










Services on Demand
Journal

Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
 Cited by SciELO
Cited by SciELO Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkRevista mexicana de ciencias geológicas
On-line version ISSN 2007-2902Print version ISSN 1026-8774
Rev. mex. cienc. geol vol.29 n.1 Ciudad de México Apr. 2012
Bioestratigrafía de conodontes del Darriwiliano medio (Ordovícico) de Argentina: la Formación Las Aguaditas, Precordillera Central
Biostratigraphy of Middle Darriwilian (Ordovician) conodonts from Argentina: Las Aguaditas Formation, Central Precordillera
Susana Heredia
CONICET – IM, Universidad Nacional de San Juan, Urquiza y Libertador (5400), San Juan, Argentina. sheredia@unsj.edu.ar
Manuscrito recibido: Junio 3, 2011
Manuscrito corregido recibido: Septiembre 14, 2011
Manuscrito aceptado: Septiembre 16, 2011
RESUMEN
Se recuperaron elementos del género Eoplacognathus de muestras de la Formación Las Aguaditas aflorante en el río de Las Chacritas, provincia de San Juan, Argentina. Estos depósitos de calizas oscuras son grainstones oscuros, packstones y wackestones y representan facies de rampa carbonática distal. Los conodontes guía Eoplacognathuspseudoplanus (Viira) y Eoplacognathus suecicus Bergström documentan las Zonas de E. pseudoplanus y de E. suecicus indicando una edad darriwiliana media para los niveles portadores de conodontes en la Formación Las Aguaditas. Se realiza una evaluación de los antecedentes sobre el registro fósil del género Eoplacognathus en la Precordillera Argentina. Por último, se propone el uso del esquema de biozonas de conodontes del Darriwiliano medio de Báltica para la Precordillera Argentina.
Palabras clave: Bioestratigrafía, Eoplacognathus, Darriwiliano, Precordillera Argentina.
ABSTRACT
Several P elements of Eoplacognathus were recovered from samples collected in Las Aguaditas Formation, Las Chacritas river section, San Juan Province, Argentina. These dark grainstones, packstones, and wackestones represent distal carbonate rampfacies. The key conodonts Eoplacognathus pseudoplanus (Viira) and Eoplacognathus suecicus Bergström, document the Eoplacognathus pseudoplanus and the E. suecicus Zones pointing out middle Darriwilian age for the conodont–bearing strata in Las Aguaditas Formation. The species E. suecicus is recorded here for first time in the Argentine Precordillera. An evaluation of the record of this middle Darriwilian conodont genus in the Precordillera is presented here. Finally, the use of the middle Darriwilian Baltic conodont chart is proposed for the Argentine Precordillera.
Key words: Biostratigraphy, Eoplacognathus, Darriwilian, Argentine Precordillera.
INTRODUCCIÓN
La Precordillera del Oeste de Argentina es parte de una región mayor denominada Cuyania (Ramos, 1995; Ramos et al., 1998), la cual es interpretada como alóctona y que no ocupó su presente posición hasta el Ordovícico Medio (Astini, 1998; Astini y Thomas, 1999) o hasta el Devónico (Aceñolaza et al., 2002; Finney, 2007). La Precordillera exhibe espesas sucesiones de rocas carbonáticas de edad cámbrica y ordovícica inferior y media, indicando estabilidad de la sedimentación en las diferentes áreas de la plataforma. Durante el Ordovícico Medio y Superior se observa un brusco cambio en la sedimentación con el desarrollo de espesas sucesiones clásticas por encima los carbonatos; estas facies siliciclásticas muestran una gran variabilidad y heterogeneidad lateral y han sido interpretadas como el registro de una convulsión tectónica y paleogeográfica en el Ordovícico (Astini, 2003).
En el ámbito occidental de la Precordillera Central, en la Sierra de La Trampa, afloran depósitos del Paleozoico Inferior que han sido estudiados en las últimas décadas por diversos especialistas (Espisúa, 1968; Astini, 1994; Peralta y Baldis, 1995; Carrera y Astini, 1998; Peralta et al., 1999a, 1999b ; Heredia et al., 2005 a, b) (Figura 1). De este sector se dieron a conocer depósitos carbonáticos de edad ordovícica cuya mejor exposición se encuentra en la Quebrada del río Las Chacritas. Los materiales de esta localidad fueron descriptos por primera vez por Espisúa (1968) como Formación San Juan, quien además les dio nombres formales a los miembros inferior (Miembro de Calizas Macizas), y superior (Miembro de Calizas Lajosas).
Por sus características litológicas y paleontológicas el miembro inferior ha sido homologado a la Formación San Juan por Peralta y Baldis (1995) y por Carrera y Astini (1998). En tanto que el Miembro de Calizas Lajosas ha recibido diferentes denominaciones formales e informales; Peralta y Baldis (1995) lo homologan con la Formación Las Aguaditas (en el sentido de Baldis et al., 1982) cuya localidad tipo es la Quebrada de las Aguaditas, señalando además la presencia de dos miembros en la Quebrada del río de Las Chacritas, uno inferior compuesto por bancos calcáreos delgados con estratofábrica tabular de edad "llanvirniana" (en el sentido de las Series Británicas) y un miembro superior, representado por paraconglomerados calcáreo y calcipelitas negras, que contiene graptolitos de la Zona de H. teretiusculus y trilobites asignados a Incaia deormaecheai Baldis y Cabaleri arrojando el conjunto una edad "Llandeiliana tardía". También, Astini (1994) reconoce dos litosomas para este "Miembro superior de la Formación San Juan" en el sentido de Espisúa (1968), el litosoma inferior de 65 metros de espesor promedio constituido por calizas lajosas, muy fosilíferas con indicios de somerización al tope, y el litosoma superior de espesor creciente hacia el norte y ausente hacia el sur, compuesto por calizas margosas negras con abundantes graptolitos, al que Astini (1998) refiere como Formación Las Chacritas y correlaciona con la Formación Las Aguaditas. Carrera (1997) denomina informalmente al Miembro superior de Espisúa en la Quebrada Las Tunas, como "Unidad Calcárea las Tunas". Los últimos aportes estratigráficos y bioestratigráficos sobre estos afloramientos corresponden a Peralta et al. (1999a, 1999b), quienes definen formalmente una nueva unidad que sería equivalente al Miembro de Calizas Lajosas de Espisúa (1968) como Formación Las Chacritas. La edad de esta unidad es darriwiliana en base al registro de tres biozonas y dos subzonas de conodontes: Lenodus variabilis, Subzona de Paroistodus horridus (Peralta et al., 1999 a, b), Eoplacognathus pseudoplanus/Dzikodus tablepointensis y Eoplacognathus suecicus, Subzona de Histiodella kristinae (Heredia et al., 2005a).
Luego de una revisión bibliográfica y de datos propios de campo de la Quebrada del río de Las Chacritas y de la localidad de Las Aguaditas, se referirá aquí a los afloramientos que sobreyacen a la Formación San Juan en el río de Las Chacritas como Formación Las Aguaditas (siguiendo el criterio de Peralta y Baldis, 1995), estando presentes en la localidad de estudio el Miembro Inferior y parte del Miembro Lajoso definidos en la localidad tipo por Baldis y Blasco (1974). Los datos propios de los afloramientos ordovícicos en el sector de estudio y una revisión de las antiguas colecciones de conodontes estudiadas por Heredia et al. (2005a), condujeron a advertir que la mayoría de las muestras colectadas y mencionadas en Peralta et al. (1999a, 1999b) y en Heredia et al. (2005a) pertenecen en realidad a los tramos más altos de la Formación San Juan y algunas a la Formación Las Aguaditas (LCH 10 a la 18).
Los conodontes recuperados de la Formación Las Aguaditas han aportado resultados que permiten dar una mayor precisión y puntualizar las edades de estos afloramientos. El hallazgo de las especies guía Eoplacognathus pseudoplanus (Viira) y Eoplacognathus suecicus Bergström acotan bioestratigráficamente estos afloramientos haciendo posible compararlos con otros de similares edades y litologías en el ámbito de la Precordillera Central. Además, se revisó la antigua colección de conodontes y se reinterpretaron los elementos señalados como Eoplacognathus? (INGEO MP 101/1) y Lenodus variabilis (Sergeeva) (INGEO MP 100/1) (Heredia et al., 2005a, fig. 3 A y B), y cuya procedencia coincide con parte del muestreo realizado en este trabajo, y que a diferencia de lo expresado en el perfil de Peralta et al. (1999a, 1999b) aparecen distanciadas a escasos 0.70 metros. Esta contribución es parte de los primeros resultados de un estudio más amplio cuyo objetivo es establecer un esquema bioestratigráfico basado en estudios de detalle de conodontes del Ordovícico Medio de Precordillera.
ESTRATIGRAFÍA
Formación Las Aguaditas
En la Quebrada del río Las Chacritas sobre los grainstones y packstones encriníticos bioclásticos del techo de la Formación San Juan, se dispone la Formación Las Aguaditas. Esta unidad, de unos 45 metros de espesor en afloramiento, está caracterizada por mudstones alternantes con pelitas negras, wackestones, packstones bioclásticos, brechas bioclásticas, calcarenitas gradadas y grainstones bioclásticos y calcilutitas y pelitas negras (Figura 2). Los fragmentos de trilobites y esponjas son abundantes, como así también braquiópodos bien conservados. Esta breve descripción resulta coincidente con la realizada por Astini (1995). Asimismo, es similar a la descripción de los dos miembros definidos por Peralta y Baldis (1995), donde el inferior está compuesto por bancos calcáreos delgados con estratofábrica tabular, mientras que el superior está representado por mudstones, pelitas y calcipelitas negras que contienen graptolitos y trilobites.

MATERIALES Y MÉTODOS
Se levantó un perfil estratigráfico en la Quebrada del río Las Chacritas (30°33'50.2''S; 68°51'51.6"O) y se colectaron muestras en los packstones bioclásticos (Lag 8), wackestones (Lag 7', 7, 6, 5, 4 y 3), y grainstones bioclásticos (Lag 2, 1 y 0) con el fin de obtener conodontes. Las muestras fueron procesadas en laboratorio mediante los métodos convencionales de ácido fórmico (Stone, 1987). En los residuos obtenidos se rescataron mudas de trilobites, abundantes ostrácodos, conodontes, espículas de poríferos, braquiópodos inarticulados y artejos de crinoideos. El color que exhiben los conodontes corresponde a un valor de 3 del CAI (Color Alteration Index de Epstein et al., 1977). La conservación de los conodontes, en general, es muy buena; sin embargo, los procedentes de las muestras Lag 0 y Lag 1, aparecen rotos e incompletos, especialmente los ejemplares más robustos. Las superficies de los elementos se observan límpidas y sin evidencias de abrasión. Las microfotografías fueron obtenidas por microscopía electrónica de barrido en el Laboratorio de Microscopía Electrónica del CCT–Mendoza y en el Servicio de Microscopía de la Universidad Nacional del Comahue (Neuquén). Los ejemplares están depositados en la colección de Microvertebrados Lillo– Conodontes de la Facultad de Ciencias Naturales e Instituto Miguel Lillo, bajo las siglas CML–C, excepto dos de ellos que están en la colección del INGEO (Universidad Nacional de San Juan) bajo las siglas INGEO–MP.
CONODONTES
Hasta el presente se han recuperado unos 998 elementos de conodontes, entre ejemplares determinables y fragmentos, procedentes de 9 muestras fértiles de aproximadamente 1.8 kg cada una, además de los dos elementos mencionados en Heredia et al. (2005a) provenientes de las muestras LCH 12 y 14. El presente estudio registra el género Eoplacognathus, cuyas especies E. pseudo–planus (Viira) y E. suecicus Bergström son importantes indicadores bioestratigráficos. Algunas de las especies que acompañan a E. pseudoplanus son: "Bryantodiná" aff. typicalis (Stauffer), Ansella jemtlandica (Löfgren), Periodon aculeatus Hadding, Paroistodus (Barnes y Poplawski), Microzarkodina cf. ozarkodella Lindstrom, Baltoniodus medius Löfgren, Rossodus barnesi Albanesi, Protopanderodus gradatus Serpagli, Drepanoistodus basiovalis (Sergeeva), Parapaltodus simplissisimus Stouge, Costiconus ethingtoni Fâhraeus, Erraticodon balticus Dzik e Histiodella kristinae Stouge. Las especies que acompañan a E. suecicus son: Ansella jemtlandica, Baltoniodus sp., Dzikodus sp., Pygodus anitae Bergström, Histiodella sp., Paroistodus horridus, Periodon aculeatus, Protopanderodus sp., "Bryantodina" aff. typicalis.
Taxonomía
El aparato de Eoplacognathus fue interpretado por Bergström (1971), quien describió únicamente formas P (Pa y Pb derechos e izquierdos). Löfgren y Zhang (2003) reconocieron los elementos S (Sa,Sb, Sc y Sd) y el elemento M del aparato de Eoplacognathus pseudoplanus.
En este aporte sobre las especies de Eoplacognathus de la Formación Las Aguaditas se mencionan brevemente sus características principales y se ilustran, pues han sido descriptas y señaladas frecuentemente en la literatura especifica, no siendo necesario realizar una caracterización taxonómica detallada de ellas.
La lista sinonímica es condensada y contiene especialmente las descripciones y las citas de estudios realizados en la Precordillera correspondientes a los elementos reconsiderados como las especies en cuestión. En las descripciones se ha utilizado los términos convencionales de orientación (anterior, posterior y lateral) dando por sentado que éstos no guardan relación con la orientación anatómica de los elementos en el individuo (Purnell et al., 2000).
Orden Prioniodontida Dzik, 1976
Superfamilia Prioniodontacea Bassler, 1925
Familia Polyplacognathidae Bergström, 1971
Género Eoplacognathus Hamar, 1966
Especie tipo. Ambolodus lindstroemi Hamar, 1964
Eoplacognathus pseudoplanus (Viira, 1974)
Figura 3,1–3
1974, Ambaloduspseudoplanus Viira, pág. 54, figs. 43–46, lám. 6, figs 25, 29, 31.
Sinonimia completa en Löfgren y Zhang, 2003. Elementos P
?1985, Eoplacognathus pseudoplanus (Viira). Sarmiento, lám. 1, figs. 5–10.
?1991, Eoplacognathus suecicus Bergström. Sarmiento, pág.119, lám. 1, figs. 9–15.
1998, Lenodus variabilis Sergeeva Albanesi, págs. 159–160, lám. 5, figs. 18 y 19.
1995, Lenodus pseudoplanus (Viira). Lehnert, p. 94, lám. 13, figs. 8, 14.
2002, Lenodus variabilis Sergeeva. Albanesi y Ortega, lám. 1, fig. 11.
2005a, Eoplacognathus pseudoplanus (Viira). Heredia et al., lám. 3, fig. D.
2008, Lenodus variabilis (Sergeeva). Voldman et al. , lám. 4, fig. j.
Descripción: El aparato completo de E. pseudoplanus (en el sentido de Löfgren y Zhang, 2003) no ha sido recuperado de la sección estudiada y se cuenta únicamente con los elementos P, pero tal como afirma Zhang (1999) al analizar el aparato de E. suecicus son los elementos P, especialmente los Pa adultos y gerontes, los que resultan diagnósticos para identificar las diferentes especies de Eoplacognathus.
La mayoría de los elementos P son formas juveniles y adultas. Los elementos Pa derecho e izquierdo son similares (no idénticos) y aproximadamente especulares, con cuatro procesos: anterior, antero–lateral, posterior y postero–lateral.
Estos elementos presentan características propias de aquellos en transición hacia la especie siguiente (en los escasos individuos gerontes se observa cierta similitud con formas muy tempranas de E. suecicus) y permiten compararlos con los figurados para el Sur de China (Zhang, 1998a). A modo de ejemplo se destaca el elemento Pa donde el proceso posterior se curva (Figura 3, 2) configurando una hilera de dentículos sinuosa sobre la plataforma, al igual que los ejemplares que exhibe Mestre (2010) provenientes del techo de la Formación San Juan que aparece infrayaciendo a la Formación Las Aguaditas.
Los elementos Pb tienen 3 procesos, el proceso anterior y el antero–lateral presentan plataformas estrechas y el proceso posterior tiene una plataforma más amplia en los elementos Pb derecho. No se han recuperado elementos S y M.
Material. Muestra Lag 8, 1 Pa izquierdo, 3 Pa derechos, 3 Pb izquierdos, 2 Pb derechos y 3 fragmentos indeterminables.
Repositorio. CML–C 3400/1–12.
Distribución. El material aquí descrito proviene de la sección del río Las Chacritas, muestra Lag 8 (Figura 2).
Discusión: El uso de E. pseudoplanus como conodonte guía para la parte alta del piso Kunda fue propuesto por primera vez por Viira (1974). Löfgren (1978) incluyó elementos tardíos de E. pseudoplanus ("E. suecicus'") para definir la Subzona de conjunto de Eoplacognathus suecicus–Panderodus sulcatus. Recientemente, Löfgren (2000) menciona que el registro más antiguo de E. pseudoplanus coincide con la última aparición de Lenodus variabilis y Yangtzeplacognathus crassus, y con la primera aparición de Dapsilodus viruensis (Fâhrasus). Zhang (1998b) estableció para la Zona de E. pseudoplanus dos subzonas:
Microzarkodina hagetiana y Microzarkodina ozarkodella; finalmente, la Zona de E. pseudoplanus se caracteriza porque en su parte inferior aparece Dzikodus tablepointensis (Stouge) reemplazando a D. hunanensis Zhang.
Löfgren y Zhang (2003) propusieron un aparato septimenbrado para E. pseudoplanus, y analizaron el linaje de esta especie, además Löfgren (2004) describió la conodontofauna de la Zona de E. pseudoplanus en Baltoescandia.
El reconocimiento de la Zona de E. pseudoplanus/D. tablepointensis, basado en elementos de las especies nominales, fue realizado por Heredia et al. (2005a) en el ámbito de la Precordillera Argentina. La distribución de la especie E. pseudoplanus está restringida al centro–sur de China, Baltoescandia, Newfoundland (con dudas) (Stouge, 1984) y Precordillera argentina. Si bien los conodontes de esta última región que se atribuyen a la Zona de E. pseudoplanus coexisten con un reducido número de taxones que son característicos de la Provincia del Midcontinent, se debe señalar que la bioestratigrafía está basada en la presencia de especies de afinidad báltica cuya filogenia es bien conocida (Bergström, 1983). Los ejemplares del techo de la Formación San Juan en la Sierra de Villicúm que fueron atribuidos por Sarmiento (1985) en primera instancia a E. pseudoplanus y luego (Sarmiento, 1991) a E. suecicus se asignan con dudas a E. pseudoplanus (ver sinonimia) porque exhiben una mayor afinidad morfológica con esta especie y no con E. suecicus.
Eoplacognathus suecicus Bergström, 1971
Figura 4, 1–6
Elementos P
Descripción. Se han rescatado 49 elementos Pa y Pb, entre juveniles, adultos y gerontes. El Pb izquierdo tiene un proceso anterior curvado cuya longitud varía entre los individuos gerontes y juveniles. El Pb derecho tiene tres procesos aproximadamente de la misma longitud. Los elementos Pa, tanto izquierdos como derechos son aproximadamente similares, con una expansión sobre el lado interno y un corto y ancho proceso postero lateral. Todos los procesos llevan una hilera de dentículos. Se observan hileras radiales de tubérculos en los procesos postero laterales (Figura 4, 1 y 2) similares a las que presentan los elementos de Dzikodus y Pygodus. Superficialmente, sobre las plataformas, se observa la típica estructura poligonal que caracteriza tanto a este grupo como a Dzikodus y Pygodus. Morfológicamente todos los ejemplares estudiados y figurados representan formas tardías de E. suecicus por comparación con los elementos figurados por Zhang (1999) que provienen de niveles atribuidos a la Subzona de P. anitae.
Material. Muestras Lag 7' (LCH 12), Lag 7 (LCH 14), Lag 5, Lag 1, Lag 0. Elementos recuperados: 5 Pa izquierdo, 3 Pa derecho, 6 Pb izquierdo, 2 Pb derecho y 35 fragmentos. Repositorio: Lag 7' /INGEO MP 101/1; Lag 7/CML–C 3401(1–20) e INGEO MP 100/1; Lag 5/CML–C 3402(1), Lag 1 y 0/CML–C 3403 (1–28).
Distribución. El material aquí descripto proviene de la sección del río Las Chacritas, muestras Lag 7' a Lag 0 (Figura 2); se incluyen también los ejemplares obtenidos de las muestras LCH 12 y 14, como ya se mencionó.
Discusión. Eoplacognathus suecicus fue nominado, descripto y utilizado como conodonte guía para definir una subzona por Bergström (1971). Este autor consideró que E. suecicus poseía un aparato con dos tipos de elementos pastiniplanados no especulares y dos tipos de elementos esteliplanados no especulares; estos elementos fueron descriptos por Bergström (1971) excepto el elemento esteliplanado derecho. Zhang (1999) considera que las descripciones originales de Bergström (1971) no permiten distinguir a E. suecicus de E. pseudoplanus o de Lenodus variabilis (Sergeeva), y propone utilizar los elementos esteliplanate para la diagnosis de la especie.
Los elementos que se atribuyen en este trabajo a E. suecicus exhiben una variación morfológica importante según se trate de individuos juveniles, adultos o gerontes, que se traduce en el desarrollo y ensanchamiento de los procesos posteriores en los elementos Pb (pastiniplanado) y con la aparición y desarrollo de lóbulos en los elementos Pa (esteliplanado). Al igual que otras especies del género Eoplacognathus, E. suecicus muestra una gran variabilidad morfológica desde las formas tempranas a las tardías, las que fueron ejemplificadas por Zhang (1999). Estas formas fueron comparadas con los ejemplares de la Formación Las Aguaditas, lo que ha llevado a concluir que estos elementos corresponden a formas tardías.
Lehnert (1995) figuró ejemplares atribuidos a Eoplacognathus suecicus que provienen del techo de la Formación San Juan. Al analizar el material figurado por este autor se puede apreciar que se trata de elementos Pa y Pb de Eoplacognathus, pero en los que resulta difícil asignar una especie debido a la vista antero–lateral del elemento Pa (lám. 10, fig. 10) o que el elemento Pb es un ejemplar juvenil (lám. 10, fig. 14 A y B) con caracteres definitorios muy ambiguos. Löfgren y Zhang (2003, pág.731) asignan los elementos figurados por Lehnert (1995) a Yangtzeplacognathus crassus Chen y Zhang en función de caracteres generales en el elemento juvenil y por presentar un diseño poligonal superficial, que estas autoras atribuyen como definitorio del género Yangtzeplacognathus.
Bioestratigrafía de conodontes de la Formación Las Aguaditas
La presente fauna de conodontes de la Formación Las Aguaditas resulta escasa comparada con la abundante asociación rescatada de los últimos metros de la Formación San Juan (Heredia et al., 2005a, 2005b; Mestre, 2010). Los niveles basales resultaron estériles, estando el primer nivel con conodontes a dos metros (Lag 8) de la base de la Formación Las Aguaditas. Este nivel puede atribuirse a la Zona de E. pseudoplanus por la presencia de la especie guía (Figura 5). En la conodontofauna de las muestras Lag 7'/7 a la Lag 0 se identificaron elementos P (formas tardías) de E. suecicus Bergström. De esto se desprende que la Formación Las Aguaditas, en sus primeros metros, incluiría o bien el pase entre las Zonas de E. pseudoplanus a E. suecicus o un hiato representado por la ausencia de formas tempranas y quizás intermedias de E. suecicus que resultan equivalentes a la subzona de Pygodus lunnensis (Figura 5). La subzona inferior de la Zona de E. suecius se correlacionaría con parte de la Zona de Pterograptus elegans (Chen et al., 2006).
Las muestras provenientes de los niveles más altos (calcarenitas), Lag 0 y 1 arrojaron un gran número de conodontes entre los que se distinguen faunas autóctonas (E. suecicus) y faunas alóctonas (i.e. E. pseudoplanus y Dzikodus hunanensis Zhang) evidenciando erosión y reelaboración de niveles más antiguos en este sector de la cuenca (Figura 2).
Atendiendo a los esquemas bioestratigráficos propuestos por Albanesi y Ortega (2002) (Figura 5), Heredia et al. (2005 a) y por Albanesi (en Benedetto et al., 2007) para el Darriwiliano de Precordillera, es importante realizar ciertas reconsideraciones. Con respecto a la Subzona de Histiodella kristinae, que estos autores señalan como la subzona inferior de la Zona de E. suecicus, se debe hacer notar que la primera aparición de la especie H. kristinae Stouge, tal como lo señala Mestre (2010) y Mestre y Heredia (en prensa), se verifica en la parte superior de la Zona de E. pseudoplanus, Subzona de Microzarkodina ozarkodella, en los últimos metros de la Formación San Juan en el sector occidental de la Precordillera Central. Esta afirmación se confirma aquí, pues en los niveles más bajos de la Formación Las Aguaditas aparece H. kristinae con E. pseudoplanus. Por lo tanto, no es aconsejable la utilización de esta especie para definir la parte inferior de la Zona de E. suecicus en la Precordillera argentina. Con respecto a la Subzona superior, es importante ratificar que el hallazgo de moldes en pelitas de la especie guía Pygodus anitae Bergström por Albanesi (1998) en la localidad de Cerro Potrerillo registra efectivamente la sub–zona superior de la Zona de E. suecicus (Figura 5).
Se propone la utilización del esquema bioestratigráfico báltico para el Darriwiliano de la Precordillera, especialmente el basado en el esquema de Löfgren (2000) según lo sugerido en la figura 5 del presente aporte, modificando sustancialmente la propuesta de Albanesi y Ortega (2002), Heredia et al. (2005 a) y Albanesi (en Benedetto et al., 2007).
El análisis biofacial de las asociaciones de conodontes de la Formación Las Aguaditas en el sector del río Las Chacritas sugiere un ambiente oceánico de rampa distal, con libre circulación marina. Se considera entonces, a los conodontes darriwilianos del sector de estudio como vinculados al Reino Noratlántico (Bergström, 1983) o al Dominio de Aguas Templadas en el sentido de Zhen y Percival (2003).
Correlación
Zona de E. pseudoplanus: Los conodontes del tramo basal de la Formación Las Aguaditas en la Quebrada del río de Las Chacritas permiten acotar el lapso temporal a la Zona de E. pseudoplanus y, luego del aporte de Mestre (2010), correlacionarlo con el techo de la Formación San Juan y los tramos basales de las unidades calcáreo–pelíticas suprayacentes que aparecen en distintas localidades de la Precordillera Central. Basado en colecciones propias y en las colecciones de Mestre (2010) este intervalo se considera sincrónico con los pases expuestos en las secciones de la Quebrada de La Brecha (Las Chacritas) y en el Cerro La Chilca (Figura 6). Se considera también el techo de Formación San Juan aflorante en la Quebrada de Las Aguaditas (Figura 6) por la información proporcionada por Albanesi y Ortega (2002, lám. 1, figs. 11 y 4) donde se figura un ejemplar de E. pseudoplanus (no L. variabilis) y un ejemplar de Histiodella kristinae (no H. tableheadensis Stouge), ambas especies caracterizan la Subzona de Microzarkodina ozarkodella, Zona de E. pseudoplanus según el esquema Báltico (Löfgren, 2004). Los conodontes figurados por Voldman et al. (2008), que provienen de la Formación Yerba Loca (Puerta de Ancaucha) (Figura 6), se interpretan aquí como parcialmente correlacionables con la Zona de E. pseudoplanus por la presencia de la asociación E. pseudoplanus y Dzikodus tablepointensis (Stouge) (fig. 4: y, v, w; en Voldman et al., 2008) pues D. tablepointensis está restringido a la Zona de E. pseudoplanus (Zhang, 1998 a, b).
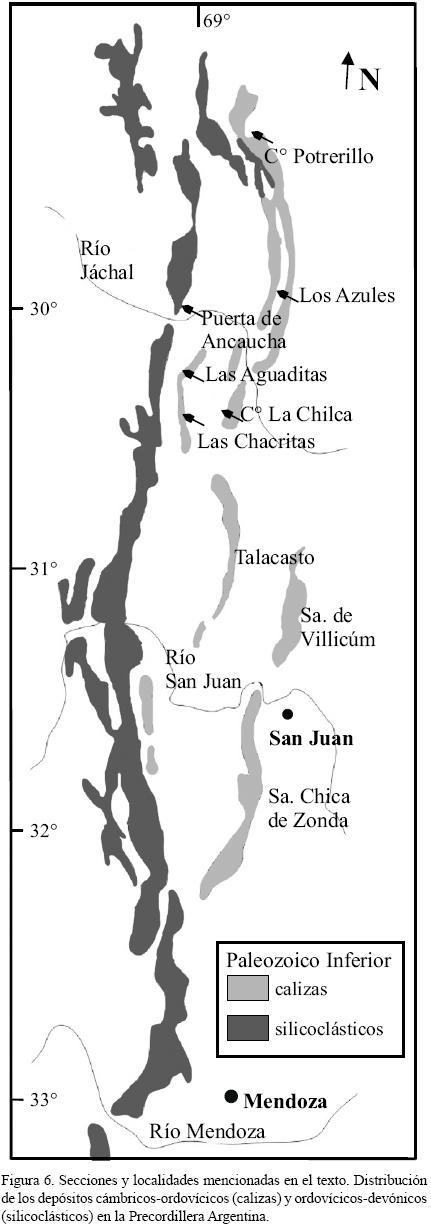
Zona de E. suecicus: El reconocimiento de este lapso temporal para la Formación Las Aguaditas en la sección estudiada, permite su correlación en base a los graptolitos de los tramos medios de la Formación Los Azules (Hünicken y Ortega, 1987; Ortega et al., 2007) en Los Azules, con el Miembro medio de la Formación Gualcamayo en el Cerro Potrerillo (Albanesi, 1998) y con el Miembro Inferior y Lajoso de la Formación Las Aguaditas expuesta en su localidad tipo de la Quebrada de Las Aguaditas (Peralta y Baldis, 1995; Astini, 1995, Brussa, 1996) (Figura 6).
CONCLUSIONES
Se da a conocer la bioestratigrafía de conodontes del Miembro Inferior de la Formación Las Aguaditas de edad darriwiliana media alta señalando que los tramos más bajos están bioestratigráficamente acotados a la Zona de Eoplacognathus pseudoplanus. Se reconoce por primera vez la Zona de Eoplacognathus suecicus en la Precordillera argentina por la aparición de la especie E. suecicus Bergström en la parte inferior de la Formación Las Aguaditas aflorante en el río de Las Chacritas. Se considera que está representado el tramo superior de la Zona de E. suecicus porque los elementos P corresponden a formas tardías de esta especie; asimismo la presencia de Pygodus anitae confirma esta atribución. Este hecho revela la existencia de un hiato representado por los términos inferior y quizás medio de la Zona de E. suecicus. Se correlaciona esta unidad con otras de la Precordillera Central, especialmente por el contenido de graptolitos atendiendo a la estrecha correspondencia de las biozonas de ambos grupos. Los nuevos datos presentados en este estudio permiten realizar una propuesta bioestratigráfica para estos niveles del Ordovícico Medio expuestos en la Quebrada de las Chacritas, al utilizar para la Precordillera argentina el esquema báltico de biozonas de conodontes.
AGRADECIMIENTOS
Al CONICET. A la Lic. A. Mestre por su inestimable colaboración en las tareas de campo y lupa, a la Lic. C. Rodríguez por su colaboración en las tareas de campaña. A la Téc. Química M. González por su labor en laboratorio. A los dos revisores por sus sugerencias que mejoraron la versión original.
REFERENCIAS
Aceñolaza, F.G. Miller, H., Toselli, A.J., 2002, Proterozoic–Early Paleozoic evolution in western South America – a discussion: Tectonophysics, 354, 121–137. [ Links ]
Albanesi, G., 1998, Taxonomía de conodontes de las secuencias ordovícicas del Cerro Potrerillo, Precordillera Central de San Juan, República Argentina: Academia Nacional de Ciencias Córdoba, 12, 99–253. [ Links ]
Albanesi, G., Ortega, G., 2002, Advances on Conodont–Graptolite Biostratigraphy of the Ordovician System of Argentina, en Aceñolaza, F.G. (ed.), Aspects of Ordovician System in Argentina: Serie Correlación Geológica, 16, 143–166. [ Links ]
Astini, R.A., 1994, Significado estratigráfico del Miembro Superior de la Formación San Juan, cordón de Las Chacritas, Ordovícico medio de la Precordillera de San Juan: Revista Asociación Geológica Argentina, 49 (3–4), 365–367. [ Links ]
Astini, R.A., 1995, Sedimentología de la Formación Las Aguaditas (talud carbonático) y evolución de la cuenca precordillerana durante el Ordovícico medio: Revista de la Asociación Geológica Argentina, 50 (1–4), 143–164. [ Links ]
Astini, R.A., 1998, Stratigraphic evidence supporting the rifting, drifting and collision of the Laurentian Precordillera terrane of western Argentina, en Pankhurst, R.J. and Rapela, C.W. (eds.), The Proto–Andean Margin of Gondwana: Geological Society of London Special Publication, 142, 11–33. [ Links ]
Astini, R.A., 2003, The Ordovician Proto–Andean basins, en Benedetto, J.L. (ed.), Ordovician fossils of Argentina: Secretaría de Ciencia y Tecnología, Universidad Nacional de Córdoba, 1–74. [ Links ]
Astini, R.A., Thomas, W.A., 1999, Origin and evolution of the Precordillera terrane of western Argentina: A drifted Laurentian orphan, en Ramos, V.A. and Keppie, J.D. (eds.), Laurentia–Gondwana Connections before Pangea: Geological Society of America Special Paper, 336, 1–20. [ Links ]
Baldis, B.A., Blasco, G., 1974, Trilobites ordovícicos de la comarca de Jáchal, Precordillera Argentina. I. Telephidae: Ameghiniana, 11(1), 71–78. [ Links ]
Baldis, B., Beresi, M., Bordonaro, O., Vaca, A., 1982, Síntesis evolutiva de la Precordillera Argentina, en 5° Congreso Latinoamericano de Geología, Buenos Aires, Argentina: Actas 4, 399–445. [ Links ]
Bassler R.S., 1925, Classification and stratigraphic of McLish and Tulip Creek Formation (Middle Ordovician) of south–central Oklahoma: Oklahoma Geological Survey Bulletin, 141, 1–53. [ Links ]
Benedetto, J.L., Aceñolaza, G., Albanesi, G., Alfaro, M., Brussa, E., Buatois, L., Carrera, M., Cech, N., Esteban, S., Heredia, S., Mángano, M.G., Ortega, G., Ottone, E., Rubinstein, C., Salas, M., Sánchez, T., Toro, B., Tortello, F., Vaccari, N., Waisfeld., B., 2007, Los fósiles del Proterozoico Superior y Paleozoico Inferior de Argentina: Edición del 50 Aniversario de Ameghiniana, Publicación Especial 11, 9–32. [ Links ]
Bergström, S., 1971, Conodont biostratigraphy of the Middle and Upper Ordovician of Europe and Eastern North America, en Sweet, W.C. and Bergström, S. (eds.), Symposium on Conodont Biostratigraphy: Boulder, USA, Geological Society of America Memoir, 127, 83–161. [ Links ]
Bergström, S., 1983, Biogeography, evolutionary relationships and biostratigraphic significance of Ordovician platform conodontes: Fossils and Strata, 15, 35–58. Oslo. [ Links ]
Brussa, E.D., 1996, Las graptofaunas ordovícicas de la Formación Las Aguaditas, Precordillera de San Juan, Argentina. Parte I: Familias Thamnograptoidae, Dichograptidae, Abrograptidae y Glossograptidae: Ameghiniana, 33, 421–434. [ Links ]
Carrera, M.G., 1997, Análisis paleoecológico de la fauna de poríferos del Llanvirniano tardío de la Precordillera Argentina: Ameghiniana, 34(3), 309–316. [ Links ]
Carrera, M.G., Astini, R. 1998, Valoración de las restricciones ambientales durante la transición Arenigiano–Llanvirniano, Ordovícico de la Precordillera Argentina: Revista de la Asociación Geológica Argentina, 53, 41–56. [ Links ]
Chen, X., Zhang, Y.–D., Bergström, S., Xu, H.–G., 2006, Upper Darriwilian graptolite and conodont zonation in the global stratotype section of the Darriwilian stage (Ordovician) at Huangnitang, Changshan, Zhejiang, China: Palaeoworld, 15, 150–170. [ Links ]
Dzik, J., 1976, Remarks on the evolution of Ordovician conodontes: Acta Palaeontologica Polonica, 21, 395—455. [ Links ]
Epstein, A., Epstein, J., Harris, L., 1977, Conodont color alteration– An Index to organic metamorphism: United States Geological Survey Professional Paper, 955, 1–27. [ Links ]
Espisúa, E., 1968, El Paleozoico inferior del Río de Las Chacritas, Dpto. de Jáchal, Provincia de San Juan, con especial referencia al Silúrico: Revista Asociación Geológica Argentina, 23 (4), 297–311. [ Links ]
Finney, S., 2007, The parautochthonous Gondwanan origin of the Cuyania (greater Precordillera) terrene of Argentina: A re–evaluation of evidence used to support an allochthonous Laurentian origin: Geologica Acta, 5(2), 127–158. [ Links ]
Hamar, G., 1964, The Middle Ordovician of the Oslo Region, Norway, 17. Conodonts from the Lower Middle Ordovician of Ringerike: Norsk Geologisk Tidsskrift, 44(2), 243–292. [ Links ]
Hamar, G., 1966, The Middle Ordovician of the Oslo region, Norway. 22 Preliminary report on conodonts from the Oslo–Asker and Ringerike districts. Conodonts from Lower Middle Ordovician of Ringerike: Norsk Geologisk Tidsskrift, 46(1), 27–83. [ Links ]
Heredia, S., Beresi, M., Peralta, S., 2005a, Darriwilian conodont biostratigraphy of the Las Chacritas Formation, Central Precordillera, San Juan Province, Argentina: Geologica Acta, 3 (4), 385–394. [ Links ]
Heredia, S., Rosales, C., Peralta, S., Beresi, M., 2005b, Conodontes de la Formación Las Chacritas (Darriwiliano) y su significado tecto–sedimentario en el Bloque Sierra de La Trampa–Los Blanquitos, Precordillera de San Juan, en XVI Congreso Geológico Argentino, La Plata, Argentina: Actas III, 445–450. [ Links ]
Hünicken, M., Ortega, G., 1987, Lower Llanvirn–Lower Caradoc (Ordovician) conodonts and graptolites from the Argentine Central Precordillera, en Austin, R.L. (ed.). Conodonts: Investigative Techniques and Applications: Chichester, Ellis Horwood Limited, 7, 136–145 [ Links ]
Lehnert, O., 1995, Ordovizische Conodonten aus der Prãkordillere Westargentiniens: Ihre Bedeutung für Stratigraphie und Palãogeographie: Erlanger Geologische Abhandlungen, Erlangen, 125, 1–193. [ Links ]
Löfgren, A., 1978, Arenigian and Llanvirnian conodonts from Jãmtland, northern Sweden: Fossils and Strata, 13, 1–129. [ Links ]
Löfgren, A., 2000, Early to early Middle Ordovician conodont biostratigraphy of the Gillberga quarry, northern Öland, Sweden: GFF, 122, 321–338. [ Links ]
Löfgren, A., 2004, The conodont fauna in the Middle Ordovician Eoplacognathus pseudoplanus Zone of Baltoscandia: Geological Magazine, 141(4), 505–524. [ Links ]
Löfgren, A., Zhang, J.–H., 2003, Element association and morphology in some Middle Ordovician platform–equipped conodonts: Journal of Paleontology, 77, 723–739. [ Links ]
Mestre, A., 2010, Estratigrafía y Bioestratigrafía de Conodontes de la "Transición cuspidal" de la Formación San Juan al sur del paralelo 30°, Precordillera de San Juan: San Juan, Argentina, Universidad Nacional de San Juan, Tesis doctoral, 330 pp. [ Links ]
Mestre, A., Heredia, S., (en prensa), Darriwilian species of the genus Histiodella (Conodonta) in the Argentine Precordillera: Alcheringa, 36. [ Links ]
Ortega, G., Albanesi, G., Frigerio, S., 2007, Graptolite–conodont bioestratigraphy and biofacies of the Middle Ordovician Cerro Viejo succession, San Juan, Precordillera, Argentina: Palaeogeography, Palaeoclimatology, Palaeoecology 245, 245–264. [ Links ]
Peralta, S., Baldis, B., 1995, Graptolites y trilobites del Ordovícico tardío en el perfil del río de Las Chacritas, Precordillera Central de San Juan, Argentina, en V Congreso Argentino Paleontología y Bioestratigrafía, Trelew, Argentina (1994): Actas, 201–205. [ Links ]
Peralta, S., Heredia, S., Beresi, M., 1999 a, Upper Arenig–Lower Llanvirn sequence of the Las Chacritas River, Central Precordillera, San Juan Province, Argentina: Acta Universitatis Carolinae, Geologica, 43 (1/2), 123–126. [ Links ]
Peralta, S., Heredia, S., Beresi, M., 1999 b, Estratigrafía del Ordovícico del río de Las Chacritas, Sierra de La Trampa, Precordillera Central de San Juan, en XIV Congreso Geológico Argentino, Salta, Argentina: Actas I, 397–400. [ Links ]
Purnell, M. A., Donoghue, P. C. J., Aldridge, R.J., 2000, Orientation and anatomical notation in conodontes: Journal of Paleontology, 74, 113–122. [ Links ]
Ramos, V.A., 1995, Sudamérica: un mosaico de continentes y océanos: Ciencia Hoy, 6, 24–29. [ Links ]
Ramos, V.A., Dallmeyer, R.D., Vujovich, G., 1998, Time constraints on the Early Palaeozoic docking of the Precordillera, central Argentina, en Pankhurst, R.J., and Rapela, C.W. (eds.). The Proto–Andean Margin of Gondwana: London, England, Geological Society, Special Publication, 142, 143–158. [ Links ]
Sarmiento, G., 1985, La Biozona de Amorphognathus variabilis –Eoplacognatus pseudoplanus (Conodonta), Llanvirniano inferior, en el flanco oriental de la sierra de Villicúm, en 1° Jornadas Sobre Geología de Precordillera, San Juan, Argentina: Asociación Geológica Argentina, Actas, 2, 119–123. [ Links ]
Sarmiento, G., 1991, Conodontos de la Zona de Suecicus (Llanvirniano inferior) en la sierra de Villicúm, Precordillera de San Juan, Argentina: Revista Española de Micropaleontología, 23 (3), 113–132. [ Links ]
Stone, J., 1987, Review of investigative techniques used in the study of conodontes, en Austin, R., (ed.), Conodonts: Investigative Techniques and Applications: Chichester, Ellis Horwood Limited, 17–34. [ Links ]
Stouge, S., 1984, Conodonts of the Middle Ordovician Table Head Formation, western Newfoundland: Fossils and Strata, 16, 1–145. [ Links ]
Viira, V., 1974, Ordovician conodonts of east Baltic: Eesti NSV Teaduste Akadeemia Geologia Institut Valgus, 1–142. [ Links ]
Voldman, G., Albanesi, G., Do Campo, M., 2008, Conodont palaeothermometry of contact metamorphism in Middle Ordovician rocks from the Precordillera of western Argentina: Geological Magazine, 145, 449–462. [ Links ]
Zhang, J.–H., 1998a, Middle Ordovician conodonts from the Atlantic faunal Region and the evolution of key conodont genera: Stockholm, Sweden, Meddelanden frân Stockholms Universitets Institution för Geologi och Geokemi, tesis doctoral, pp. 27. [ Links ]
Zhang, J.–H., 1998b, Conodonts from the Guniutan Formation (Llanvirnian) in Hubei and Hunan Provinces, south–central China: Stockholm Contributions in Geology, 46, 1–161. [ Links ]
Zhang, J–H., 1999, Review of the Ordovician conodont zonal index Eoplacognathus suecicus Bergström, 1971: Journal of Paleontology, 73(3), 487–493 [ Links ]
Zhen, Y., Percival, G., 2003, Ordovician conodont biogeography reconsidered: Lethaia, 36, 357–370. [ Links ]

