










Servicios Personalizados
Revista
Articulo
 Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista mexicana de ciencias geológicas
versión On-line ISSN 2007-2902versión impresa ISSN 1026-8774
Rev. mex. cienc. geol vol.28 no.3 Ciudad de México dic. 2011
Biostratigraphy and paleoecology of the Qom Formation in Chenar area (northwestern Kashan), Iran
Bioestratigrafía y paleoecología de la Formación Qom en el área de Chenar (noroeste de Kashan), Irán
Elham Behforouzi* and Amrollah Safari
Department of Geology, Faculty of Sciences, University of Isfahan, Isfahan, Iran, 81746–73441. *elibehforouzi@yahoo.com.
Manuscript received: December 18, 2010
Corrected manuscript received: June 16, 2011
Manuscript accepted: June 26, 2011
ABSTRACT
A study of large benthic foraminifera from the 147 m–thick Qom Formation in the Chenar area (northwestern Kashan) is reported. One hundred and twelve thin sections were prepared and the distribution of benthic foraminifera was analyzed to reconstruct paleoenvironmental conditions. Study of these sections led to the identification of 28 genera and 38 species. On the basis of the recognized foraminifera, the section is comparable to Lepidocyclina– Operculina– Ditrupa Assemblage zone, and the age of the Qom Formation in the studied section is assigned to the Oligocene.
Evidence of sea level changes is observed from bottom to top of the studied section. On the basis of large benthic foraminifera assemblages and microfacies features, three major depositional environments (inner shelf, middle shelf and outer shelf) were defined. The inner shelf facies is characterized by wackstone–packstone, dominated by miliolid and small perforate foraminifera. The middle shelf facies is represented by packstone–grainstone with diverse assemblage of large perforate benthic foraminifera. Outer shelf facies is dominated by large perforate benthic foraminifera as well as planktic foraminifera. The distribution of the Oligocene large benthic foraminifera in the studied area indicates that shallow marine carbonate sediments of the Qom Formation were deposited in a photic zone of tropical to subtropical environments. Finally, the correlation between the study area and some other sections of Central Iran indicates that sedimentation of the Qom Formation is continuedfrom Late Rupelian to Chattian in northwest and Late Rupelian to Aquitanian in southeast direction.
Key words: biostratigraphy, benthic foraminifera, Qom Formation, Oligocene. Aquitanian, Iran.
RESUMEN
Se reporta el estudio de foraminíferos bentónicos grandes de la Formación Qom, con 147 m de espesor, en el area de Chenar (noroeste de Kashan). Ciento doce secciones delgadas fueronpreparadasy la distribución de foraminíferos bentónicosfue analizada para reconstruir las condiciones paleoamebietales. El estudio de esas secciones llevó a la identificación de 28 géneros y 38 especies. Con base en los foraminíferos identificados, la sección es atribuible a la zona de Lepidocyclina– Operculina– Ditrupa y la edad de la Formación Qom en la sección estudiada es asignable al Oligoceno.
Evidencias de cambios del nivel del mar pueden ser observados desde la base y hacia a cima de la sección estudiada. Con base en los ensambles de foraminíferos bentónicos grandes y en las características de las microfacies, tres ambientes de depósito fueron definidos (plataforma interna, media y externa). Las facies de plataforma interna se caracterizan por wackstone–packstone, dominada por miliólidos y pequeños foraminíferos perforados. La plataforma media está representada por packstone–grainstone, con diversos ensambles de foraminíferos bentónicos grandes perforados. Las facies de plataforma externa están dominadas porforaminíferos bentónicos perforados grandes, así como por foraminíferos planctónicos. La distribución de los foraminíferos bentónicos grandes del Oligoceno indica que los sedimentos marinos carbonatados someros de la Formación Qom fueron depositados en la zona fótica de un ambiente tropical a sub–tropical. Finalmente, la correlación entre el área estudiada y otras secciones de Irán Central indica que la sedimentación de la Formación Qom fue continua en dirección noroeste desde el Rupeliano Tardío al Chattiano, y lo fue en dirección sureste del Rupeliano Tardío al Aquitaniano.
Palabras clave: bioestratigrafía, foraminíferos bentónicos, Formación Qom, Oligoceno, Aquitaniano, Irán.
INTRODUCTION
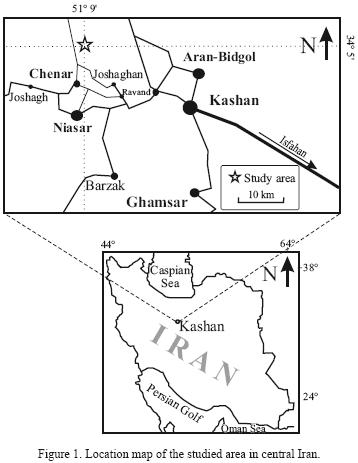
In this study, the foraminifera assemblage in the stratigraphic section located at Chenar village, 50 km northwest of Kashan, is described. The study area exposes Cenozoic sediments and is located at 51°09'02"E longitude and 34°05'37"N latitude (Figure 1).
Due to the high evolution rate, abundance, widespread distribution and abrupt extinction of species, benthic foraminifera are considered an important tool for the study of the biostratigraphy and evolution of genera and species, relative depth or paleoenvironmental reconstruction (Vaziri–Moghaddam et al., 2010). Considering the distribution of foraminifera and analyzing some factors like light, temperature, nutrient supply, substrate, hydrodynamic energy, depth, water locomotion and symbiosis, the paleoenvironment could be reconstructed. There are some similarities between the benthic foraminifera assemblages of Qom Formation and Asmari Formation and due to lack of a formal proposed biozonation in the Qom Formation, the biozonation offered by Wynd (1965) and Laursen et al. (2009) for the Asmari Formation were used in this investigation.
Geological investigation of the Qom Formation in Central Iran started with the work of Tietze (1875). Subsequent workers such as Stahl (1911), Riben (1935), Furon and Marie (1939), Furon (1941), Furrer and Soder (1955), Gansser (1955), Dozy (1944, 1955), Abaie et al. (1964) and Bozorgnia (1966) reported on the marine sediments of the Qom Formation. A few foraminiferal species were illustrated by Furon and Marie (1939), Furon (1941), Furrer and Soder (1955), Abaie et al. (1964) and Bozorgnia (1966). Rahimzade (1994) collected the names of most researchers, part of whose work covered the Qom Formation.
Furrer and Soder (1955) divided the Qom Formation into six members; Abaie et al. (1964) increased the number of members to ten. Abaie et al. (1964) noted that two members, c–1 and c–3, were the main objectives in exploration wells, due to fracture development enhancing porosity/permeability and hydrocarbon shows. Bozorgnia (1966) proposed and introduction ten members for the Qom Formation and distinguished several local basins of deposition in Central Iran, on the basis of its lithological characters. Rahaghi (1973, 1976, and 1980) suggested Oligo–Miocene age for the Qom Formation. Okhravi and Amini (1998) reconstructed palaeoenvironment of the f–member of the Qom Formation based on microfacies analysis. Based on echinoderms and microfosils, Khaksar and Maghfouri–Moghaddam (2007) proposed Middle to Late Oligocene age for the Qom Formation. Aalaeobiogeographic reconstruction of the Qom Formation was proposed by Reuter et al., 2007.
METHODS AND STUDY AREA
Field work was concentrated on an outcrop of the Qom Formation, located 10 km to the north of Chenar village. A 147 m thick section was measured in detail, and a total of 112 specimens were sampled during the detailed field investigation. Samples were taken from the carbonate and marly layers almost every meter according to facies variation. Thin sections were provided for harder litologies whilst softer litologies were disaggregated and the foraminifera picked and analyzed. Disaggregated samples were wet sieved through a 151 Lim. Thin sections were studied under the microscope for the analysis of benthic foraminifera. Taxonomic classification was based on Loeblich and Tappan (1988), Adams and Bourgeois (1967) and Adams (1969).
GEOLOGICAL SETTING
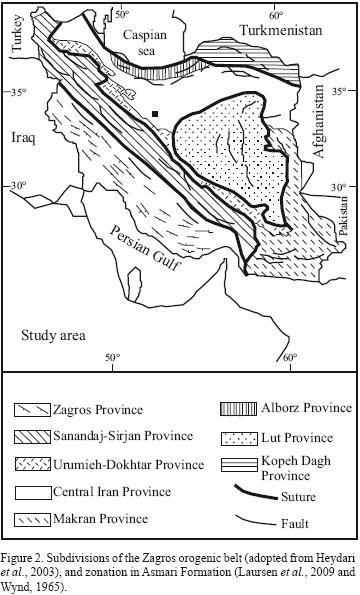
On the basis of the sedimentary sequence, magmatism, metamorphism, structural setting and intensity of deformation, Iranian plateau has been subdivided into eight continental fragments, including, Zagros, Sanandaj–Sirjan, Urumieh–Doktar, Central Iran, Alborz, Kopeh–Dagh, Lut and Makran (Heydari et al., 2003). The study area is located in the Central Iran basin (Figure 2).
During the Early Paleogene, the Tethyan seaway was a wide ocean that connected the two major oceanic realms, the Atlantic and the Pacific (Schustr and Wielandt, 1999). The subduction and final collision of the African–Arabian plate around Eocene–Oligocene boundary was accompanied by the vanishing of the Tethyan seaway, the disconnection of the Atlantic and the Pacific Ocean and the birth of the Indian Ocean and the Mediterranean. As a consequence, Central–Iranian paleogeography changed dramatically by the development of a volcanic arc which separated a fore arc from a back arc basin during Eocene times. Marine sedimentation of the Qom Formation began during the Oligocene and continued to the end of the Early Miocene in the Esfahan–Sirjan fore arc and in the Qom back arc basin (Schustr and Wielandt, 1999).
BIOSTRATIGRAPHY OF FORAMINIFERA
Larger benthic foraminifera are widely distributed in the Tertiary carbonate platform of the Qom Formation. They developed complicated internal structures which can be identified when they are randomly thin sectioned. These organisms can provide complete and detailed evidence for biostratigraphic analysis of the shelf limestone because of rapid diversification, abrupt extinction and abundance (Beavingtone–Penny and Racey, 2004).
The sedimentary deposits of the study area yielded abundant larger benthic foraminifera, therefore, biostratigraphic zonation is based on these organisms. So far, a formal biostratigraphic framework has not yet been established for the Qom Formation. However, based on foraminiferal similarities a general agreement exists to correlate the Qom Formation (Cental Iran Basin) with its coeval counterpart in the Zagros basin in southwest Iran (Asmari Formation) (Stöcklin 1952; Bozorgnia 1966 and Kashfi 1988). Therefore, biozonations established for the Qom Formation in this work are largely based on the biozonations of Wynd (1965) and Laursen et al. (2009), which was used for the Asmari Formation that comprises an Oligocene to Early Miocene carbonate sequence (Table 1).
From base to top a two foraminifera assemblages were recognized in the studied section (Figure 3):
Assemblage 1: Is characterized by the presence of Lepidocyclina sp., Eulepidina sp., Eulepidina dilatata, Eulepidina elephantina, Nephrolepidina cf. marginata, Nephrolepidina cf. tournoueri, Nephrolepidina sp., Operculina complanata, Operculina sp., Sphaerogypsina globulusa, Haplophragmium slingri, Planorbulina sp., Neorotalia viennoti, Neorotalia sp., Valvulinid sp., Amphistegina sp., Amphistegina lessonii, Elphidium sp., Bigenerina sp., Textularia sp., Discorbis sp., Quinqueloculina sp., Triluculina trigouenula, Globorotalia cf. nana, Globorotalia siakensis (Figures 4 and 5). This assemblage corresponds to the "Lepidocyclina–Operculina–Ditrupa assemblage zone". Assemblage zone of Wynd (1965) and Laursen et al. (2009) applied for the Asmari Formation. This assemblage is attributed to the Oligocene and is correlated with the lower part of the Asmari Formation.
Assemblage 2: Lepidocyclina–Operculina–Ditrupa assemblage zone. The assemblage has this associated fauna: Planorbulina sp., Eulepidina dilatata, Haplophragmium slingri, Rotalia viennoti, and algae. This zone ranges from the Rupelian into the Chattian (Laursen et al., 2009).
PALEOECOLOGY
In this section, an attempt to investigate factors that influence the benthic foraminifera distribution in the Qom Formation is presented. In this regards, environmental agents such as nutrient supply, light, temperature, water locomotion, substrate, salinity, hydrodynamic energy, depth and symbiosis are very significant. In terms of the paleoecology, paleoecological constraints porposed by Hallok and Glenn (1986) include three families observed in studied section (Nummulitidea, Amphisteginidea and Peneroplidea).
Nutrient supply
Large benthic foraminifera are highly adapted to stable, oligotrophic and nutrient–deficient conditions, but they cannot respond competitively when nutrient resources become abundant (Hallock, 1985). Inorganic, "biolimiting" nutrients enter shallow–water communities principally by turnover or upwelling of deeper waters, by run–off from land or by advection from areas of upwelling or runoff (Hallock and Schlager, 1986).
Large symbiont–bearing benthic foraminifera are compatible to nutrient deficiency conditions and in such situation, the algal symbiont uses organic waste substances of the host and CO2 originated from host respiration for photosynthesis (Beavingtone–Penney and Racey, 2004). And so, the produced substances supplies nutrient for the host.
In the studied section, semi–restricted lagoon environments inferred by the presence of perforate and imperforate foraminifera indicate mesotrophic to eutrophotic conditions and shallow to deep marine environments point to mesotrophic to oligotrophic conditions. Show that Lepidocyclina harboured endosymbiontic algae to provide sufficient light for the photosynthesis of the symbionts (Chaproniere, 1975). Numerous chambers of twenty tests of Nephrolepidina cf. tournouer indicated in sample (B16) (Figure 6).

Light
The abundance of robust and thick tests of Operculina in the study area reflects increasing light intensity (Figure 7). Light intensity and hydrodynamic force control test morphology through symbiotic interactions. In shallow, well–lit waters the calcification rate in foraminiferal test is enhanced by photosynthetic symbionts, in order to prevent photoinhibition of symbiotic algae within the test or test damage in turbulent water (Beavingtone–Penney and Racey, 2004).

Larger foraminifera are sensitive to changes in environmental factors including light intensity and hydrodynamic force along depth gradients which are mirrored by changes in test shape and size of symbiont–bearing foraminifera (Beavingtone–Penney and Racey, 2004). The shallow dwelling microperforated hyaline foraminifera produce small, robust and ovate tests and increase calcification rate which brings about a thick wall structure to prevent photoinhibition of the endosymbiont within the test in highly illuminated shallow waters, or test damages in mobile substrates, while hyaline foraminifera from greater depths shifts to larger, thinner and more transparent test to thrive in strongly reduced light intensity (Beavingtone–Penney and Racey, 2004). On the basis of the dependence of carbonate production to light penetration, three different groups of benthic organisms could be considered.
Red algae and some hyaline benthic foraminifera such as Heterostegina, Operculina and Lepidocyclina identified in the Qom Formation samples are indicators of dysphotic zones. Bassi et al. (2007) divided the photic zone into upper and lower parts; in this classification, Neorotalia live in the upper part of the upper photic zone, and Heterostegina, Operculina and Lepidocyclina are dominant in the lower part of the upper photic zone, whilst in the lower photic zone Lepidocyclina is developed.
Light–independent biota includes bryozoans, mollusks, crinoids, brachiopods and sponges that are widespread in studied section (sample B12). Organisms that compose the light–dependent biota identified include green algae, reef–builder corals and miliolids constituting indicators of photic zone.
Several large foraminifers that host algal symbionts present a more evolved skeletal structure than those without symbiont. This process is also seen in Lepidocyclina (Figure 4). Large benthic foraminifers host unicellular eukaryotic algal symbionts that are dependent on light and nutrient and are therefore restricted to euphotic zone (Romero et al., 2002). Symbiont–bearing large foraminifera are restricted to warm water of tropical realms where water temperature is higher than 14–18 °C in the coldest months of the year (Renema, 2006).
Water motion
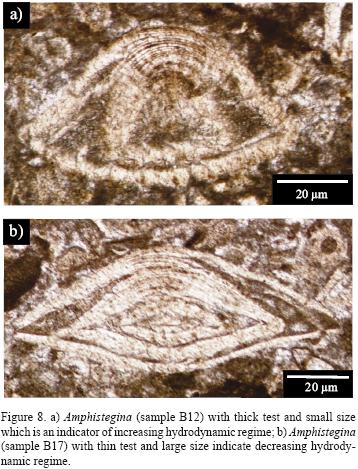
This factor influences the test shape (diameter/thickness ratio). Generally, light intensity and water motion promotes the formation of secondary layers in foraminiferal tests. On the contrary, when the light intensity and the hydrodynamics are weak the growth rate decreases. In the studied area, Amphistegina from lagoon environment with high energy and intense light are thicker than those of deeper environments with lower energy (Beavingtone–Penney and Racey, 2004) (Figure 8).
Substrate nature
Substrate nature also depends on water turbulence. Foraminifera which live on a coarse grain substrate have thicker tests and are fusiform. Amphistegina also prefer hard substrates with high energy (Figure 8), while Operculina live on soft and muddy substrates and have thin shells (Figure 8).
Salinity
High salinity plays a preventative role in growth and evolution of large foraminifera, but medium degree of salinity is not so effective. In semi–restricted lagoon environment, because of the connection with open marine environments and low percent of salinity, perforate and imperforate foraminifers can be observed together (Figure 9a), whereas, considering normal salinity in shallow parts of open marine of the studied section, only perforate foraminifers have been found (Figure 9b). The presence of stenohaline organisms (echinoid and bryozoan) is an indicator of marine normal salinity (Figure 9c, 9d).
Hydrodynamic energy and depth
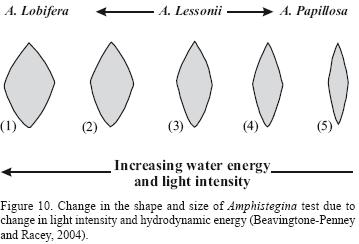
The occurrence of benthic organisms reflects their compatibility in high or low energy environments (Flugel, 2004). For instance, in shallow environments within the photic zone, with increasing water motion, benthic foreminifers shape changes. It means that high energy causes the test to be thick (increasing in carbonate production) and decreases its growth rate and eventually reduces their test size (Beavingtone–Penney and Racey, 2004). Haynes (1965) stated that the shape of large foraminifera changes under the influence of hydrodynamic conditions and its symbiotic relationship with algae. The author expressed that species with free life (non–epizoan) and spherical species are indicators of reef environment cleaned by flows, while fragile thin tests with maximum surface/volume ratio could be found in calm environments with low light. Generally, foraminifers in compatibility with high energy, creates lamellar and thick tests like Amphistegina. The elongated Operculina with thin test found in this work is an indicator of deep environments (Beavingtone–Penney and Racey, 2004). In this research, a comparison is shown between thick Amphistegina with lamellar tests in high energy lagoon environment and elongated Amphistegina that is indicator of low energy environment (Figures 10 and 11).
CONCLUSION
On the basis of the foraminifera recognized in the studied section, the Qom Formation is comparable to the Lepidocyclina–Operculina–Ditrupa Assemblage Zone. The age of the Qom Formation in the studied section is Oligocene.
The carbonate rocks of the study area contain a photozoan association composed predominantly of large benthic foraminifera and coralline red algae in association with corals, heterotrophs. This photozoan assemblage indicates oligotrophic conditions. Moreover, the abundance and association of large benthic foraminifera with coralline red algae are referred to as foralgal facies developed in shallow, warm water environments of the photic zone where oligotrophic condition was prevalent.
Based on the paleoecology and lithology, three distinct depositional setting can be recognized: inner shelf, middle shelf and outer shelf.
Inner shelf facies contain abundant imperforate tests of foraminifera. Middle shelf facies are characterized by abundant large perforate foraminifer tests. Toward the basin, planktic foraminifera and large foraminifers with perforate tests occur contemporaneously. Basin (outer shelf) facies is marked by high planktic foraminifera contents embedded in wackstone.
ACKNOWLEDGMENTS
In particular, we would like to express our most sincere gratitude to Prof. Dr. Francisco Vega for his helpful advice and guidance. The authors wish to thank the reviewers for their helpful and constructive comments. We also thank Dr. Hossein Vaziri–Moghaddam, Dr. Ali Bahrami, Dr. Akbar Ghazi–Fard and Dr. Mohammad Ali Makki–Zade for his support and thoughtful comments. We also would like to thank the University of Isfahan for providing financial support. Also we thank the Revista Mexicana de Ciencias Geológicas reviewers for their constructive comments.
REFERENCES
Abaie, I., Ansari, H.J., Badakhshan, A., Jaafari, A., 1964, History and development of the Alborz and Sarajeh fields of Central Iran: Bulletin of Iranian Petroleum Institute 15, 561–574. [ Links ]
Adams, T.D., 1969, The Asmari Formation of Lurestan and Khuzestan Proviences: National Iranian Oil Company Report 1151 , unpublished. [ Links ]
Adams, T.D., Bourgeois, F., 1967, Asmari biostratigraphy: Iranian Oil Operating Companies, Geological and Exploration Division, Report 1074. [ Links ]
Bassi, D., Hottinger, L. Nebelsick, H., 2007, Larger Foraminifera from the Upper Oligocene of the Venetian area, northeast Italy: Palaeontology, 5(4), 845–868. [ Links ]
Beavingtone–Penney, S.J., Racey, A., 2004, Ecology of extant nummulitids and other larger benthic foraminifera. applications in Paleoenvironmental analysis: Earth Science Review, 67(3–4), 219–265. [ Links ]
Bozorgnia, F., 1966, Qom Formation stratigraphy of the Central basin of Iran and its intercontinental position: Bull. Iran Pet. Inst., 24, 69–76. [ Links ]
Chaproniere, G.C.H., 1975, Paleoecology of Oligocene–Miocene larger Foraminiferida: Australia: Alcheringa, 1, 37–58. [ Links ]
Dozy, J.J., 1944, Comments on geological report no.1 by Thiebaud (on the Qum–Saveh area): Geological Report 308. [ Links ]
Dozy, J.J., 1955, A Sketch of past Cretaceous volcanism in Central Iran: Leidsche, Geologische Mededeelingen 20, 48–57. [ Links ]
Flugel. E., 2004, Microfacies of Carbonate Rocks, Analysis, Interpretation and Application: Berlin, Springer–Verlag, Berlin, 976 pp. [ Links ]
Furon, R., 1941, Geologie du plateau Iranien (Perse–Afghanistan–Beloutchistan): Mémoires du Muséum National d'Histoire Naturelle, Nouvelle Serie, Paris 7 (2), 177–414. [ Links ]
Furon, R., Marie, P., 1939, Sur la microfaune des marnes aquitaniennes Lepidocyclines de Qoum (in Persian): Compte Rendu Sommaire des Séances de la Société Géologique de France, 79–80. [ Links ]
Furrer, M.A., Soder, P.A., 1955. The Oligo–Miocene marine formation in the Qom region (Central Iran): Proceedings of the 4th World Petroleum Congress, Rome, Section I/A/5, 267–277. [ Links ]
Gansser, A. , 1955. New aspects of the geology in Central Iran, in Proceedings of the 4th World Petroleum Congress, Rome, Section I/A/5, 279–300. [ Links ]
Hallock, P., 1985, Why are larger foraminifera large?: Paleobiology 11, p.195–208. [ Links ]
Hallock, P., Glenn, E.C., 1986, Larger foraminifera: a tool for paleoenvironmental analysis of Cenozoic depositional facies: Palaios, 1(1), 55–64. [ Links ]
Hallock, P., Schlager, W., 1986, Nutrient excess and the demise of coral reefs and carbonate platforms: Palaios 1, 389–398. [ Links ]
Haynes, J.R., 1965, Symbiosis wall structure and habitate in foraminifera: Special Publication–Cushman Foundation for Foraminiferal: Research, 16, 40–43. [ Links ]
Heydari, E., Hassanzadeh, J., Wade, W.J., Ghazi, A.M., 2003, Permian–Triassic boundary interval in the Abadeh section of Iran with implications for mass extinction, Part 1–Sedimentology: Paleogeography, Paleoclimatology, Paleoecology, 193(3), 405–423. [ Links ]
Kashfi, M.S., 1988, Evidence for non–collision geology in the Middle East: Journal of Petroleum Geology 11 (4), 443–460. [ Links ]
Khaksar, K., Maghfouri–Moghadam, I., 2007, Paleontological study of the echinoderms in the Qom Formation (Central Iran): Islamic Azad University, Earth Sci. Res. 11(1), 57–79. [ Links ]
Laursen, G.V., Monibi, S., Allan, T.L., Pickard, N.A., Hosseiney, A., Vincent, B., Hamon, Y., Van–Buchem, F.S.P., Moallemi, A., Druillion, G., 2009, The Asmari Formation revisited: changed stratigraphic allocation and new biozonation: Shiraz, First International Petroleum Conference & Exhibition, European Association of Geoscientists and Engineers. [ Links ]
Loeblich, A.R., Tappan, J.H., 1988, Foraminiferal Genera and their Classification. Van Nostrand Reinhold. 2 vols. pls. 847. New York, 869 p. [ Links ]
Okhravi, R., Amini, A., 1998, An example of mixed carbonate–pyroclastic sedimentation (Miocene, Central Basin Iran): Sedimentary Geology, 118, 37–57. [ Links ]
Rahaghi, A., 1973, Étude de quelques grands foraminifères de la Formation de Qum (Iran Central): Revue de Micropaleontologie, 16, 23–38. [ Links ]
Rahaghi, A., 1976, Contribution a l'É tude de quelques grands foraminifères de l'Iran: Société National Iranienne des Pétroles Laboratoire de Micropaléontologie, Publication 6, 1–79. [ Links ]
Rahaghi, A., 1980, Tertiary faunal Assemblage of Qum–Kashan, Sabzewar and Jahrum area: National Iranian Oil Company, Geological Laboratories, Publication 8. [ Links ]
Rahimzade, F., 1994, Treatise on the Geology of Iran: Oligocene–Miocene, Pliocene (in Persian): Ministry of Mines and Metals, Geological Survey of Iran,12. [ Links ]
Renema, W., 2006, Large benthic foraminifera from the deep photic zone of a mixed siliciclastic–carbonate shelf of East Kalimantan: Indonesia, Marine Micropaleontology, 58, 73–82. [ Links ]
Reuter, M., Pillar, W.E., Harzhauser, M., Mandic, O., Berning, B., Rogl, F., Kroh, A., Aubry, M.P., Wielandt, U., Hamedani, A., 2007, The Oligo–Miocene Qom Formation (Iran): evidence for and early Burdigalian restriction of Tethyan Seaway and clouser of its Iranian getways: International Journal of Earth Sciences, 98, 627–650. [ Links ]
Riben, H., 1935. Contribution a la géologie de l'Azerbeidjan Persan: Bulletin de la Société Neuchateloise des Sciences Naturelles, 59 (1934), 20–144. [ Links ]
Romero, J., Caus, E., Rossel, J., 2002, A Model for the Palaeoenvironmental distribution of larger foraminifera based on Late Middel Eocene deposits on the margine of the south Pyrenean Basine (SE Spain): Palaeogeography, Palaeoeclimatology, Palaeoecology, 179, 43–56. [ Links ]
Schustr, F., Wielandt, U., 1999, Oligocene and Early Miocene coral faunas from Iran: palaeoecology and palaeobiogeography: International Journal of Earth Sciences, 88, 571–581. [ Links ]
Stahl, A.F., 1911, Persien, in Handbuch der Regionalen Geologie: Heidelberg (Winter), Hft. 8, 5(6), 46 pp. [ Links ]
Stocklin, J., 1952, Stratigraphical investigation in the Qom–Arak–Gulpaigan Delidjan area: National Iranian Of Oil Company, Report 95. [ Links ]
Tietze, E., 1875, Ein Ausflug nach dem Siahkuh (Schwarzer Berg) in Persien. Mitteilungen der Geographischen Geographischen Gesellschaft Wien 18 (8), 257–267. [ Links ]
Vaziri Moghaddam, H., Kimiagari, M., Taheri, A., 2010, Principles of Foraminifera: Isfahan, Isfahan, 366 p. [ Links ]
Wynd, J., 1965, Biofacies of Iranian oil consortium agreement area: Iranian Oil Offshore Company, Report 1082, unpublished. [ Links ]

