










Servicios Personalizados
Revista
Articulo
 Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista mexicana de ciencias geológicas
versión On-line ISSN 2007-2902versión impresa ISSN 1026-8774
Rev. mex. cienc. geol vol.28 no.3 Ciudad de México dic. 2011
Extinct ground sloth dermal bones and their role in the taphonomic research of caves: the case of Baño Nuevo–1 (Andean Central Patagonia, Chile)
Huesos dérmicos de perezosos terrestres extintos y su papel en la investigación tafonómica de cuevas: El caso de Baño Nuevo–1 (Patagonia Central, Chile)
Patricio López–Mendoza1* and Francisco Mena–Larraín2
1 Universidad Católica del Norte, Instituto de Investigaciones Arqueológicas y Museo Gustavo Le Paige (IIAM), Calle Gustavo Le Paige N° 380, San Pedro de Atacama, Chile.*patriciolopezmend@yahoo.es.
2 Centro de Investigación en Ecosistemas de la Patagonia (CIEP), Calle Francisco Bilbao N° 449, Coyhaique, Chile.
Manuscript received: April 8, 2011
Corrected manuscript received: June 5, 2011
Manuscript accepted: June 5, 2011
ABSTRACT
The results of the taphonomic analysis of extinct ground sloth dermal bones from the Baño Nuevo–1 archaeological site (Andean Central Patagonia, Chile) are presented. These pieces were recovered on all but one of the stratigraphic layers, including both the deepest levels (dated between 13,500 and 10,500 BP) and the most recent one (dated to the last century) and thus reveal complex formation processes. Since it is important to take into account this all too common depositional complexity, particularly when dealing with cave and rock shelter environments and with extinct fauna, we study the abundant ground sloth dermal bones found at the Baño Nuevo–1 cave. This effort is oriented to "make the most" (extract as much information as possible) from a single material, that is particularly abundant at this site and we believe it is especially suited to answer taphonomic questions.
Key words: Dermal bones, Extinct Ground Sloth, Taphonomy, Central Patagonia, Baño Nuevo–1 cave, Chile.
RESUMEN
Se presentan los resultados del análisis tafonómico de huesos dérmicos de perezosos terrestres extintos del sitio arqueológico y paleontológico Baño Nuevo–1 (Patagonia Central, Chile). Estos restos fueron recuperados tanto en los niveles más profundos (datados entre 13,500 y 10,500 AP) como en el más reciente (datado en el siglo recién pasado), lo que revela complejos procesos de formación. Debido a la relevancia de los procesos de formación de los depósitos estratigráficos, especialmente en refugios rocosos con restos de fauna extinta, el presente trabajo estudia los huesos dérmicos de perezosos extintos del sitio cueva Baño Nuevo–1. Este esfuerzo está orientado a extraer la mayor cantidad de información posible de estos restos óseos que son abundantes en el sitio y que creemos son ideales para responder a preguntas tafonómicas.
Palabras claves: Huesos dérmicos, Perezoso extinto, Tafonomía, Patagonia Central, Baño Nuevo–1, Chile.
INTRODUCTION
Dermal bones are rather peculiar structures found on some species within the Xenarthra superorder (orders Pilosa and Cingulata). Dermal bones have particular morphologi–cal and histological traits that have been described both for extinct and extant species (Carlini et al., 1997; Vickaryous and Hall, 2006; Hill, 2006; Chávez Aponte et al., 2008; Krmpotic et al., 2009). Among Cingulata (armadillos, pam–patheres and glyptodons) these bones are found articulated into a strong carapace with polygonal shapes that either meet side to side or are imbricated to one another (Hill, 2006; Chávez Aponte et al., 2008). Among the Pilosa dermal bones are isolated specimens which do not articulate with each other, and therefore lack articular faces (Hill, 2006).
Dermal bones are common in Late Pleistocene paleontological and archaeological sites in Patagonia and Tierra del Fuego (Borrero, 1986; Massone, 1987; Borrero et al., 1991, 1997; Latorre, 1998; Menegaz and Nami, 1999; Mena et al., 2000; San Román et al. 2000; Borrero and Martin, 2008; Martin, 2008). This situation is due to the abundant remains of animals of the Mylodon genus in several rock shelters throughout the region. In the case of Milodon Cave at Última Esperanza, Chile even soft tissues such as dung and hide have preserved exceptionally well (Hauthal 1899; Emperaire and Laming 1954, among others) The presence of Mylodon remains in caves and rock shelters is attributable both to the redundant use of these sites by the animal and to their being carried in by predators of several different sizes, as well as by hunting and/or scavenging by ancient human populations (Martin, 2008). These different processes produce specific taphonomic and zooarchaeological modifications that are best seen on certain fossils remains (e.g., dermal bones) due both to their size, structure and/or surface and to predator's preference of certain taxa and anatomical units, among other factors (Martin and Borrero, 1997; Martin, 2008).
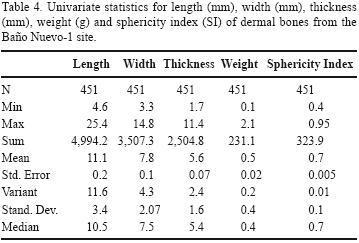
The aim of this article is to present the results of the taphonomical analysis of Xenarthra dermal bones from Baño Nuevo–1 (Andean Central Patagonia, Chile). We chose this material to discuss taphonomical and zooarchaeological issues since dermal bones are temporally diagnostic, they are often found in fairly large numbers on sheltered spaces with or without human occupations and their shape and size make them adequate targets of formation processes traces. We find these specimens particularly useful to identify several kinds of alterations, although they are generally ignored, being merely counted, given their poor value for taxonomic quantification of assemblages. We herein pro–vide not only a general quantification, but also record their main dimensions (length, width, thickness and weight) and their spatial and chronological distribution as well as their microscopic features in order to address problems such as vertical migration, the role of carnivores in the species bones accumulation within sheltered spaces and the possible human impact on this animal in the final Pleistocene.
THE BAÑO NUEVO–1 SITE: STRATIGRAPHY AND DATES
Baño Nuevo–1 is located some 80 km to the northeast of the modern city of Coyhaique (45°17'S – 71°32,W), within an area that is characterized by large plains of moderate slopes traversed by several ravines including Arroyo Goichel and Arroyo Baño Nuevo that drain to the Ñirehuao river, one of the main tributaries of the Mañihuales river, part of the large basin of the Aysén river (Figure 1). The cave has a depth of 20 m and an average width of 4 m, with a restricted access due to the entrance in the late Pleistocene of colluvial debris from a lateral gully (Figure 2a). This material provides a natural barrier, thus sheltering the interior from abrupt thermic fluctuations as it has been reported for other caves in the world (Legge 1972).
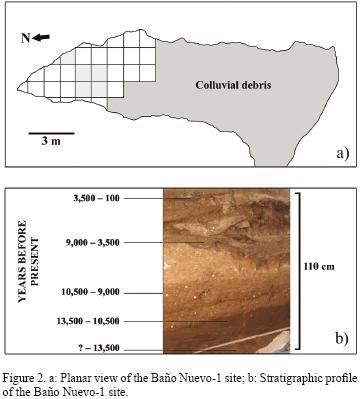
The stratigraphic deposit of the cave has a depth of 150 cm (Figure 2b). From top to bottom, we can first recognize a layer 1 that has very few cultural items, most of them due to perturbation processes such as those that –as we will see– have acted upon bones. Below it, we find several layers that are largely anthropogenic in origin. On the lower part of layer 3, the fragmented remains of ten human skeletons are found (Mena et al., 2003). Below this cultural layer and in a discordant fashion, we find layers 4 and 5 that are formed by rocky debris and collapse, and by organic clay and sand, dating to 10,500–9,000 BP and 10,50013,500 BP, respectively. Layer 4 corresponds to a complex and not yet well understood deposit (it encompasses a 1,500 years hiatus and at least one rock fall and active debris entry, which could have impinged on vertical migration of dermal bones, but not at the site's scale). Layers 5 (and perhaps the lower section of 4 or 4B) are characterized by abundant bones of extinct species and by what seems to be degraded extinct ground sloth feces. Finally, Layer 6 corresponds to stratified sands belonging to a Pleistocene proglacial lake whose original date is unknown (it contacts the rocky foundations of the cave). This layer doesn't have any faunal remains (not even diatoms or other microscopic organisms) and we only know it was deposited before 13,500 BP (Núñez et al., 2005).
DIVERSITY AND TAPHONOMY OF LATE PLEISTOCENE FAUNAL REMAINS
The first mention of extinct fauna at Baño Nuevo–1 was done by Felipe Bate in the 70's, associating in principle cultural evidence such as hearths and lithic artifacts to the bones of American native horse and Xenarhra dermal bones (Bate, 1979). Recent excavations and analyses have recognized a larger scope of extinct fauna, although they have failed to show any association with early humans (Trejo and Jackson, 1998; Mena et al., 2000; Velásquez y Mena, 2006; López, 2009). Within the assemblage, we recognize fossil remains from the families Equidae, Camelidae, Macraucheniidae, Canidae, Ursidae, Megalonychidae, Felidae and the Xenarthra superorder. Megalonychidae remains correspond to as yet unpublished mandible remains and one phalanx, currently under study by E. Bostelmann. He has formerly identified the remains as belonging to Diabolotherium cf. nordenskioldi (Mammalia, Xenarthra, Tardigrada, Megalonychidae), a form up to now recognized outside of Chile, on late Pleistocene sediments from north and south Peru and on central Chubut, southern Argentina (Bostelmann personal communication). This species has been recognized –Albeit not yet published– on three localities in Chile (Bostelmann et al., in preparation). Equid remains are rare, although morphometric analyses suggest that the material from Baño Nuevo–1 can be attributed to Hippidion saldiasi, abundant in late Pleistocene Patagonian deposits (López, 2009). The camelid record consists both of Lama guanicoe and an undetermined Camelidae, larger than present day guanaco and closer in size to genera like Palaeolama and Hemiauchenia or to species named as Lama morphotype L. owenii (Menegaz and Nami, 1999). The scarce Macraucheniidae remains have been identified as Macrauchenia sp. On the other hand, Canidae remains are abundant particularly on layers 4 and 3, and it is worth mentioning that the extinct Dusicyon avus has been directly dated to 7,070 ± 25 AP (Mena et al., 2000. Table 1) and is now known to have subsisted until even later in Central Patagonia (Valentina Trejo, personal observation). Compared to southern Patagonian regions, the faunal record from Baño Nuevo–1 is similar in terms of diversity and taxonomic composition. In fact six out of nine extinct mammals recorded from southern Patagonia have been identified at Baño Nuevo–1. Xenarthra remains from Baño Nuevo–1 (Table 2) have been radiocarbon dated (AMS) between 11,255 ± 30 and 12,510 ± 30 BP (Mena et al., 2000; Velásquez and Mena, 2006), which fall in the middle of the range of dates on Mylodon remains from Última Esperanza (Borrero et al., 1997).
Among the many zooarchaeological and taphonomic analyses from the different layers at Baño Nuevo–1 (Trejo and Jackson, 1998; Núñez et al., 2005; Velásquez and Mena, 2006; Labarca and Lucero, 2007; Mena, 2009), those conducted by López (2009) on the late Pleistocene fossil bones addressed in a general fashion the formation processes that determined their presence at the cave. López (2009: 127) speculated that all these remains were related to carnivores that brought their kills or scavenged carcasses to the site. That zooarchaeological study provided the basic hypotheses and motivation for the more specific taphonomic research now reported. The fossil record from those layers with a larger taxonomic diversity displays a homogeneous distribution of juvenile and adult individuals among herbivore species (Table 3). Among species that occupied open spaces, such as equids, the presence of juveniles (at least two individuals younger than 5 months) is particularly important. On these specimens, as well as on Camelidae bones, traces of carnivore damage were recorded, often on the epiphysiary extremes of long bones. On the basis of their size, shape and localization, these traces were attributed to canid activity (López, 2009), although we cannot rule out the possibility of a large predator, as we discuss below. The finding of extinct ground sloth feces on the oldest layers of the site reveals the intermittent use of the cave by this animal (Mena and Stafford, 2006). As it has been documented that some mylodontines used to dig burrows (Vizcaíno et al., 2001; Dondas et al., 2010) it may well be that they occupied the cave on a seasonal basis. However the finding of only two teeth and no skeletal remains at all indicates that no natural deaths occurred in Baño Nuevo–1. As we have said, the sample is overwhelmingly comprised by dermal bones, a few of which are even found on the late Layer 1.
One of the many problems stemming from these observations is the identification of the agent or agents that accumulated the dermal bones in the cave and how this relates to other extinct fauna (Figures 3 and 4). A second problem is the vertical and horizontal mobility of dermal bones within the cave as this relates to their weight, their size and sphericity (Mena et al., 2000). These variables dont have value in themselves and must necessarily be interpreted in light of their specific archaeological context. The Baño Nuevo–1 site is ideally suited for such an evaluation, since it comprises a basically uninterrupted chronological sequence from the late Pleistocene up to the present, including layers with (Layers 4 and 3) and without (Layers 5 and 1) cultural remains. Here complex formation processes have been identified involving the interplay of several variables such as human occupations, changing surface topographies and the site use by different animal species, among others (Mena et al., 2000; Velásquez and Mena, 2006; López, 2009).
MATERIALS AND METHOD
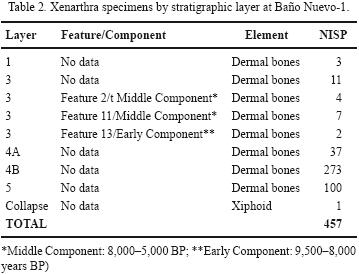
All the analyzed remains (579) come from excavations conducted in early 2006. Although all faunal remains were analyzed, 456 of them (nearly 79%) correspond to dermal bones from Layers 1, 3, 4 and 5 on squares from the back end of the cave (3D, 4D, 5C, 6C, 5B y 6B), the area we would expect for a den or carnivore feeding and bone–chewing spot. Those samples from Layer 3 are reported by the following cultural components: Early (9,500–8,000 BP, Middle (8,000–5,000 BP) and Late (5,000–3,000 BP), as defined on the basis of discrete grass "floors" and other dated features of the units from where the analyzed material comes (Velásquez and Mena, 2006).
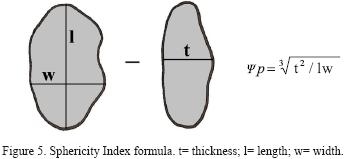
The sample is generally well–preserved and only one specimen (Layer 4) presents exfoliation traces, although the diaganetic processes responsible for this are unknown. The osteometric analysis was based on categories related to each bone's size and sphericity. Each complete dermal bone was measured to its maximum length (l), width (w) and thickness (t), as shown in Figure 5. These values were used to calculate a Sphericity Index (SI), Ψp, according to the formula provided in Figure 5. This parameter measures how close a certain shape is to a perfect sphere (Sneed and Folk, 1958), and its values go from 0 (less spherical) to 1 (more spherical). There are other sphericity analyses particularly common in sedimentology that use form classes (i.e., platy, bladed, elongated) and use them as a basis to compare the thickness–lenght ratio ((l–w)/(l–t) of an spheroid (Sneed and Folk, 1958). This method, however, requires a previous form classification, that in our case is nonexistent.
In order to identify the dermal bones surface alterations and structure, we used a Scanning Electronic Microscope Jeol 5410© enhanced by the Anamaker© software to generate spectroscopic images. This work was conducted at the electronic microscopy lab (Laboratorio de Microscopia Electrónica) of the Metallurgical Engineering Department at the Universidad de Santiago de Chile. The surface of the analyzed bones was subjected to mold making through high precision Silicone type C and replicated with epoxyc resin. Each sample was coated with gold–palladium in order to generate images from the microscope examination.
RESULTS
Size, sphericity and mobility
We have already mentioned that one of the problems to address at Baño Nuevo–1 is the migration of dermal bones from Late Pleistocene strata up to the recent, uppermost layer, an observation that is confirmed by direct radiocarbon dates. On Layer 4, for instance, a extinct ground sloth dermal bone dated to 11,480 ± 50 BP (CAMS–32685) was found above a human made hearth dated to 9,200 ± 80 BP (Beta 90888) (Mena et al., 2000).
Vertical migration could be related to funerary activities, although those recorded at the site do not involve deep digging. It could also be due to sediment relocation during the cleaning of hearths, preparation of grass beddings or other soil disturbing activities that were not clearly recognized. But it seems more likely that large scale displacement of tiny dermal ossicles was not caused by anthropic activities, but by digging and/or burrowing animals, extinct and/or extant. We must also consider the floor relief and the cave's topography, although no stratigraphic reversions were noted anywhere within the deposits and the few incongruencies between absolute depth of the samples and the radiocarbon dates obtained from them are minimal (never contradicting the main stratigraphic sequence) and can easily be explained by irregularities on the surface of the occupation floors. Finally, we must consider collecting of old bones by more recent human occupants of the cave, a phenomenon that has been observed at other Patagonian sites such as Cueva de Los Chingues at Pali Aike (San Román et al., 2000; Martin, 2008) and Arroyo Feo at the Río Pinturas locality, Argentina (Mengoni, personal communication).
Layer 5 and the bottom of Layer 4 are the ones that contain most dermal bones. Layer 4 can be divided into sub–layers –4A and 4B– based on some sedimentological changes and, above all, differences observed on the fossil assemblages: only mylodontine remains were found on 4A, while on 4B faunal diversity is much larger, including unidentified Camelidae, Lama guanicoe, Equidae aff. Hippidion sp., Macrauchenia sp., Felidae, Xenarthra and Diabolotherium cf. nordenskioldi (Table 3). On 4B, 273 dermal ossicles were recovered, while on 4A only 37 were recorded. This difference may suggest that dermal bones found at Layer 4 are due to migration from Layer 5, immediately below, considering absolute dates and the gradual diminution of these remains along Layers 3 and, obviously, in Layer 1.
As Table 4 shows, no difference was observed within the measures and sphericity of dermal bones on different layers. Between Layer 1 and Layer 3 analysis by Studenfs t test indicates no difference between the SI mean (t= 0.78625, P<0.5), which also was no observed between the Layer 3 and Layer 4 (t= 0.40036, P>0.5), and between this layer and Layer 5 (t= –0.85656, P<0.5). As they all tend to be small and semi–spherical, this variable does not seem to be a relevant predictor of mobility nor does it seem to be important with other materials. It is interesting to note, though, that those found on the uppermost layer –although too few– are the heaviest ones, thus indicating that gravity is not a relevant variable either and that there was a force acting from below. Since the analyzed deposits are sheltered from sunlight and the cave does not sustain any kind of vegetation, floralturbation mechanisms such as root and rootlets cannot be considered. Trampling, which is common in restricted spaces with an intense traffic by humans and other heavy animals, such as the Baño Nuevo–1 cave, tends to move artifacts and ecofacts a few centimeters down (Barton, 1982; Villa and Courtin, 1983; Gifford–González et al., 1985; Vermeersch and Bubel, 1997), but not in the opposite direction.
As we have said before, the funerary contexts imply limited sediment movement since adult individuals were placed close to the rocky walls and covered by rocks while the infant corpses seem to have been laid to rest on the surface. On the other hand, their direct dating between 9,000 and 8,500 BP (Mena and Stafford, 2006; Velásquez and Mena, 2006) means that if there were any sediments removal it would have affected early layers and thus, cannot explain the presence of dermal bones on Layer 1. This situation also applies to the hearth cleaning and other human activities even if they went undetected (e.g., post holes) since no dermal bones have been reported from the intervening Layer 2 (3,500–100 BP). Finally, we must consider the collection of old objects and their being carried and used by the site occupants (Schiffer, 1987). However, we would expect these actions to be related to artifact making (for practical or decorative use) which is not the case.
We are left, therefore, with bioturbation or cryoturbation as the only agents to consider for the observed upward migration. There are several site–specific and actualistic studies of animals as agents of stratigraphic disturbances (Langmaid, 1964; Stein, 1983; Erlandson, 1984; Bocek, 1986; McBrearty, 1990; Balek, 2002; Araujo and Marcelino, 2003). At Baño Nuevo–1, the most important stratigraphic disturbance recorded consists of rodent burrows, clearly observed in the lower layers, although it is less clear whether they occurred on the Holocene layers (Figure 6). The study of rodent remains (Labarca and Lucero, 2007) recovered in the excavations suggests that most of them correspond to small Cricetidae that were introduced by predator birds, thus ruling out the importance of in situ habitation or tunnel digging by fossorial caviomorphs, but this analysis refers to only two excavation units. Although their presence and burrowing inside a cave with no grassy cover is not well understood, the presence within the burrows of positively identified skulls of Ctenomys sp. leads us to conclude that this is the organism that dug the observed burrows. This kind of alteration has been observed by Borrero (2003) at Tres Arroyos 1 (Tierra del Fuego, Chile) where rabbit (Oryctolagus cuniculus) bones were collected on the late Pleistocene levels, while this species was just introduced to the island in the 1930's. Since at Baño Nuevo–1 we are dealing with an upward migration, burrowing rodents are a good candidate to be the agents responsible for this movement, since it is well known that these animals can carry sedimentary particles from lower levels, particularly if they are easy to transport due to their size and shape and have a smaller radius than the galleries (Bocek, 1986; Balek, 2002). Thus, although rodent and leporid action has been invoked to explain the downward migration of artifacts and ecofacts in stratified Patagonian archaeological sites (Massone et al., 1993; Borrero, 2003; Martin, 2008), in Baño Nuevo–1 the inverse phenomenon seems to be most common, since on late Pleistocene levels with extinct fauna the presence of cultural materials that may have fallen from above is minimal. Another case of vertical migration is represented at the site by remains of the Felidae that most likely belong to a single individual despite being recorded in the two lowermost contiguous layers.

Cryoturbation or alternate thawing and icing (freezethaw weathering) is another mechanism that may explain the upward migration of ground sloth dermal bones. We know that these processes can indeed transport particles upward (Wood and Johnson, 1978; Schweger, 1985; Schiffer, 1987), but we also know that present day temperature conditions at the cave are fairly stable (between 7 and 12 °C, as recorded daily during the January 2006 – southern summer– field campaign) and are not likely to have promoted this phenomenon. None of the alterations to sediments, bones and artifacts characteristic of these kind of disturbance (Laville et al., 1980; Farrand, 2001) have been recorded. It may never get below 0 °C inside the cave and even if it does, the stability of the thermal conditions could not promote the alternate freeze and thawing that cause vertical migration.
Carnivore alterations
The dermal bones of the Mylodontidae are morphologically similar, as they tend to be round when viewed from above and ovoid in a lateral perspective (Hill, 2006). Their surface is rough to smooth and in some cases it is covered by several small depressions. Osteological studies performed on Glossotherium chapadmalense dermal bones reveal their interior to be smooth and formed by a network of mineralized fibers with different orientations. Close to the surface, these fibers tend to be distributed on orthogonal layers, while a second group of fibers are oriented perpendicular to the bone's main axis. Towards the center, mineralized fibers are less well defined and do not display a clear orientation. This area is formed by small vascular channels limited by concentric bone lamellae (primary osteons) in which we can observe osteocite lagunae. On the bone's periphery, fibers are orientated in a perpendicular fashion to the dermal bone surface that are cut by growth lines parallel to the surface (Hill, 2006). Figures 7a, 7b, 7c and 7d show an extinct ground sloth dermal bone from Baño Nuevo–1 both on its surface and its interior, and it can be observed that the internal fibers are very well–preserved. These characteristics point to the dermal ossicles as very dense units, well–suited to record taphonomic modifications both at the macroscopical and microscopical level.
Despite the integrity of the dermal bones sample from Baño Nuevo–1 and the generally good preservation a few specimens display important external and internal alterations, that can be macroscopically characterized as erosion and perforations of the bony tissue affecting the central core of these pieces from Layers 3, 4 and 5 (Figure 7e). At first, these damages were interpreted as resulting from the action of digestive acids during the passage of ground sloth remains through the digestive tract of large carnivores (López, 2009). This phenomenon is not unusual in Fuego–Patagonian archaeological sites, where alternating occupations by humans and animals of the same sheltered spaces facilitated the preservation of feces from several late–Pleistocene carnivores (Borrero et al., 1997; San Román et al., 2000; Martin, 2008). Damaged dermal bones are usually found isolated, although at Milodon Cave carnivore (Panthera onca mesembrina) feces were recorded in perfect preservation condition containing Mylodon hide fragments and dermal bones (Martin, 2008). This kind of damage by digestive acids had also been reported on eroded surface sites in the semiarid regions of Chile, where small scatters of bone splinters and dermal bones with this kind of damage were explained by the activities of a predator (López and Jackson, 2004; López 2007).
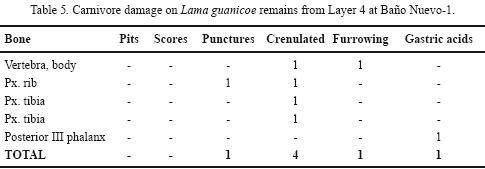
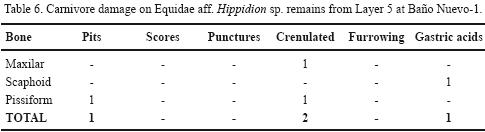
The SEM analysis of a selected dermal bone (NISP= 1) from Baño Nuevo–1 as compared with another one (NISP=1) with no macroscopically observed damage reveals erosion affecting the uppermost bony layer that expresses itself as irregularities and small, homogeneously distributed pits (Figure 7f). As mentioned before, the internal structure of dermal bones is shaped as a net of fibers with different orientations that are distributed on the surface as orthogonal layers. On the altered specimen, the interior is partially removed and the fibers appear corroded and the original structure cannot be observed (Figure 7g). However, the structure of the tissue towards the surface of the damaged dermal does not present major differences with the unaffected control (Figure 7h). We conclude that intense acid corrosion can affect the surface and the interior of dermal bones, while leaving the subsuperficial structure largely intact. These characteristics are similar to those produced by carnivore digestive acids and their low incidence on the sample could reveal little hide consumption by a small to medium sized predator and/or by the age of the predator (Horwitz and Goldberg, 1989; Andrews, 1990; Tappen and Wrangham, 2000). This kind of damage has also been observed on a Lama guanicoe third phalanx from the bottom of Layer 4 and on an Equidae cf. Hippidion sp. radial carpal from Layer 5 (Tables 5 and 6).
Was there consumption of extinct ground sloth by humans at Baño Nuevo–1?
The human consumption of extinct ground sloth has been discussed at length in Patagonian archaeology (Hauthal, 1899; Lehmann–Nitsche, 1899; Emperaire and Laming, 1954; Borrero, 1986, 1997, 2001; Massone, 1987; Mengoni Goñalons, 1987; Borrero et al., 1991; Prieto, 1991; San Román et al., 2000; Tonni et al. 2003; Hajduk et al. 2004; Borrero and Martin, 2008; Martin, 2008), as well as in other parts of the Americas (López, 2007). When dealing with Baño Nuevo –1 this might appear a difficult topic to evaluate, since most of the pieces that can be attributed to this species are dermal bones (there are in addition two teeth and the fragment of a xyphoid). This situation does not differ from the information available from other Patagonian sites, where small bones such as phalanges, carpals, tarsals and dermal bones are dominant (Martin, 2008). The exceptions are represented by Fell Cave, where Mylodon consumption by humans has been documented (Martin 2008). At cave sites in Última Esperanza (i.e., Milodon Cave, Lago Sofía–4) cranial and postcranial remains are fairly abundant, but they may pertain to natural deaths or in situ killing by non–human predators (Martin 2008). The alleged cut marks on mylodontine dermal bones at El Trébol (Río Negro, Argentina) could reflect some use of the animal by humans (Hajduk et al. 2004), although the observed grooves are most likely the results of trampling (F. Mena, personal observation). Macroscopic and microscopic observations on the Baño Nuevo–1 dermal bones were also aimed at identifying anthropogenic marks and one specimen was found to present thin parallel or subparallel grooves on one of its faces. The analysis through SEM, however, reveals that their microtopography does not exhibit the expected traits for traces of a cutting instrument (Shipman, 1981), with a more irregular shape than that expected for them and a less parallel direction. In fact, the subparallel groove distribution and their size (Figures 8a and 8b), suggest they were produced by trampling on abrasive sediments, as has been observed for extinct horse and undetermined mammal remains from Layers 4 and 5. No evidences of fire that could suggest burning of hide or cooking of meat directly on the fire were recorded. Despite these relatively humble and negative results, it is important that researchers use this tool and devote special attention to the analysis of the surface of all materials recovered at an archaeological site, even those often neglected, as are dermal bones.
DISCUSSION
Xenarthra dermal bones tend to be an abundant occurrence at Late Pleistocene/Early Holocene deposits in Fuego–Patagonia. One value of these remains is that these pieces are chronologically diagnostic as the extinction dates for animals of the Mylodon and Glossotherium genera are well established for the Southern Cone (Borrero, 1997). As they are relatively dense particles that provide an adequate surface for recording the imprint of different taphonomic processes they are particularly well suited for investigations of formation processes, but they have been largely ignored while interpreting archaeological and paleontological sites.
As it has been previously discussed, the introduction of dermal bones to sheltered spaces can be the result of different agents (or combinations of agents), including natural in situ deaths, transport by different predators and hunting and consumption by humans. It is particularly important, therefore, to disentangle this complex multicausality and to attempt to interpret the presence of dermal bones in terms of the agent (or agents) responsible for their transport, presence and intersite migration, since they can throw valuable light on the taphonomic history of a deposit. Since these small and fairly ubiquitous particles are particularly prone to vertical migration, their study can also provide information about stratigraphic integrity in the absence of more evident or large scale disturbances.
Baño Nuevo–1 provides and example of the utility of dermal ossicles in stratigraphic analysis. As if it were not enough to know that the bearers of the dermal bones were extinct by ten thousand years ago, many of them have been directly dated to the 12th millennium BP but they are nonetheless found in minor quantities on layers that correspond to the last century. One dermal bone was even dated to 11,480±50 BP over a hearth dated to 9,200±80 BP (Mena et al., 2000). We have attempted to explain this migration both in terms of the bone's dimensions and shape and in terms of the transport agents, most likely burrowing rodents. Vertical migration can also occur with other small and compact bones such as carpals and tarsals, although this has not been observed at Baño Nuevo–1, given the absence of these elements from other extinct taxa on Holocene levels.
As suspected by a preliminary inspection and confirmed by Electronic Scanning Microscopy, some of these bones were part of the feces of a carnivore. This observation raises the question about the carnivore or carnivores that preyed on mylodontines near the cave and that occupied it before the arrival of humans on the Early Holocene. On the site, two potential candidates for moving and/or chewing extinct ground sloth hides are canids, Arctotherium and a species similar to Panthera onca mesembrina (Trejo and Jackson, 1998; López, 2009). No remains of Dusicyon avus have been found in the Pleistocene layers at the site (Valentina Trejo, personal observation), but is most likely that they were there. From the bone evidence of other late Pleistocene species at the site (i.e., Camelidae, Lama guanicoe, Equidae) we observe fairly frequent damage marks, including pits, scores, punctures and furrowing (Tables 5 and 6), of a size closer to those produced by canids than those left by larger predators such as Panthera onca mesembrina or Arctotherium (Martin, 2008). However, feline and bears marks are not very abundant and tend to be localized on specific places of the bones (Martin, 2008). In those cases in which the age of the herbivore taxa found on the Pleistocene layers could be detected, we find a large proportion of juvenile horse and guanaco, including –as we said before– two equid individuals ranging in age from 1 to 5 months (see Table 3). Big animals such as extinct ground sloth and Macrauchenia are only represented by small non skeletal elements such as dermal bones and teeth. This record hence suggests that the accumulating agent at Baño Nuevo–1 could not hunt and transport to the cave large prey but it was effective in collecting and destroying bones of animals between 100 and 300 kg. The role of Dusicyon avus and/or other canids on the formation of the paleontological context must be reevaluated as they could have hunted smaller species and/or scavenged those prey hunted and consumed by larger predators like Panthera onca mesembrina and Arctotherium. The presence of ground sloth feces point to the effective occupation of the cave by this extinct edentate, and it is possible that different carnivores also dwelled in the site, although in alternating fashion. As it has been observed at den sites like Lago Sofía 4, where a larger diversity of Mylodon elements are found, carnivores selected specific bones with meat attached and the hide may have been abandoned at the kill, an argument that would also explain the scarcity of dermal bones with digestive acid traces at Cueva de Los Chingues (Pali Aike region) and other sites (Martin, 2008). Unlike most Patagonian sites, however, Baño Nuevo–1 does not exhibit a clear pattern of damage by large felines or bears, and the size of the observed punctures appear more like those produced by small to middle–sized carnivores like foxes.
Another possible scenario is related to the introduction by humans of ground sloth hides adhered to meat for consumption inside the cave. As we have seen, however, no evidence suggests anthropic action on these animals and/or their bones. In this respect, the Baño Nuevo–1 site reveals itself as a particularly strong case against the coexistence and contemporaneity of Pleistocene species and human beings, since there are the well dates pre–human sediments with a good fossil record of many extinct species, and if there was any human activity there it should have left some evidence. This might be a strictly local phenomenon and does not grant us permission to generalize and to negate, for instance, the role of human pressure on the extinction of these species.
Just as the present taphonomical research stems from largely "intuitive" or "heuristic" observations and questions in the framework of a first zooarchaeological approach to the Pleistocene assemblage collection, we are certain that these observations have reduced the alternative hypotheses and have provided a better focus to questions to be addressed by future research. Dermal bones had not been the specific focus of observation before and we have learned that in the future more specific data must be collected in the field and more analytical analyses are due (i.e., definition of sedimentological and/or microscopic indicators of cryoturbation and argilliturbation). Since dermal bones are –as we have said– particularly well suited for formation processes research (abundant, dense, good imprinting surfaces), it was important to initiate their thorough study, although a more complete interpretation in the future must consider other contextual variables. Since no final definitive conclusions exist in science, if we were only to report "success stories" and/or sensational results we would fail to provide a feeling of the long enquiry that is any scientific research, which all too often doesn't lead us directly to the expected results. The challenge for future research is precisely to tread these unpredictable paths and to build new knowledge while applying information already obtained to the analysis of those sites where human consumption of extinct ground sloth is claimed.
ACKNOWLEDGEMENTS
Our thanks to Emily Lindsey for reading and comment on an earlier version of this work. We also thank Gladys Olivares (Laboratorio de Microscopia Electrónica, USACH) for their help and Enrique Bostelmann. We also want to thank Dr. Gustavo Juan Scillato–Yané for his comments to a previous version of this manuscript, and to a second anonymous reviewer. Research financed by FONDECYT 1090027 Project, Ser humano y contexto de la ocupación inicial de la estepa del norte y centro de Aisén (transición Pleistoceno terminal /Holoceno): estrategias de búsqueda e interpretación del registro arqueológico temprano.
REFERENCES
Andrews P., 1990, Owls, caves and fossils: Chicago, University of Chicago Press, 239 pp. [ Links ]
Araujo, A.G., Marcelino, J.C., 2003, The role of armadillos in the movement of archaeological materials: an experimental approach: Geoarchaeology, 18, 433–460. [ Links ]
Balek, C.L., 2002, Buried artifacts in stable upland sites and the role of bioturbation: a review: Geoarchaeology, 17, 41–51. [ Links ]
Barton, N.E., 1982, Vertical distribution of artefacts and some post–depositacional factor affecting site formation, in Rowley–Conwy, P., Zvelebil M., Blankholm, P. (eds), Mesolithic Northwest Europe: Recent Trends: Sheffield. University of Sheffield, 55–62. [ Links ]
Bate, L., 1979, Las investigaciones sobre los cazadores tempranos en Chile austral: Trapananda, 1(2), 150–162. [ Links ]
Bocek, B., 1986, Rodent ecology and burrowing behaviorpredicted effects on archaeological site formation: American Antiquity, 51, 589–603. [ Links ]
Borrero, L.A., 1986, Cazadores de Mylodon en la Patagonia austral, in Bryan, A. (ed.), New Evidence for the Pleistocene Peopling of the Americas: Orono, Center for the Study of Early Man, 281–284. [ Links ]
Borrero, L.A., 1997, La extinción de la megafauna en la Patagonia: Anales del Instituto de la Patagonia, 25, 89–102. [ Links ]
Borrero, L.A., 2001, El poblamiento de la Patagonia Toldos, milodones y volcanes: Buenos Aires, Emecé, 195 pp. [ Links ]
Borrero, L.A., 2003, Taphonomy of the Tres Arroyos 1 rockshelter, Tierra del Fuego, Chile. Quaternary International, 109–110, 87–93. [ Links ]
Borrero, L.A., Martin, F., 2008, A reinterpretation of the Pleistocene human and faunal association at Las Buitreras Cave, Santa Cruz, Argentina: Quaternary Science Reviews, 27, 2509–2515. [ Links ]
Borrero, L.A., Lanata, J.L., Cárdenas, P., 1991, Reestudiando cuevas: nuevas excavaciones en Ultima Esperanza, Magallanes: Anales del Instituto de la Patagonia, 20, 101–110. [ Links ]
Borrero, L.A., Martin, F.M., Prieto, A., 1997, La cueva Lago Sofía 4, Ultima Esperanza: una madriguera de felino del Pleistoceno tardío: Anales del Instituto de la Patagonia (Serie Ciencias Sociales), 25, 103–122. [ Links ]
Carlini A., Vizcaíno S., Scillato–Yané, G., 1997, Armored xenarthrans: A unique taxonomic and ecologic assemblage, in Kay R., Madden R., Cifelli R., Flynn J. (eds.), The Miocene Fauna of La Venta, Colombia: Washington, DC, Smithsonian Institution Press, 213–226. [ Links ]
Chávez Aponte, E., Hernández, I.A., Finol, H. J., Barrios, C., Boada, A., Carrillo, J., 2008, Histología y ultraestructura de los osteodermos fósiles de Glyptodon clavipes y Holmesina sp. (Xeanrthra: Cingulata): Interciencia, 33, 616–619. [ Links ]
Dondas, A., Isla, F., Carballido, J.L., 2010, Paleocaves exhumed from the Miramar Formation (Ensenadan Stage–age, Pleistocene), Mar del Plata, Argentina: Quaternary International, 210 (1–2), 44–50. [ Links ]
Emperaire, J., Laming, A., 1954, La grotte du Mylodon (Patagonie Occidentale): Journal de la Société des Américanistes Nouvelle Série, XLIII, 173–205. [ Links ]
Erlandson, J.M., 1984, A case–study in faunalturbation–delineating the effects of the burrowing–pocket–gopher on the distribution of archaeological materials: American Antiquity, 49, 785–790. [ Links ]
Farrand, W.R., 2001, Archaeological Sediments in Rockshelters and Caves, in Stein, J. K. and Farrand, W. R., Sediments in Archaeological Context: Salt Lake City, The University of Utah Press, 29–66. [ Links ]
Gifford–González, D., Damrosch, D., Damrosch, D., Pryor, J., Thunen, R., 1985, The third dimension in site structure: an experiment in trampling and vertical displacement: American Antiquity, 50, 803–818. [ Links ]
Hajduk, A., Albornoz, A.M., Lezcano, M.J., 2004, El "Mylodon" en el patio de atrás. Informe preliminar sobre los trabajos en el sitio El Trébol, ejido urbano de San Carlos de Bariloche, Provincia de Río Negro, in Civalero, M., Fernández, P., Guráeib, A.G. (eds.), Contra Viento y Marea. Arqueología de Patagonia: Buenos Aires, Instituto Nacional de Antropología y Pensamiento Latinoamericano/ Sociedad Argentina de Antropología, 715–131. [ Links ]
Hauthal, R., 1899, Reseña de los hallazgos en las cavernas de Última Esperanza (Patagonia austral): Revista del Museo de La Plata, IX, 411–420. [ Links ]
Hill, R., 2006, Comparative anatomy and histology of xenarhran osteoderms: Journal of Morphology, 267, 1441–1460. [ Links ]
Horwitz, L.K., Goldberg, P., 1989, A study of Pleistocene and Holocene hyaena coprolites: Journal of Archaeological Science, 16, 71–94. [ Links ]
Krmpotic, C., Ciancio, M., Barbeito, C., Mario, R., Carlini, A., 2009, Osteoderm Morphology in Recent and Fossil Euphractinae Xenarthrans: Acta Zoologica, 90, 339–351. [ Links ]
Labarca, R., Lucero, V., 2007, Tafonomía de micromamíferos y procesos de formación en la Cueva Baño Nuevo (XI Región, Chile): Actas de las VI Jornadas de Arqueología de la Patagonia, 391–402. [ Links ]
Langmaid, K.K. ,1964, Some effects of earthworm invasion in virgin podzols: Canadian Journal of Soil Science, 44, 34–37. [ Links ]
Latorre, C. 1998, Paleontología de mamíferos del alero Tres Arroyos I, Tierra del Fuego, XII Región, Chile: Anales del Instituto de la Patagonia (Serie Ciencias Naturales), 26, 77–90. [ Links ]
Laville, H., Rigaud, J.P., Sackett, J., 1980, Rock Shelters of the Perigord. Geological Stratigraphy and Archaeological Succession: New York, Academic Press, 371 p. [ Links ]
Legge, Anthony, 1972, Cave climates, in Higgs, E. (ed.), Papers in Economic Prehistory: Cambridge, Cambridge University Press, 97–103. [ Links ]
Lehmann–Nitsche, R., 1899, Coexistencia del hombre con un gran desdentado y un equino en las cavernas patagónicas: Revista del Museo de La Plata, IX, 455–472. [ Links ]
López, P., 2007, Tafonomía de los mamíferos extintos del pleistoceno tardío de la costa meridional del semiárido de Chile (IV Región–32° Latitud S). Alcances culturales y paleoecológicos: Chungará, 39(1), 69–86. [ Links ]
López, P., 2009, El mundo perdido de Patagonia Central: una aproximación tafonómica al estudio de los mamíferos extintos del sitio Baño Nuevo–1 (XI Región–Chile), in López, P., Cartajena, I., García, C., Mena, F. (eds.), Zooarqueología y Tafonomía en el Confín del Mundo: Santiago, Ediciones Universidad Internacional SEK–Chile, 181–198. [ Links ]
López, P., Jackson, D., 2004, Ground Sloth Predation in the Northern Semiarid Region of Chile: Current Research in the Pleistocene, 21, 14–16. [ Links ]
Martin, F., 2008, Bone Crunching felids at the End of the Pleistocene in Fuego–Patagonia, Chile: Journal of Taphonomy, 6(3–4), 337–372. [ Links ]
Martin, F.M., Borrero, L.A., 1997, A Puma lair in southern Patagonia: implications for the archaeological record: Current Anthropology, 38, 453–461. [ Links ]
Massone, M., 1987, Los cazadores paleoindios de Tres Arroyos (Tierra del Fuego): Anales del Instituto de la Patagonia (Serie Ciencias Sociales), 17, 47–60. [ Links ]
Massone, M., Jackson, D., Prieto, A., 1993, Perspectiva arqueológica de los Selk'nam: Santiago, Centro de Investigaciones Diego Barros Arana, 170 pp. [ Links ]
McBrearty, S., 1990, Consider the humble termite: Termites as agents of post–depositional disturbance at African archaeological sites: Journal of Archaeological Science, 17, 111–143. [ Links ]
Mena, F., 2009, Aves en Cueva Baño Nuevo, in López, P., Cartajena, I., García, C., Mena, F. (eds.), Zooarqueología y Tafonomía en el Confín del Mundo: Santiago, Ediciones Universidad Internacional SEK–Chile, 59–71. [ Links ]
Mena, F., Stafford, T., 2006, Contexto estratigráfico y fechación directa de esqueletos humanos del Holoceno Temprano en Cueva Baño Nuevo (Patagonia Centra, Chile), in Jiménez, J. (ed.), Segundo Simposio Internacional el Hombre Temprano en América: México, Instituto Nacional de Antropología e Historia, 139–154. [ Links ]
Mena, F., Lucero, V., Reyes, O., Trejo, V., Velásquez, H., 2000, Cazadores tempranos y tardíos en la Cueva Baño Nuevo–1, margen occidental de la estepa centropatagónica (XI Región de Aysén; Chile): Anales del Instituto de la Patagonia, 28, 173–195. [ Links ]
Mena, F., Reyes, O., Stafford, T., Southon, J., 2003, Early human remains from Baño Nuevo–1 cave, Central Patagonian Andes, Chile: Quaternary International, 109–110, 113–121. [ Links ]
Menegaz, A., Nami, H.G., 1999, Nota sobre los hallazgos paleontológicos de la fauna del Pleistoceno final en la cueva de Las Buitreras (Provincia de Santa Cruz, Argentina): Praehistoria, 3, 183–187. [ Links ]
Mengoni Goñalons, G., 1987, Modificaciones culturales y animales en los huesos de los niveles inferiores del sitio Tres Arroyos 1 (Tierra del Fuego, Chile): Anales del Instituto de la Patagonia (Serie Ciencias Sociales), 17, 61–66. [ Links ]
Núñez, H., Stafford, T., Frassinetti, D., 2005, Primer registro de fósiles de Liolaemus en Chile (Reptilia, Sauria): Noticiario Mensual del Museo de Historia Natural, 356, 3–7. [ Links ]
Prieto, A., 1991, Cazadores Tempranos y Tardíos en Cueva del Lago Sofía 1: Anales del Instituto de la Patagonia (Serie Ciencias Sociales), 20, 75–99. [ Links ]
San Román, M., Morello, F., Prieto, A., 2000, Cueva de Los Chingues (Parque Nacional Pali–Aike), Magallanes, Chile. Historia natural y cultural I: Anales del Instituto de la Patagonia (Serie Ciencias Humanas), 28, 125–143. [ Links ]
Schweger, C., 1985, Geoarchaeology of northern regions: lessons from cryoturbation. in Stein, J.K., Farrand, W.R. (eds.) Archaeological Sediments in Context: Orono, Center for the Study of Early Man, 127–141. [ Links ]
Schiffer, M.B., 1987, Formation processes of the archaeological record: Albuquerque, University of New Mexico Press, 428 p. [ Links ]
Shipman, P., 1981, Applications of scanning electron microscopy to taphonomic problems: Annals of the New York Academy of Sciences, 376, 357–385. [ Links ]
Sneed, E., Folk, R., 1958, Pebbles in the lower Colorado River, Texas a study in particle morphogenesis: The Journal of Geology, 66(2), 114–150. [ Links ]
Stein, J.K., 1983, Earthworm activity: A source of potential disturbance of archaeological sediments: American Antiquity, 48, 277–289. [ Links ]
Tappen, M., Wrangham, R., 2000, Recognizing Hominoid–Modified Bones: The Taphonomy of Colobus Bones Partially Digested by Free–Ranging Chimpanzees in the Kibale Forest, Uganda: American Journal of Physical Anthropology, 113, 217–234. [ Links ]
Tonni, E.P., Carlini, A.A., Scillato–Yané, G.J. y Figini, A.J., 2003. Cronología radiocarbónica y condiciones climáticas en la "Cueva del Milodón" (sur de Chile) durante el Pleistoceno Tardío. Ameghiniana 40 (4): 609–615. [ Links ]
Trejo, V. , Jackson, D., 1998, Cánidos patagónicos: identificación taxonómica de mandíbulas y molares del sitio arqueológico cueva Baño Nevo–1 (Alto Ñirehuao, XI Region): Anales del Instituto de la Patagonia, 26, 181–194. [ Links ]
Velásquez, H., Mena, F., 2006, Distribuciones óseas de ungulados en la Cueva Baño Nuevo–1 /XI Region, Chile): un primer acercamiento: Magallania, 34(2), 91–106. [ Links ]
Vermeersch, P., Bubel, S., 1997, Postdepositational artifact scattering in a podzol. Processes and consequences for late Paleolithic and Mesolithic: Anthropologie, 35(2), 45–56. [ Links ]
Vickaryous, M., Hall, B., 2006, Osteoderm morphology and development in the nine–banded armadillo, Dasypus novemcinctus (Mammalia, Xenarthra, Cingulata): Journal of Morphology, 267, 1273–1283. [ Links ]
Villa, P., Courtin, J.,1983, The interpretation of stratified sites: a view from underground. Journal of Archaeological Science, 10, 267–281. [ Links ]
Vizcaíno, S., Zárate, M., Bargó, S., Dondas, A., 2001, Pleistocene burrows in the Mar del Plata area (Argentina) and their probable builders: Acta Paleontológica Polonica, 46 (2), 289–301. [ Links ]
Wood, R.W., Johnson, D.L., 1978, A Survey of Disturbance Processes in Archeological Site Formation, in Schiffer M.B. (ed.). Advances in Archaeological Method and Theory, New York: Academic Press, 315–381. [ Links ]

