










Servicios Personalizados
Revista
Articulo
 Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista mexicana de ciencias geológicas
versión On-line ISSN 2007-2902versión impresa ISSN 1026-8774
Rev. mex. cienc. geol vol.28 no.3 Ciudad de México dic. 2011
Late Cretaceous nodosaurids (Ankylosauria: Ornithischia) from México
Nodosáuridos (Ankylosauria: Ornithischia) del Cretácico Tardío de México
Héctor E. Rivera–Sylva1*, Kenneth Carpenter2,3, and Francisco Javier Aranda–Manteca4
1 Departamento de Paleontología, Museo del Desierto, Carlos Abedrop Dávila 3745, 25022, Saltillo, Coahuila, Mexico.*hrivera@museodeldesierto.org.
2 Prehistoric Museum, Utah State University – College of Eastern Utah, 155 East main Street, Price, 84501 Utah, USA.
3 University of Colorado Museum, 80309 Boulder, Colorado, USA.
4 Laboratorio de Paleontología, Facultad de Ciencias Marinas, Universidad Autónoma de Baja California, Ensenada, Baja California, Mexico.
Manuscript received: February 15, 2011
Corrected manuscript received: April 15, 2011
Manuscript accepted: May 9, 2011
ABSTRACT
Nodosaurid ankylosaur remains from the Upper Cretaceous of Mexico are summarized. The specimens are from the El Gallo Formation of Baja California, the Pen and Aguja Formations of northwestern Coahuila, and the Cerro del Pueblo Formation of southeast Coahuila, Mexico. These specimens show differences from other known nodosaurids, including an ulna with a well developed olecranon and prominent humeral notch, the distal end of the femur not flaring to the extent seen in other nodosaurids, and a horn–like spine with vascular grooves on one side. The specimens represent the southernmost occurrences of nodosaur remains in North America, and provide an important biogeographical link between nodosaurids of the United States and Canada on the one hand, and Argentina and Antarctica on the other.
Key words: nodosaurid, Ankylosauria, Late Cretaceous, Baja California, Coahuila, Mexico.
RESUMEN
Restos de anquilosaurios nodosáuridos del Cretácico Superior de México son descritos aquí. Las muestras proceden de la Formación El Gallo de Baja California, de las formaciones Pen y Aguja del noroeste de Coahuila, y de la Formación Cerro del Pueblo, sureste de Coahuila, México. Estas muestras presentaron diferencias significativas con otros nodosáuridos conocidos, incluyendo el cúbito con olécranon bien desarrollado y la muesca del húmero prominente, el extremo distal delfémur de diferente medida no visto en otros nodosáuridos, y una espina similar a un cuerno con los surcos vasculares en un lado. Las muestras son importantes porque son los hallazgos más meridionales de América del Norte, y proporcionan un vínculo biogeográfico importante entre los nodosáuridos de los Estados Unidos y Canadá, por un lado, y Argentina y la Antártida por el otro.
Palabras clave: nodosáurido, Anquilosaurio, Cretácico Tardío, Baja California, Coahuila, México.
INTRODUCTION
Nodosaurid ankylosaurs form an important component of the Late Cretaceous dinosaur fauna in the northern hemisphere, especially Europe, the United States, and Canada (Coombs, 1978; Carpenter, 2001). Recently, how–ever, nodosaurids have been reported from the southern hemisphere, including Argentina (Coria and Salgado, 2001) and Antarctica (Gasparini et al., 1987; 1996). Crucial to understanding the distribution of these ankylosaurs are specimens from Mexico.
In 1971, Harley J. Garbani found the first evidence of an ankylosaur from Baja California, an osteoderm from El Gallo Formation (Campanian). The specimen was only mentioned by Morris (1967) and the osteoderm was held in the Los Angeles County Museum. Later it was transferred to the Colección Nacional de Paleontología in the Universidad Nacional Autónoma de México (UNAM). Unfortunately, the specimen cannot be located (Perrilliat, pers. com., 2009), thus its identity as nodosaurid or ankylosaurid remains unknown. A new specimen of ankylosaur from Baja is a tooth from the El Jabón creek locality, El Disecado member, El Gallo Formation (Campanian), Baja California, Mexico. In 2006, Rivera–Sylva and Espinosa–Chávez reported the presence of ankylosaur osteoderms from the Cerro del Pueblo Formation in Coahuila, Mexico. Since then, additional undescribed ankylosaur remains have been found in the Upper Cretaceous of Mexico (Meyer et al., 2005), including two vertebrae and a metatarsal from the San Carlos Formation in Aldama, Chihuahua (Figure 1). We describe some of this new material here and highlight diíferences with specimens from contemporaneous formations of the northern United States and southern Canada. These diíferences suggest that the mexican nodosaurids represent at least one new taxon, but at present we refrain from naming it owing to the fragmentary nature of the material.
The abbreviations used in this study are as follows: AMNH = American Museum of Natural History, New York, New York, USA; CPC = Colección Paleontológica de Coahuila, Museo del Desierto, Saltillo, Coahuila, Mexico; UABC FCM = Universidad Autónoma de Baja California Facultad de Ciencias Marinas, Ensenada, Baja California, Mexico.
SYSTEMATIC PALAEONTOLOGY
Suborder Thyreophora Nopcsa, 1915
Infraorder Ankylosauria Osborn, 1923
Family Nodosauridae Marsh, 1890
Nodosauridae indet.
Specimen A
Material examined. UABC FCM 2625, one tooth crown (Figure 2).
Occurrence. The material was recovered in grey mudstone from El Disecado Member of the El Gallo Formation (Campanian), El Jabón ravine, Baja California, Mexico (N30°03'14"; W115°45'44") (Figure 1). Description. The specimen consists of a tooth crown lacking the root. It most closely resembles those of other nodosaurids in its relatively large size, presence of a complete shelf–like cingulum, and low denticle count along the margins (6 anteriorly and 3 posteriorly). The cingulum is irregular on the labial side, whereas on the lingual side it is smooth and concave.
Teeth are taxonomically useful at the family level in ankylosaurs (Coombs and Maryanska, 1990). Tooth crowns of nodosaurids are proportionally large, with few denticles (4–8) along the margins that in most cases extend onto the crown face as ridges (Coombs, 1990). The base of the crown is swollen and a complete cingulum is present on one or both sides. In contrast, the teeth of polacanthids and anky–losaurids are small (Carpenter, pers. obs., 2001).
Specimen B
Material examined. CPC 272, one dorsal vertebra, one caudal vertebra, distal end of humerus, partial ulna, distal end of femur, one rib fragment, and one osteodermal spine (Figure 3).
Occurrence. The material was recovered from the shale of the marine member of the Pen Formation (Campanian), south of San Miguel, Coahuila, Mexico (N28°38'16"; W102°56'55") (Figure 1).
Description. The dorsal vertebra consists primarily of the centrum (Figure 3e–3h). It has a typical ankylosaurian morphology, being amphiplatyan, spool–shaped and constricted in the middle; it is longer and taller than wide. A portion of one peduncle is present, but otherwise the neural arch and spine are missing. The caudal vertebra consists only of a damaged centrum, with the neural arch and spine missing. The centrum is anteroposteriorly shorter than wide, and somewhat hexagonal in proximal view, as in many nodosaurids (Carpenter, pers. obs.).
The left humerus consists of the distal end (Figure 3a–3d). The incomplete shaft is mediolaterally elliptical in cross–section. The distal condyles are well preserved, and both are about equal in size, although the medial condyle extends ventrally beyond the lateral condyle. In contrast, in both Edmontonia and Panoplosaurus the medial condyle is smaller than the lateral condyle and does not extend ventrally very much beyond the lateral condyle.
The left ulna lacks the distal third (Figure 3i–3l). The olecranon is most similar to that of Stegopelta (Carpenter and Kirkland, 1998) in that it is a prominent, nearly parallel–sided form in lateral view and is truncated proximally; this is in contrast to the more usual tapering or conical process in many ankylosaurs (Carpenter, pers. obs). In anterior view, the olecranon resembles that of most ankylosaurs, being a broad, inverted right triangle. However, the proximal end is rounded, rather than tapered as in Sauropelta. The humeral notch at the base of the olecranon is well developed, whereas the laterally placed radial notch is shallow as is typical of ankylosaurs (Carpenter, pers. obs).
The left femur is represented by the distal third. The intercondylar groove is shallow. The distal end is slightly wider than the shaft (Figure 3), whereas in Edmontonia, the distal end flares significantly wider than the shaft (the femur is unknown in Panoplosaurus). Posteriorly, the lateral condylar ridge (sometimes referred to as the fibular process), is square and prominent.
A long, tapered, slightly curved "horn"–like structure is probably a distal thoracic osteoderm that is missing its base (Figure 3m–3p). In cross–section, the osteoderm is elliptical, being deeper than wide. One side (anterior?) has vascular grooves extending the length of the bone, thus somewhat resembling a ceratopsian horn–core.
Specimen C
Material examined. CPC 273 consists of two caudal vertebra (CPC 273/1; CPC 273/2), one cervical rib (CPC 273/3), six rib fragments (CPC 273/4), a posterior fragment of ilium (CPC 273/5), proximal fragment of ?tibia (CPC 273/6), proximal end of ?femur (CPC 273/7); thirteen osteoderms (CPC 273/9; CPC 273/10; CPC 273/11; CPC 273/12; CPC 273/13; CPC 273/14; CPC 273/15; CPC 273/16; CPC 273/17; CPC 273/18; CPC 273/19; CPC 273/20; CPC 273/21; CPC 273/22), proximal end of scapula (CPC 273/23), and twenty seven undetermined bone fragments (CPC 273/24) (Figure 4, Table 1).
Occurrence. The material was recovered in sandy mudstone layers from the the upper unnamed member of the Aguja Formation (Campanian), west of the El Carricito, township of Ocampo, Coahuila, Mexico (N28°55'17"; W103°19'72") (Figure 1).
Description. The vertebrae and the tibia were collected from the same site as the osteoderms and these are therefore assumed to belong to the same individual.
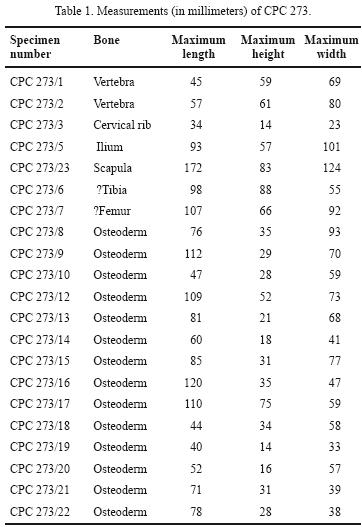
Vertebrae. CPC 273/1 is a caudal vertebra, bearing facets for rib articulation. The neural canal is circular and broken dorsally. The neural spine is broken. The ventral surface is weathered. In contrast, CPC 273/2 has a short transverse process. Most of the surface of the centrum is eroded; there is a foramen on its ventral surface.
Tibia. A partial right tibia is represented by the proximal end. It has a short cnemial crest located near the medial edge. The lateral side is missing, thus the relative width versus length of the tibial plateau cannot be determined.
Ilium. A rounded fragment may be the postacetabular process of the right ilium. The dorsal surface is slightly concave, whereas the ventral side has a rounded reinforcing ridge or buttress that extends the length of the fragment. Its distal termination is blunt and may have been the origin of the M. caudofemoralis brevis. This ridge is offset towards the medial half. The preserved portion is too incomplete for comparison with other ilia.
Scapula. The scapula is represented by the scapular margin of the glenoid cavity. The angle of the glenoid suggests it is a left scapula. Unfortunately, it does not preserve the dorsal margin or acromion process.
Osteoderms. CPC 273/9 (Figure 4g) is large and rectangular, with a median keel that is taller posteriorly and terminates in a conical point near the osteoderm's posterior margin; its lateral margin is broken. CPC 273/10 and CPC 273/11 have the same anatomical features as CPC 273/9 but both are broken in their middles. CPC 273/12 is a conical anterolaterally–directed cervical spine (Figure 4i). CPC 273/13, CPC 273/14, and CPC 273/15 are circular with a low keel and are of the type found in the thoracic region of Edmontonia (Carpenter, 1990) (Figure 4a–4c). CPC 273/16 is a distal thoracic osteoderm. CPC 273/17 is a caudal osteoderm. CPC 273/19 is a circular ossicle (Figure 4e). The superficial surface is flat; the deep surface is flat to slightly concave. The superficial surface is pitted, grooved, and rugose. The margin is rounded.
CPC 273 can be attributed to the Nodosauridae based on the anterolaterally–directed distal cervical spine and the keeled thoracic osteoderms that are slightly excavated on their deep surface (Vickaryous et al., 2004; Lee, 1996; Sereno, 1986). The superficial surface texture of the osteoderms most closely matches that reported by Burns (2008) for Glyptodontopelta in that the surface is generally smooth, but there are a few randomly distributed small pits and vascular grooves that radiate from the keel. These differ, however, in that the pitting is less extensive.
Specimen D
Material examined. CPC 275, caudal vertebra.
Occurrence. The material was recovered from sandstone layers from the Cerro del Pueblo Formation (Campanian), north of Saltillo, Coahuila, Mexico (N26°10'48"; W101°16'48"). (Figure 1).
Description. CPC 275 is a caudal vertebra eroded on most of its surface. It consists of a centrum lacking most of the neural arch and spine, and the transverse process. The centrum is short anteroposteriorly and has a pair of chevron facets on the posteroventral edge. In posterior view, the centrum has the characteristic hexagonal shape of a nodosaurid.
DISCUSSION
The tooth (UABC FCM 2625) is similar to that of Aletopelta coombsi (Ford and Kirkland, 2001) from the Upper Cretaceous of southern California. Ford and Kirkland (2001) consider Aletopelta to be an ankylosaurid and not a nodosaurid as originally described by Coombs and Demere (1996). However, Aletopelta also has nodosaurid characters including tooth morphology and osteoderms that are thicker than those of typical ankylosaurids (Carpenter, pers. obs.).
The specimen CPC 273 is a nodosaurid because the osteoderms are thick and have a slightly concave base. CPC 272 is also identified as a nodosaurid primarily because of the elongate osteodermal spine.
Although four nodosaurids have thus far been identified from the Upper Cretaceous of North America (Panoplosaurus, Edmontonia, Glyptodontopelta, and Aletopelta), specimen B from the Pen Formation suggests the presence of a fifth genus (Table 2). However, the material is too fragmentary to be named.
Osteoderms represent the most abundant material among the Mexican specimens. CPC 273/12 is similar to the distal osteoderm of the first cervical ring (Sternberg, 1928), somewhat like that projecting anterolaterally from the cervical half rings of AMNH 5665 (Edmontonia; Matthew, 1922; Carpenter, 1990). The oval, low keeled osteoderm is similar to the lateral cervical osteoderms illustrated for Edmontonia (Carpenter, 1997), CPC 273, and AMNH 5665. However, the surface texture is different from Edmontonia (Table 2) in that it has pitting and a dentritic pattern radiating from the keel (Burns, 2008).
Lehman (1997) estimated that ankylosaurs comprised only 8% of the dinosaur fauna in the nearby Aguja Formation in the Big Bend region of Texas. Based on North American localities of ankylosaur tracksites (McCrea et al., 2001) and the presence of aquatic vertebrates associated with ankylosaur bones and teeth, ankylosaurs probably preferred humid environments (Vickaryous et al., 2004).
However, many Asian ankylosaurs lived in arid or semi–arid environments (Carpenter, 1997) and their rarity in the Aguja Formation and other coastal and floodplain settings, such as the Cerro del Pueblo Formation, may indicate that they were more common in inland areas that are not well documented in the North American fossil record.
CONCLUSIONS
The discovery of nodosaurid remains in the Upper Cretaceous of Mexico is not unexpected. These are the southernmost occurrences of the family in North America. Two genera of nodosaurid ankylosaurs have been reported from the Campanian Aguja Formation of Texas, Panoplosaurus sp. and Edmontonia sp. (Lehman, 1987; Carpenter, 1990; Weishampel et al., 2004; Vickaryous et al., 2004; Sankey, 2010). The remains from Mexico may represent one, both, or a new genus. CPC 275 also represents the first record of a nodosaurid from the Cerro del Pueblo Formation.
ACKNOWLEDGEMENTS
We are grateful to the following individuals for their contribution to this project: José Manuel Padilla, Abraham Isaí Oyervidez Salazar, and José López, from the Museo del Desierto. Ileana de la Peña Oviedo, Jorge A. Ortíz Mendieta, Rubén Guzmán Gutiérrez (SMA), Elizabeth Jardón, Tatiana Sánchez Rodríguez (MHN), and Homero Hernández helped during field seasons. The Museo del Desierto is also thanked for supporting this investigation, and therefore we appreciate Arturo H. González González for his support of this project. We are obliged to thank Daniel Posada Martínez (MUDE) for the donation of Specimen D. In the same manner we wish to extend our acknowledgements to the authorities of Ocampo and the people of San Miguel for their kind assistance and support for this proj ect. We are indebted to Francisco Palomino, Rafael Rodríguez and Edgar Guzmán (now retired) from INEGI and to the institutions and individuals involved in many ways to the success of this project.
REFERENCES
Burns, M.E., 2008, Taxonomic utility of ankylosaur (Dinosauria, Ornithischia) osteoderms: Journal of Vertebrate Paleontology, 28(4), 1102–1109. [ Links ]
Carpenter, K. 1990, Ankylosaur systematics: Example using Panoplosaurus and Edmontonia, in Carpenter, K., Currie, P.J. (eds.), Dinosaur Systematics: Approaches and Persepectives: Cambridge, Cambridge University Press, 281–298. [ Links ]
Carpenter, K., 1997, Ankylosaurs, in Farlow, J., Brett–Surman, M., (eds.), The Complete Dinosaur: Bloomington, Indiana University Press, 308–316. [ Links ]
Carpenter, K., 2001, Phylogenetic analysis of the Ankylosauria, in Carpenter, K. (ed.), The Armored Dinosaurs: Bloomington, Indiana University Press, 454–483. [ Links ]
Carpenter, K., Kirkland, J.I., 1998, Review of Lower and Middle Cretaceous ankylosaurs from North America, in Lucas, S.G., Kirkland, J.I., and Estep, J.W., (eds.), Lower and Middle Cretaceous Terrestrial Ecosystems: New Mexico Museum of Natural History and Sciencie Bulletin, 14, 249–270. [ Links ]
Coombs, W., 1978, The families of the ornithischian dinosaur order Ankylosauria: Journal of Paleontology, 21, 143–170. [ Links ]
Coombs, W.P., Jr., 1990, Teeth and taxonomy in ankylosaurs, in Carpenter, K., Currie, P. J. (eds.), Dinosaur Systematics: Approaches and Perspectives: Cambridge, Cambridge University Press, 269–279. [ Links ]
Coombs, W.P., Demeré, T.A., 1996, A Late Cretaceous nodosaurid ankylosaur (Dinosauria: Ornithischia) from marine sediments of Coastal California: Journal of Paleontology, 70, 311–326. [ Links ]
Coombs, W.P., Jr., Maryañska, T. 1990, Ankylosauria, in Weishampel, D.B., Dodson, P., Osmólska, H. (eds.), The Dinosauria: Berkeley, University of California Press, 1st Edition, 456–483. [ Links ]
Coria, R.A., Salgado, L., 2001, South American Ankylosaurs, in Carpenter, K., (ed. ), The Armored Dinosaurs: Bloomington, Indiana University Press, 159–168. [ Links ]
Ford, T., Kirkland, J.I., 2001, Carlsbad ankylosaur (Ornithischia, Ankylosauria): An ankylosaurid and not a nodosaurid, in Carpenter, K. (ed.), The Armored Dinosaurs: Bloomington, Indiana University Press, 239–260. [ Links ]
Gasparini, Z., Olivero, E., Scasso, R., Rinaldi, C., 1987, Un ankylosaurio (Reptilia, Ornithischia) campaniano en el continente antártico: Anais X Congreso Brasilero de Paleontologia, Río de Janeiro, Sociedad Brasileira de Paleontologia, 131–141. [ Links ]
Gasparini, Z., Peresa Suberbiola, X., Molnar, R.E., 1996, New data on the ankylosaurian dinosaur from the Late Cretaceous of the Antartica peninsula, in Novas, F.E., Molnar, R.E. (eds.), Proceedings of the Gondwanan Dinosaur Symposium, Memoirs of the Queensland Museum, 39, 583–594. [ Links ]
Lee, Y., M., 1996, A new nodosaurid ankylosaur (Dinosauria: Ornitischia) from the Paw Paw Formation (Late Albian) of Texas: Journal of Vertebrate Paleontology, 16, 232–245. [ Links ]
Lehman, T.M., 1987, Late Maastrichtian paleoenvironments and dinosaur biogeography in the Western Interior of North America: Palaeogeography, Palaeoclimatology, Palaeoecology, 60, 189–217. [ Links ]
Lehman, T.M., 1997, Late Campanian dinosaur biogeography in the Western Interior of North America, in Wolberg, D.A., Stump, E. (eds.), Dinofest International, Proceedings: Academy of Natural Sciences, Philadephia, 223–240. [ Links ]
Marsh, O.C., 1890, Additional characters of the Ceratopsidae, with notice of new Cretaceous dinosaurs: American Journal of Science, ser. 3, 39, 81–86. [ Links ]
Matthew, W., 1922, A super–dreadnaught of the animal world. The armoured dinosaur Palaeoscincus: Natural History, 22, 333–342. [ Links ]
McCrea, R.T., Lockley, M.G., Meyer, C.A., 2001, Global distribution of purported ankylosaur track occurrences, in Carpenter, K. (ed.), The Armored Dinosaurs: Bloomington, Indiana University Press, 413–454. [ Links ]
Meyer, C., Frey, E.D., Thüring, B., Etter, W., Stinnesbeck, W., 2005, Dinosaur tracks from the Late Cretaceous Sabinas Basin (Mexico): Kaupia, 14, 41–45. [ Links ]
Morris, W.J., 1967, Baja California: Late Cretaceous dinosaurs: Science, 155 (3769), 1539–1541. [ Links ]
Nopcsa, F., 1915, Die Dinosaurier der siebenbürgischen Landesteile Ungarns: Mitteilungen Jahrbuch der Königlichen ungarischen geologischen Reichsanstalt, 23, 1–26. [ Links ]
Osborn, H.F., 1923, Two Lower Cretaceous dinosaurs of Mongolia: American Museum Novitates, 95, 1–10. [ Links ]
Rivera–Sylva, H.E., Espinosa–Chávez, B., 2006, Ankylosaurid (Dinosauria: Thyreophora) osteoderms from the Upper Cretaceous Cerro del Pueblo of Coahuila, Mexico: Carnets de Géologie, Brest, Letter 2006/02 (CG2006 L02), 5. [ Links ]
Sankey, J., 2010, Faunal composition and significance of high–diversity, mixed bonebeds containing Agujaceratops mariscalensis and other dinosaurs, Aguja Formation (Upper Cretaceous), Big Bend, Texas, in Ryan, M.J., Chinnery–Allgeier, B.J., Eberth, D.A. (eds.), New Perspectives on Horned Dinosaurs: The Royal Tyrrell Museum Ceratopsian Symposium: Bloomington, Indiana University Press, 520–537. [ Links ]
Sereno, P. , 1986, Phylogeny of the bird–hipped dinosaurs (Order Ornithischia): National Geographic Research, 2, 234–25. [ Links ]
Sternberg, C.M., 1928, A new armored dinosaur from the Edmonton Formation of Alberta: Transactions of the Royal Society of Canada, 22 (sec. IV), 93–104. [ Links ]
Vickaryous, M.K., Maryañska, T., Weishampel, D.B. 2004, Ankylosauria, in Weishampel, D.B., Dodson, P., Osmólska, H. (eds.), The Dinosauria, 2nd Edition: Berkeley, University of California Press, 363–392. [ Links ]
Weishampel, D.B., Barrett, P.M., Coria, R.A., Le Loeuff, J., Xing, X., Xijin, Z., Sahni, A., Gomani, E.M.P., Noto, C.R., 2004, Dinosaur distribution, in Weishampel, D.B., Dodson, P., Osmólska, H., (eds.), The Dinosauria 2nd Edition: Berkeley, University of California Press, 517–606. [ Links ]

