dorsal spine.
Description. Cranidium with anterior margin strong and evenly curved; anterior border thick, inflated, only subtly longer sagittally than exsagittally, directed forward and slightly upward; anterior border furrow long (sag.) and deep, shallowing sagittally, with subtle definition of an inverted "V" shape; short, preglabellar field, more than anterior border sagittally, with abrupt slope forward, with insinuated more convex, small sagittal area, posteriorly in front of preglabellar furrow; glabella broad (tr.), rounded, exceedingly inflated and vaulted, with maximum width across L1, 90 percent of length (sag., including occipital ring), with strong dorsal convexity, glabellar median lobe with nearly poligonal outline in dorsal view; preglabellar furrow convex forward, shallow and narrow exsagitally, deepest sagittally; axial furrows convexes but somewhat sinuous, with posterior section quite deep and approximately straight in front of L1, then shallowing and curved outward laterally in front of L2, becoming nearly straight and convergent anteriorly, exhibiting an antero–lateral constriction in front of S2 at junction with eye ridges, with a deep, rounded apodemal fossula associated at junction with preglabellar furrow; LO (occipital ring) long (sag., exsag.), more than anterior border, shortening slightly behind L1, taller than posterior part of glabella in sagittal profile, convex at the top, with a brief but stout occipital spine posteriorly located; L1 medium–size, width 20 percent that of glabella, length (exsag.) barely more than 30 percent that of glabella (sag., including occipital ring), contained dorsal and laterally inside of glabellar outline (not outstanding), subrounded anteriorly; L2 large, extended laterally but no more than L1, bending outward the lateral furrow, L3 not clearly visible; SO moderately short (sag., exsag.) and shallow, shorter and deeper behind L1; S1 strongly defined, evenly curved, inflecting abruptly outward anteriorly, deeper and broader anteriorly than posteriorly; S2 well defined (for the genus); S3 very faint; eye ridge, strong, directed posterolaterally reaching the posterior part of the glabellar constriction just anterior to S2; interocular fixigena moderately narrow and inclined; palpebral lobe short (exsag.) and reniform, of slightly inclined surface; palpebral furrow weakly developed; posterior border furrow slighty curved, posterolaterally directed, moderately long (exsag.) and shallow, narrower and shallower at junction of axial furrows, exhibiting a deep slot–like depression distally; posterior border inflated, with narrow (tr.) part proximal to fulcrum, then distally more curved, broader and longer (exsag.). Anterior branch of facial suture, evenly convex toward the sagittal plane becoming rapidly concave across (exsag.) the anterior border; posterior branch of the facial suture poorly known. Librigenal field narrow, gentle and evenly sloped, with delicate caecal sculpture; eye poorly known; margin of librigena gently curved in its anterior section, then becoming nearly straight posteriorly; lateral border very broad (tr.), more than librigenal field, thick and strongly inflated, gently convex dorsally, broadening posteriorly, with anastomosed to subparalell sculptural lines; lateral border furrow strongly impressed, very deep and broad (tr.), somewhat shallower anteriorly becoming deeper posteriorly, shallowing to disappearing in front of genal angle; librigenal posterior border furrow long (exsag.) and shallow, exhibiting a slot–like apodemal fossula near junction of lateral border, then continuing as a furrow along dorsal aspect of genal spine; librigenal posterior border thick and long (exsag.); genal spine with broad and strongly inflated base, long, approximately straight, with subcylindrical transversal section, inflated and moderately narrow (tr.), incorporating the broad and inflated parts of lateral and posterior borders, exhibiting anastomosed sculptural lines running sub–parallel on dorsal and lateral surface, the tip could not be observed; doublure poorly known, considerably narrow anteriorly in appearance, broadening laterally and backward, with an evident set of terrace lines running sub–parallel to anterior and lateral margins.
Rostral plate and hypostome, unknown.
Thorax only known from the holotype, incompletely preserved, with nearly smooth dorsal surface, showing at least eleven or twelve segments; pleural lobe slighty narrower (tr.) than axial lobe (measured in the most anterior and best preserved segment near the posterior–middle position); segments moderately long (sag., exsag.); the presence of median axial tubercles or spines cannot be observed due to broken dorsal aspect of axial rings; fulcrum apparently located in the middle of pleural lobe at least in the posteriormost preserved segments, axial ring with length (sag.) around 25 percent of width (tr.); axial furrow shallow and broad (tr.); pleural furrow, adaxially barely in contact with axial furrow, deepening and broadening abaxially to run postero–laterally as a narrow and deep nearly straight line laterally beyond fulcrum; posterior pleural band about more than twice as long (exsag.) as anterior band, adaxially; sharp triangular articulating boss developed on leading edge of anterior pleural band at fulcrum; articulating facet not adequately preserved; apodeme at posterior edge of each segment at junction of axial furrow.
Pygidium broad (tr.) and relatively short (sag.), of entire margin; medium–sized, with minimum length (sag. excluding articulating half ring) about 60 percent maximum width (tr.); broad axis with anterior width about 55 percent of total pygidial width; axial furrows moderately broad (tr.) and deep, evenly converging posteriorly and meeting postero–medially before reaching the margin, leaving a narrow posterior field, ending rapidly in a slope in sagittal profile; articulating half ring and first two axial rings prominent, third faint and posterior rings not well developed; up to three pleurae distinguishable, pleural and interpleural furrows of segment one to three short and shallow, incised, curved backward, becoming increasingly less distinct backward, without reaching the lateral pygidial margin; very short (tr.) pygidial border developed.
DISCUSSION
We refer this species to Maurotarion Alberti, 1969 based on of the following synapomorphies identified by Adrain and Chatterton (1995): anterior border longer medially than laterally; anterior border furrow of inverted "V" shape; palpebral lobes extended laterally past maximum divergence of anterior sections of facial sutures; librigenal field narrow relative to length; and lateral border inflated and broader (tr.) than posterior border.
Although the conspicuous librigenal lateral border furrow differs from the shallow one described in the diagnosis of Maurotarion by Adrain and Chatterton, we consider that the remaining observed characters reasonably support the generic assignment made herein.
Among Malvinokaffric congenerics, M. megacephalum sp. nov. more closely resembles M. (Malvinotarion) racheboeufi Adrain and Edgecombe, 1996, from the Emsian of the Icla Formation, Subandean Ranges of Bolivia, because of the exceedingly vaulted and inflated glabella. Other shared characters include the only weakly defined inverted "V" contour of the anterior border furrow, distinctive antero–lateral glabellar constrictions and apodemal fossulae on axial furrows at junction with eye ridges (see Adrain and Edgecombe, 1996; pl. 54 fig. 3 and 6). Maurotarion megacephalum differs fromM. (Malvinotarion) racheboeufi mainly in having its anterior border evenly curved, thicker and only scarcely lengthened sagitally and a prominent spine–like node on the occipital ring. The most evident differences lie in the librigena of M. megacephalum, which is characterized by a very strong inflation of the lateral and posterior borders, very thick genal spine ornamented with anastomosed subparallel lines, a genal spine base which is much more inflated and moderately narrower, only slightly curved and subcylindrical in section, and a lateral border furrow which is strongly impressed and running along the dorsal aspect of the genal spine.
In spite of the fact that several cranidial characters relate both species, the lack of a flattened and very broad (tr.) genal spine and genal spine base, the very inflated lateral border and the strongly pronounced lateral border furrow of M. megacephalum, do not support an assignment to M. (Malvinotarion). This Silurian–Devonian Malvinokaffric endemic subgenus was recognized by Rustán and Vaccari (2010) on the basis of a flat and very broad (tr.) lateral border and genal spine and a very broad (tr.) genal spine base, among other diagnostic characters. In addtition, all species of M. (Malvinotarion), including representatives from Argentina, Bolivia and South Africa, exhibit an only weakly impressed lateral border furrow.
The distinctive librigenal morphology of M. megacephalum most closely resembles that of M. periergum (Haas, 1969), from Pragian of the Wenban Limestone, Cortez Range, central Nevada, U.S.A.. Shared characters supporting a close relationship (based in the original work of Haas and additional data by Adrain, 2009), include the thick lateral border, broad (tr.) and deep, lateral border furrow, broadened genal spine base –yet not so broad as in M. (Malvinotarion)–, the librigenal lateral border furrow shallowest in front of genal angle, a thick genal spine sub–cylindrical in cross section, and a distally deepest librigenal posterior border furrow which continues along the dorsal aspect of the genal spine as a delicate furrow. Characters such as the granulation, tuberculation, length (sag.) of the anterior border, curved genal spine, less inclined preglabellar furrow, more depressed glabella and longer (sag., exsag) and narrower (tr.) pygidium with more curved pygidial pleural ribs in M. periergum, separate both species.
In general terms, similar comparisons could also be extended to M. chrysion Adrain, 2009, which seems to be the most closely non Malvinokaffric Maurotarion species related to M. periergum and M. megacephalum. In turn, these species resemble M. wenbanense Adrain, 2009, M. fooi Adrain, 2009, from the same formation and age in Nevada, and M. axitiosum (Campbell, 1977) from the Lochkhovian Haragan Formation, Oklahoma, which were discussed in detail by Adrain (2009).
Mainly due to its librigenal characters, M. periergum was proposed and discussed at length as sister taxon of the Malvinokaffric Maurotarion clade (Adrain and Edgecombe, 1996; Adrain, 2009), and consequently the origins of this group were explained by migration from north to south during Early Devonian times (Adrain and Edgecombe, 1996).
However, since Maurotarion species were confirmed to be present in South America by early Silurian (Rustán and Vaccari, 2010), as previously suggested (Edgecombe and Fortey, 2000; Adrain, 2009), no Devonian migrations need to be invoked to explain the distribution. Consequently the Malvinotarion–like librigenal characters of M. periergum were interpreted by Rustán and Vaccari (2010) as probably convergent.
Furthermore, a radiation of M. (Malvinotarion) species was proposed during the Devonian (Rustán and Vaccari, 2010), which peaked during the Emsian according a highest percentage (about 60 %) of the first stratigraphic appearances recorded in this interval. This phylogenetic pattern was linked to falling sea level encouraging vicariance and concomitant speciation, particularly during the Pragian. In the context of this low sea level hindering dispersals, previous interpretations of Maurotarion periergum as the sister taxon of the Devonian Maurotarion clade (implying migrations), resulted questioned as well.
However, new evidence treated herein indicate that similarities in librigenal characters support a probable sister taxon relationship between M. periergum and M. megacephalum rather than convergence, and that both species are the most closely related to the Malvinokaffric subgenus M. (Malvinotarion).
This interpretation resurrects the idea of faunal exchange during Pragian. Furthermore it suggests that the largely rejected hypothesis of dispersals from south to north during Early Devonian (Adrain and Edgecombe, 1996; Rustán and Vaccari, 2010; Holloway and Carvalho, 2010) should not be rejected.
Compelling evidence based on records of the phacopid trilobite Reedops cf. bronni from the northernmost reaches of the Talacasto Formation in Precordillera basin, at Sierra de las Minitas, La Rioja province (Rustán et al., 2011), reinforces this hypothesis. Reedops is almost unknown to range below the Pragian elsewhere in the world, and the type species R. bronni, has been considered a Pragian index fossil in the Czech Republic (Chlupác, 1977). The occurrence of this taxon constitutes an unusual paleobiogeographical indicator of affinities with Bohemian area of the Old World Realm, and migrations have to be invoked to explain its presence in the Pragian of the Talacasto Formation.
This conclusion strongly contrasts with previous proposals suggesting low sea levels during the Pragian encouraging vicariance and speciation and restricting dispersals. Thus, evidence as a whole, accounts for a rich and complex phylo–geographical history in the development of the Malvinokaffric trilobite faunas.
ACKNOWLEDGEMENTS
We thank D. Balseiro, R. Foglia, and A. Rydczewski who have collaborated invaluably in the field work. We also thank B. Waisfeld who has collected some materials in her geology degree work and placed this at our disposal. The reviewers J. M. Adrain and D.L. Bruton greatly improved this paper. Work have been possible thanks upon contributing of CONICET and ANPCyT (Agencia Nacional de Promoción Científica y Tecnológica) PICT 2006–01272 granted to Claudia Rubinstein.
REFERENCES
Abe, F.R., Lieberman, B.S., 2009, The nature of evolutionary radiations: a case study involving Devonian trilobites: Evolutionary Biology, 36, 225–234. [ Links ]
Adrain, J.M., 2009, New and revised species of the aulacopleurid trilobite Maurotarion from the Lower Devonian (Pragian) of Nevada: Zootaxa, 2215, 1–23. [ Links ]
Adrain, J.M., Chatterton, B.D.E., 1995, The otarionine trilobites Harpidella and Maurotarion, with species from northwestern Canada, the United States, and Australia: Journal of Paleontology, 69(4), 307–326. [ Links ]
Adrain, J.M., Edgecombe, G.D., 1996, Devonian Aulacopleurid trilobites of the Malvinokaffric Realm: Geobios, 29(4), 417–436. [ Links ]
Alberti,G.K.B., 1967, Neue obersilurische sowie unter–und mitteldevonische Trilobiten aus marokko, Deutschland und einigen anderen europäischen Gebieten. 2: Senckenbergiana lethaea, 48, 481–509. [ Links ]
Alberti, G.K.B., 1969, Trilobiten des jüngeren Siluriums sowie des unter–und Mitteldevons. I: Abhandlungen der Senckenbergischen Naturforschenden Gesellschaft, 520, 521–692. [ Links ]
Angelin, N., 1854, Palaeontologia Scandinavica, Part I, Crustacea Formationis Transitionis: Academiae Regiae Scientaum Suecanae. Holmiae. fasc. 2., 21–92. [ Links ]
Astini, A.R., 1991, Sedimentología de la Formación Talacasto: plataforma fangosa del Devónico precordillerano, provincia de San Juan: Revista de la Asociación Geológica Argentina, 44, 277–294. [ Links ]
Boucot, A.J., Johnson, J.G., Talent, J.A., 1969, Early Devonian brachiopods zoogeography: Geological Society American Special Paper, 119, 1–113. [ Links ]
Bustos, U.D., Astini, A.R., 1997, Formación Punta Negra: análisis secuencial y evolución de la Cuenca Devónica Precordillerana: Revista de la Asociación Argentina de Sedimentología 4, 97–111. [ Links ]
Campbell, K.S.W., 1977, Trilobites of the Haragan, Bois d'Arc and Frisco Formations (Early Devonian) Arbuckle Mountains Region, Oklahoma: Oklahoma Geological Survey Bulletin, 123, 1–227. [ Links ]
Carvalho, M.D.G.P.D., Fonseca, V.M.M.D., 2007, The trilobite 'Dalmanites 'maecurua Clarke, 1890 (Middle Devonian, Amazon Basin, Brazil) and the new genus Amazonaspis (Synphoriidae): American Museum Novitates, 3591, 1–14. [ Links ]
Clarke, J.M., 1913, Fosseis devonianos do Paraná: Monografias Serviço Geologico e Mineralogico do Brasil 1: 1–353. [ Links ]
Chlupác, I., 1977, The phacopid trilobites of the Silurian and Devonian of Czechoslovakia: Rozpravy Ústredního Ústavu Geologického, 43, 1–172. [ Links ]
Delo, D.M., 1935, A revision of the phacopid trilobites: Journal of Paleontology, 9, 402–420. [ Links ]
Edgecombe, G.D., Fortey, R.A., 2000, Silurian trilobites from the El Carmen Formation, Bolivia: Senckenbergiana Lethaea, 79(2), 329–355. [ Links ]
Edgecombe, G.D., Vaccari, N.E., Waisfeld, B.G., 1994, Lower Devonian calmoniid trilobites from the Bouleia Group, and remarks on the tempo of calmoniid radiation: Geological Magazine, 131(4), 449–464. [ Links ]
Eldredge, N., Branisa, L., 1980, Calmoniid trilobites of the Lower Devonian Scaphiocoelia Zone of Bolivia, with remarks on related species: Bulletin American Museum of Natural History, 165(2), 183–289. [ Links ]
Eldredge, N., Ormiston, L., 1979, Biogeography of Silurian and Devonian Trilobites of the Malvinokaffric Realm, in Gray J., Boucot H.J. (eds.), Historical Biogeography, Plate Tectonics, and the changing Environment: The Oregon Satate University Press, 147–167. [ Links ]
Grahn, Y., 2005, Devonian chitinozoan biozones of Western Gondwana: Acta Geologica Polonica, 55, 211–227. [ Links ]
Haas, W., 1969, Lower Devonian trilobites from central Nevada and northern Mexico: Journal of Paleontology, 43, 641–659. [ Links ]
Herrera, Z.A. 1993. Nuevas precisiones sobre la edad de la Formación Talacasto (Precordillera Argentina) en base a su fauna de braquiópodos: XII Congreso Geológico Argentino y II Congreso de Exploración de Hidrocarburos, Buenos Aires, 289–295. [ Links ]
Herrera, Z.A., 1995, The Lower Devonian chonetoidean brachiopods from the Argentine Precordillera, in Racheboeuf, P., (ed.), Four contributions to the study of chonetoidean brachiopods: Centre des Sciences de la Terre, Université Claude–Bernard–Lyon I, 136. 101–147. [ Links ]
Herrera, Z.A., Bustos, U.D., 2001, Braquiópodos devónicos de la Formación Punta Negra, en el perfil del Río de las Chacritas, Precordillera Argentina: Ameghiniana 38(4), 367–374. [ Links ]
Holloway, D., Carvalho, M.G.P.D. 2009, The extraordinary trilobite Fenestraspis (Dalmanitidae, Synphoriinae) from the Lower Devonian of Bolivia: Palaeontology, 52(4), 933–949. [ Links ]
Holloway, D.J., Carvalho, M.G.P.D. 2010, The trilobite Chacomurus (Dalmanitidae, Synphoriinae) from the Lower Devonian of Bolivia: Memoirs of the Association of Australasian Palaeontologists, 39, 71–83. [ Links ]
Isaacson, P.E. 2007. Mid–paleozoic biogeography of the central andes: Endemic faunas, immigrants, and paleogeography, in Díaz–Martínez, E., Rábano, I., (eds.), 4th European Meeting on the Palaeontology and Stratigraphy of Latin America: Cuadernos del Museo Geominero, 8, 213–218. [ Links ]
Keidel, J., 1921, Observaciones geológicas en la Precordillera de San Juan y Mendoza: Anales del Ministerio de Agricultura, Sección Geología, Mineralogía y Minería, 15, 7–102. [ Links ]
Knod, R., 1908, Devonische faunen boliviens, in Steinmann, G., (ed.), Beiträge zur Geologie und Paläontologie von Südamerika: Neues Jahrbuch für Mineralogie, Geologie und Paläontologie, 25, 493–600. [ Links ]
Le Hérissé , A., Rubinstein, C.R., Steemans, P., 1996, Lower Devonian palynomorphs from the Talacasto Formation, Cerro del Fuerte Section,San Juan Precordillera, Argentina, in Fatka, O., Servais, T. (eds.), Acritarcha in Praha: Acta Universitatis Carolinae Geologica, 40, 497–515. [ Links ]
Murchison, R.I.,1839, The Silurian System: London, J. Murray, 1–768. [ Links ]
Rennie, J.V.L., 1930, Some phacopidae from the Bokkeveld Series: Transactions of the Royal Society of South Africa, 18, 327–360. [ Links ]
Richter, R., Richter, E., 1926, Die Trilobiten des Oberdevons. Beiträge zur Kenntnis devonischer Trilobiten IV: Abhandlungen der Preussischen Geologischen Landesanstalt, neue Folge, 99, 1–314. [ Links ]
Rustán, J.J., Vaccari, N.E., 2010, The aulacopleurid trilobite Maurotarion Alberti, 1969 in the silurian–Devonian of Argentina:systematic, phylogenetic and paleobiogeographic significance: Journal of Paleontology, 84(6), 1082–1098. [ Links ]
Rustán, J.J., Vaccari, N.E., Astini, R.A., 2011, Early Devonian trilobites from the Sierra de las Minitas, northernmost Precordillera (La Rioja Province), Argentina: Ameghiniana 48(2), 226–241. [ Links ]
Suárez–Soruco, R.S., 1971, Tarijactinoides jarcasensis n. gen. n. sp del Devónico Inferior: Boletín del Servicio Geológico de Bolivia, 15, 53–56. [ Links ]

