










Servicios Personalizados
Revista
Articulo
 Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista mexicana de ciencias geológicas
versión On-line ISSN 2007-2902versión impresa ISSN 1026-8774
Rev. mex. cienc. geol vol.28 no.2 Ciudad de México ago. 2011
Funginella? isfahanensis n. sp. from the upper Albian of Iran
Funginella? isfahanensis n. sp. del Albiano superior de Irán
Mehdi Yazdi1, Ali Bahrami1, and Jacob Leloux2*
1 Department of Geology, Faculty of Science, University of Isfahan, 81746, Iran.
2 NCB Naturalis, Leiden, The Netherlands, private address: 3e Gortestraat 82, 2311 NM Leiden, The Netherlands. E–mail: * lelouxj@xs4all.nl
Manuscript received: June 2, 2010.
Corrected manuscript received: October 26, 2010.
Manuscript accepted: January 1, 2011.
ABSTRACT
Funginella? isfahanensis n. sp. is a new nominal solitary scleractiniid coral from the upper Albian of Iran. Its dimensions, associated sedimentary facies and descriptions, all conform to those of the four specimens described as "Funginella sp. 2" in Abdel–Gawad and Gameil (1995, Cretaceous and Palaeocene Coral Faunas in Egypt and Greece: Coral Research Bulletin 4, 1–36) from North Sinai, Egypt. A preliminary discussion on small solitary cupolate coral species is presented.
Key words: coral, cupolate–discoid growth, taxonomy, Kolah–Qazi section, Iran.
RESUMEN
Funginella? isfahanensis n. sp. es un nuevo coral escleractinio solitario del Albiano superior de Irán. Sus dimensiones, facies sedimentarias asociadas y descripción, concuerdan con las de cuatro especímenes descritos como "Funginella sp. 2" por Abdel–Gawad y Gameil (1995, Cretaceous and Palaeocene Coral Faunas in Egypt and Greece: Coral Research Bulletin 4, 1–36) para el Sinaí norte, Egipto. Se presenta una descripción preliminar de especies pequeñas de coral solitario en cúpula.
Palabras clave: corales, crecimiento en cúpula–discoidal, taxonomía, sección Kolah–Qazi, Irán.
INTRODUCTION
Little has been published about the solitary corals from the Albian of the Middle Eastern and southwestern Asian countries. Baron–Szabo et al. (2003) dealt with colonial corals from the neighborhood of Isfahan. Only a few publications from the Albian of Egypt were available for comparison. Aboul Ela et al. (1991) recorded "Micrabacia sp." as a common element in shales of the lower Albian of Gabal Manzour section (North Sinai, Egypt). No description was given, but two hemispherical solitary corals of less than 10 mm in diameter were illustrated. They have four complete cycles and a fifth incomplete septal cycle. Abdel–Gawad and Gameil (1995) described solitary corals from lower Albian from the same area: "Funginella" sp. 1 (b in Figure 1.6, 96–100 septa) and "Funginella" sp. 2 (a in Figure 1.6, 48–50 septa), "Paracycloseris" sp. 1 (c in Figure 1.6, 70–80 septa) and sp. 2 (d in Figure 1.6, 60–70 septa). They are found in "highly fossiliferous argillaceous limestones alternating with ferruginous sandstones and claystones" (Abdel–Gawad and Gameil , 1995, p. 3). Their original aragonitic composition is altered into calcite, which makes exact identification difficult.
However, the genus Paracycloseris Wells, 1934 was originally described by one species from sediments of Jamaica that were regarded as Campanian, but are now considered to be upper Maastrichtian (Löser, 2005, p. 188). Its holotype looks quite different: for instance, it has a broad papillose columella and has a patellate instead of a cupolate form (see the picture and description of the holotype of P. elizabethae Wells, 1934 in Baron–Szabo, 2002: 145, pl. 105, figs 1, 3, 4).
MATERIAL AND METHODS
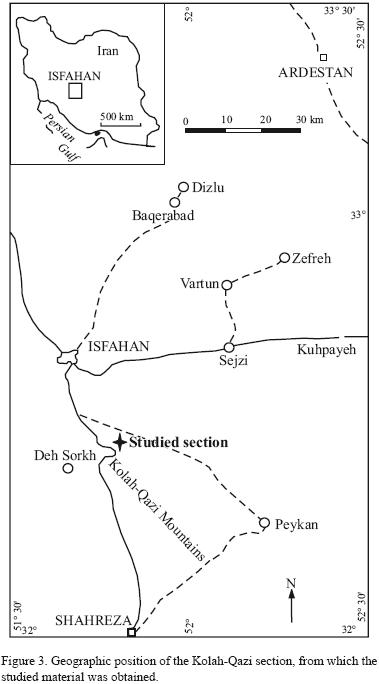
The studied material was collected from the middle part of the Albian Beudanticeras Shale (Figure 1.4 and Figure 2, see also Yazdi et al., 2009) at the Kolah–Qazi section (central Iran, 25 km southeast of Isfahan, near Baharestan new town, and 2 km south of Isfahan – Shiraz road, at the entrance of Kolah–Qazi valley, lat. N 51°46'32", long. E 32°26'50" GPS–WGS84 coordinates; Figure 3). One hundred and thirty two specimens were collected, from this material 122 are stored at Isfahan University as EUIM.3764–EUIM.3885 and ten specimens in the paleontological collection of NCB Naturalis in Leiden, The Netherlands, labeled RGM.532517 – RGM.532526.

The Beudanticeras Shale is a 120 m thick olive–green to grayish shale, with concretions, cone–in–cone structures, and intercalations of thin–bedded, lens–forming, dark gray limestone (5–30 cm), filled with small turritellid gastropods (Figure 1.5) and nuculid bivalves. Trigoniids and ammonites can be found throughout the unit (Tetragonites sp., Beudanticeras sp., Beudanticeras beudanti, Douvilleiceras sp., Puzosia sp. (Yazdi et al., 2009), which indicate the upper Albian stage.
The specimens were photographed in top, bottom and side views wherever possible, and the diameter (ø), height (h) and number of septa per 10 mm (ns/10mm) were measured. Petrographic thin sections were prepared for three specimens to establish details of the preservation and the top for one specimen was polished for analysis of the columella. Another thin section was prepared from a fragment of the limestone filled with turritellid gastropods.
Abbreviations used: EUIM: acronym for the collection at Isfahan University; MNHN: Musee National d'Histoire Naturelle, Paris, France; NCB: Netherlands Centre for Biodiversity, Leiden ,The Netherlands; RGM: acronym for the former "Rijks Geologisch en Mineralogisch Museum", now part of NCB Naturalis in Leiden, still in use as institution and collection codes for the geological collections. RMNH.COEL: acronym for the cnidarian collection of NCB Naturalis.
RESULTS
A thin section of RGM.532524 (Figure 4.1e) revealed that the original aragonite is altered into calcite. Crystals seem to cross the original boundaries of the septa and the filling between the septa. The thin sections of specimens RGM.532517 and RGM.532523 revealed the same structure. The recrystalization also explains why breaking a specimen did not result in a precise lateral view of the septa. The thin section of the consolidated packstone with the turritellid gastropods shows a very fine–grained limestone with dark elements, probably organic.
The scatter plot of height vs. diameter (Figure 4.4) of these specimens shows one cluster of points. Vaguely visible in the scatter plot are narrowings at diameters of about 4.0, 6.0 and 7.5 mm. Further statistics are presented in Table 1. The number of septa was calculated from the number of septa/10 mm counts multiplied by the diameter and π to get a comparable variable with the taxa in the literature. The range of height and diameter values for the present sample is also plotted as ellipse "L" in Figure 1.6. The dimensions of the present sample overlap those of the four specimens of "Funginella" sp. 2 in Abdel–Gawad and Gameil (1995, p. 22).
DISCUSSION
Because the dimensions of "Funginella sp. 2" in Abdel–Gawad and Gameil (1995, p. 22) are overlapped by the height and diameter of the present specimens and their description also fits the present material, it is only logical to assume that both populations belong to the same species. It is fully understandable that Abdel–Gawad and Gameil (1995) did not described their "Funginella sp. 2" as a new species, because they had only four specimens whose shell had been recrystalized. Since the sample used in the present study is large enough to have a clear picture of a population it is preferable to name the species. The maximum of the calculated septa (see Table 1) is 60. This is the case with, for instance, EUIM.3866. However, when counting the real number of septa instead of using the calculation, the number of septa turned out to be 48, but the shape of the coral was ellipsoid and not circular. EUIM.3817 has only 50 septa. So, the calculation of the number of septa by counting the number from only a part of the coral is not accurate. However, more than 48 septa mean that the fifth septal cycle has begun to develop.
Both Iranian and Egyptian materials seem to be found in comparable sedimentary rocks. Samples of the fossiliferous limestones of the Beudanticeras Shale that were collected with the corals show an association of gastropods dominated by only one form and are comparable with monospecific turritellid associations in Tertiary deposits, which would point to fully marine fine–grained muddy bottom about 10 to 40 m depth below sea grass environments (pers. comm. Frank Wesselingh). The low energy conditions as well as the organic part in the limestones confirm this "deeper" environment. It is well possible that sea level changes during deposition of the Beudanticeras Shale varied from about 30–50 m depth where the consolidated limestone lenses are probably the shallowest parts of the depositional environment.
SYSTEMATIC PALEONTOLOGY
Order Scleractinia Bourne, 1900
Suborder Fungiina Verrill, 1865
Family Funginellidae Alloiteau, 1952
Genus Funginella d'Orbigny, 1850
Funginella? isfahanensis n. sp.
Figures 1.1–1.3, 4.1a–4.1e, 4.2, 4.3
?1995 Funginella sp. 2. Abdel–Gawad and Gameil, 1995, p. 22, pl. 9, fig. 5.
Derivation of the name. After the city of Isfahan, close to the type locality.
Holotype. RGM.532526 (Figures 1.1, 1.2).
Paratypes. RGM.532517–RGM.532525; EUIM.3764–EUIM.3885 (Figures 4.1–4.3, 1.3).
Type locality. The Kolah–Qazi section, central Iran, 25 km southeast of Isfahan, near Baharestan new town, and 2 km south of Isfahan–Shiraz road, at the entrance of Kolah–Qazi valley (GPS–WGS84 coordinates lat. N 51°46'32", long. E 32°26'50").
Stratigraphic horizon. Beudanticeras Shale, upper Albian, Lower Cretaceous.
Diagnosis. Cupolate to discoid solitary corallum of about 6 mm in diameter and circa 1.5 mm in height having four cycles of costosepta, sometimes an incomplete fifth cycle all arranged in six systems and irregularly forking away from the center of the corallite, when looked from above. Costosepta equal in thickness, compact, synapticulae seldom present, septal margins dentate and septal sides granulate. Columella weakly developed.
Description. Small cupolate solitary coral without concentric rings on base. Shallow circular calicular fossa. Four complete septal cycles, commonly with an incomplete fifth cycle and arranged in six systems are present. Costosepta more or less regularly dentate and granulate on their sides. The first cycle continues to the centre of the corallite, the second has some large dentate structures which, together with the proximal dents on the first septa, can form a structure looking like a spongy columella. The third cycle seems to branch away from the second, while the fourth seems to branch away from the second in some specimens and sometimes away from the third septa in others. Costosepta are compact. In the thin section (Figure 4.1d) some intervals in the costosepta near the centre are visible. They are probably due to dentations of the costosepta. In some broken fragments (RGM 532520) a few wavy structures are visible, which could be interpreted as possible fulturae. However, since the breaking of the specimens goes through the septa, it is not completely sure if this observation is trustworthy. Thickness of costosepta is more or less the same for all the cycles. Calicular fossa is circular and small. Columella is not clearly visible, synapticulae are seldom present. (For dimensions see Table 1).
Remarks. The assignment of this species to a particular genus is not an easy task. There are several genera known to have small cupolate to discoid solitary corals. Wells (1956) cited Cyclastraea Alloiteau, 1952; Cycloseris Milne Edwards and Haime, 1849; Cyclolites Lamarck, 1801; Micrabacia Milne Edwards and Haime, 1849, and Discocyathus Milne Edwards and Haime, 1848. Since the specimens described from the Sinai were published as Funginella we take that genus also into account. Russo et al. (1996) listed Funginellastraea Alloiteau, 1952, Cyclolitopsis and Cyclophyllopsis having similar growth forms. Löser (pers. comm.) suggested regarding Microseris, Actinoseris and Asteroseris as possibilities.
Below, all these genera will be treated in alphabetical order having first a selected historical overview of the development of the concepts of these taxa combined with some remarks and comparisons with F.? isfahanensis.
Genus Actinoseris d'Orbigny, 1849
Type species. Actinoseris cenomaniensis d'Orbigny, 1850.
Actinoseris was established as "une Cycloseris dont la columelle est central, ronde et non en fente allongée" (d'Orbigny, 1849a: p. 12 [a Cycloseris where the columella is central, round and the calicinal fossa is not elongated]). Fromentel (1861) synonymised Actinoseris with Cycloseris. Actinoseris was considered a foraminifer by Vaughan and Wells (1943, p. 345), was kept as a separate species in Alloiteau (1957) and it was a junior synonym of Cycloseris according to Wells (1956, F388). Baron–Szabo (2002, p. 130) questionably regarded it as a junior synonym of Micrabacia. Löser (pers. comm.) regards it as a separate genus in the family Asteroseriidae, closely related to Microseris and related to the Fungiidae. Funginella? isfahanensis does fit into the original description of Actinoseris. The type species of Actinoseris differs from F.? isfahanensis in having medium common simple synapticulae, a columella of isolated granules and nine cycles of septa and being larger. Other Actinoseris species are Actinoseris alloiteaui Beauvais and Zlatarski, 1966 from upper Barremian to Lower Aptian of Bulgary and Actinoseris provincialis d'Orbigny, 1850 from the Cenomanian to Maastrichtian of France and Spain. A. provincialis is bigger. The holotype of A. alloiteaui is slightly larger than the largest specimens of F.? isfahanensis and its density of septa is smaller, but not out of the range of the current sample of F.? isfahanensis. The anastomization of the septa is similar.
Genus Asteroseris de Fromentel, 1867
Type species. Stephanoseris coronula de Fromentel, 1863.
Asteroseris de Fromentel was considered a junior synonym of Micrabacia by Vaughan and Wells (1943, p. 145). Asteroseris was characterized by "septes anastomosés sans ordre, endothèque absente, columelle essentielle, fasciculaire" (Alloiteau, 1952, p. 664). Wells (1956, p. F388) regarded Asteroseris as a junior synonym of Cycloseris. This opinion was shared by Baron–Szabo (2002, p. 97). Alloiteau (1957) and Löser (pers. comm.) retains Asteroseris as a separate genus. This is supported by the difference in shape of the calicular fossa (circular for Asteroseris versus elliptical for Cycloseris). F.? isfahanensis is not placed in Asteroseris due to the lack of a clear columella.
Asteroseris curonula differs from F.? isfahanensis in having a fair amount of simple synapticulae and a columella that is formed by fusion of the septa in the center and being relatively higher.
Genus Cyclastraea Alloiteau, 1952
Type species. Cyclolites spinosa de Fromentel, 1863.
Cyclastraea was defined as "discoid; muraille fortement plissée; septes entièrement compacts à bord distal armé de fortes dents aiguës; faces laterals avec careens verticals; columelle fasciculo–papilleuse" (Alloiteau, 1952, p. 655). Wells (1956) described Cyclastraea as "solitary, cupolate with basal epitheca. Septa imperforate with strong dentations and vertical carinae" (F379). Baron–Szabo (2002) regarded Cyclastraea as a questionable junior synonym of Cycloseris. Löser (pers. comm.) regards it to be a junior synonym of Microseris. F.? isfahanensis lacks the strong carinae.
Genus Cyclophyllopsis Alloiteau, 1952
Type species. Cyclolites aptiensis de Fromentel, 1863 (Type series: 3 paralectotypes MNHN: A25825, Lectotype M03597 selected by Alloiteau (1957, p. 341) should be the specimen figured by Fromentel in his pl. 66, fig. 3, 3a–b.).
Original diagnosis of the genus: "simple et discoid; muraille s'étendant sur la base et la moitié de la hauteur; synapticules assez nombreuses, par endroits coalescentes; pseudo–columelle fasciculo–papileuse" (Alloiteau, 1952, p. 655).
Baron–Szabo (2002) regarded Cyclophyllopsis as a questionable synonym of Cycloseris. Lectotype and paralectotypes do not belong to the same species and no thin section was found, the genus should be forgotten (Löser, pers. comm.).
Based on the original description Cyclophyllopsis aptiensis differs from Funginella? isfahanensis in being higher in juvenile stages, and having numerous synapticulae.
Genus Cycloseris Milne Edwards and Haime, 1849
Type species. Fungia cyclolites Lamarck, 1816 (syntypes: MNHN 73–74)
Cycloseris was originally described as "Polypier simple, libre. Cloisons très–nombreuses, s'unissant par leur bord interne" (Milne Edwards and Haime, 1849, p. 72).
The available material of recent C. cyclolites (RMNH.COEL.34559, '63–'64, '66, '69–'70) from New Caledonia shows a small ellipsoidal calice and a small ellipsoid spongy columella. The costae of C. cyclolites are dentate; the septa are arranged in four to six cycles in six systems. Lower cycles are stronger, thicker and higher than higher cycles. Young specimens vary from discoid to patellate, older specimens are increasingly cupolate in shape with concave bottom. Young specimens do not have a closed wall at the bottom, it closes when growing up. Adult specimens show clear rows of ornaments on the sides of the septa, more or less identical on each side of the septum and at the bottom of the septa bigger structures (fulturae) are developed, which grew on the septa in a later growth phase. F.? isfahanensis shows a closed wall at the bottom from the smallest to the largest specimens.
Russo et al. (1996) treated Cycloseris? escosurae Mallada, 1887 from the Lower Alpian of Spain, which has a similar behavior as Funginella? isfahanensis in that it often has small particles on which it has been growing. Cycloseris? escosurae is mostly attached on foraminiferal tests, while the present species prefers the more readily available turritellid shells. It has more septa and gets bigger and shows regular simple synapticulae, while F.? isfahanensis has hardly to no synapticulae.
Genus Cyclolites Lamarck, 1801
Type species. Cyclolites elliptica Lamarck, 1801 (="porpite elliptique… Guettard, 1770, mem. Vol. 2, p. 342, pl. 21, fig. 17, 18.").
Cyclolites was originally described as "Polypier libre, orbiculaire ou elliptique, convexe et lamelleux en–dessus, applati en desous avec des lignes circulaires concentriques. Il constitue une seule étoile lamelleuse" (Lamarck, 1801, p. 369), presenting four species according to the next order: C. numismalis, C. hemisphaerica, C. elliptica and C. cristata. Milne Edwards and Haime (1849) named C. elleptica [sic] Lamarck as only example of Cyclolites, which we regard as the first secondary designation of C. elliptica as type species for this genus. Alloiteau (1957, p. 331–332) presented a strong reasoning for this selection: Cyclolites as defined by Lamarck is a very polyphyletic group and only appropriate to be used as describing the overall shape of these corals: cyclolitoid, meaning hemispheroid corallite with a flat base, considered to be discoid to cupolate shaped. Cyclolites numismalis, as the first in the list of Lamarck's species, could be regarded as the type species of Cyclolites, but is a junior synonym of the Silurian rugose coral Madrepora porpita Linnaeus, 1767. This would make Palaeocyclus Milne Edwards and Haime, 1849, having the same type species, a junior synonym of Cyclolites. The second species of Lamarck, C. hemisphaerica, was suspected by Alloiteau to be a badly drawn C. elliptica, which was the third species of Lamarck. The last one, C. cristata, became the type species of Aspidiscus Kœnig, 1825. The type specimen(s) of C. elliptica are considered to be lost. To avoid confusion Alloiteau suggested that the main part of the Cretaceous "Cyclolites" were to be reassigned to Cunnolites. Alloiteau (1957) created Cunnolites barrerei as a replacement for C. elliptica and selected a specimen from coll. Depéret (lab. Geol. de Sorbonne) as "Neoholotype", supposedly from Coustouges, France, which is near Perpignan. Cyclolites as a genus name was reestablished by Löser (2009, p. 133) with Faujas de Saint–Fond, 1799 as author of the genus, since in Faujas (1799) referred directly to Lamarck (1801, p. 369) and this would mean that, if the date of publication of Faujas would have been correct, the name would indeed be first published, completely with description in Faujas and the author would have become "Lamarck in Faujas (1799)". However, Pasteur (1802), who translated Faujas (1799) into the Dutch language, stated that the first part of the Dutch translation was published only after the first five parts of Faujas had been published. The reference to Cyclolites and to Lamarck occurred in the second part of the Dutch translation (Pasteur, 1804), meaning that those references occurred in the parts of the French original that were published after 1802, so that Lamarck (1801) remains the correct author and year of publication of the generic name Cyclolites.
Cyclolites differs with Funginella in having perforations in their septa. Funginella? isfahanensis differs also from Cyclolites/Cunnolites species in being a lot smaller in its adult stage.
Genus Cyclolitopsis Reuss, 1874
Type species. Cyclolites patera d'Achiardi, 1867. Alloiteau (1952, p. 667) described this genus as patellate and fixed with a short peduncle when young. Alloiteau (1957) presented a more or less turbinate form. We know Cyclolitopsis only from Alloiteau (1952, 1957). Funginella? isfahanensis differs in being cupolate.
Genus Discocyathus Milne Edwards and Haime, 1848
Discocyathus Milne Edwards and Haime, 1848, differs in having clear pali and a clear columella and in not having a clear cupolate shape.
Genus Funginella d'Orbigny, 1849 sensu Alloiteau, 1957
Type species. Funginella neocomiensis d'Orbigny, 1850.
Funginella was originally described as "Cyclolites à calice circulaire, déprimé, don't la columelle est ronde et non pas transverse; les cloisons grosses, saillantes" (d'Orbigny, 1849a, p. 11). It was synonymised with Cyclolites in Fromentel (1867) and Wells (1936, p. 113). The latter selected Cyclolites haueriana Michelin, 1846 as "genolectotype". Alloiteau (1952, p. 662, 1957, p. 335) and also M. Beauvais (1982, tome II, 116–117) retained it as a separate species, while considering F. neocomiensis d'Orbigny, 1850, from the Hauterivian of France as the type. We assume that Alloiteau was not aware of Wells (1936) at that time and, at least, his citations of page numbers of the publications of d'Orbigny are erroneous. F. neocomiensis as described in Alloiteau (1957, p. 335) has compact septa, which separates it from Cyclolites. Funginella was regarded as a junior synonym of Cycloseris by Baron–Szabo (2002). We retain Funginella as a separate genus, regarding the shape of the calicular fossa as distinctive between the two genera, although further research is needed. Funginella was the generic assignment of the Sinai material in Abdel–Gawad and Gameil (1995), which is regarded to be the same species as the Iranian coral.
F.? isfahanensis differs from F. neocomiensis in having six systems of septa instead of eight and in having slightly less septa/10mm. F. neocomiensis has about 30 septa/10 mm. It differs from F. martini d'Orbigny, 1850 (23–26 septa/10mm) in having more septa/10mm.
Genus Funginellastraea Alloiteau, 1952
Type species. Funginella alpina d'Orbigny, 1850
First description: "Discoïde, libre; partie central du disque basal portant une Orthophragmina commensale; septes entièrement compacts, à faces laterals ornée de files nombreuses, serrées, de granules subcoalescents" (Alloiteau, 1952, p. 663). Russo et al. (1996) cited two other species with different foraminifera as substrate. Baron–Szabo (2002, p. 97) regarded Funginellastraea as a questionable synonym of Cycloseris.
Genus Micrabacia Milne Edwards and Haime, 1849
Funginella? isfahanensis does not belong to Micrabacia, since the costae and the septa are confluent, while Micrabacia is defined by the alternation of the septa and costae. However, the Campanian to Maastrichtian Micrabacia senoniensis from Western Europe is often preserved with only a small part of the basal wall, with the costae not preserved. The stratigraphic position of the present species is the same as the oldest reported Micrabacia specimens. However, their reports are vague (Micrabacia sp. in the Albian of North Sinai in Aboul Ela et al. (1991) and Micrabacia? sp. in the "Neocoomsandstein" in Tönsberg near Oerlinghausen, Germany in Weerth (1884). Further study of those specimens is necessary for better comparison and to check if those specimens are really Micrabacia.
Genus Microseris de Fromentel, 1867
Type species: Microseris hemisphaerica de Fromentel, 1867.
The genus Microseris was defined as "hémisphérique; la muraille, horizontalle, nue et couverte de granulations éparses qui ne simulent pas des côtes. Les cloisons sont larges, arquées et se réunissent en se soudant au centre où on remarque une petite fossette columellaire arrondie. Les synapticules sont rares, mais bien développées. Ce genre, voisin des Cycloseris, s'en distingue par son aspect general et l'absence de côtes sur le plateau. Celles–ci sont remplecées par des granulations éparses" (Fromentel, 1870 vol. 23, p. 367–368).
Microseris was considered to be a junior synonym of Micrabacia by Vaughan and Wells (1943, p. 145). Alloiteau (1952, p. 664) retained it as a separate genus. He diagnosed Microseris as "septes anastomosés suivant une loi precise, endothèque présente, columelle pariétale rudimentaire" (p. 664). Wells (1956, p. F388) regarded Microseris as a junior synonym of Cycloseris. Baron–Szabo (2002, p. 231) agreed with Wells. Löser (pers. comm.) retains Microseris as a separate genus within the Asteroseriidae. Funginella isfahanensis is not placed in Microseris, for its septa do not bifurcate according to a precise rule.
Microseris hemisphaerica differs from F.? isfahanensis in having nine cycles of septa and having simple synapticulae.
CONCLUSIONS
Actinoseris is a serious candidate as genus for this species. However, awaiting further research, we have assigned this species tentatively to Funginella sensu Alloiteau (1957) and Beauvais (1982) as is done with the material from the Sinai described by Abdel–Gawad and Gameil (1995). Both taxa and indeed most other small discoid to cupolate genera are in need of further revision.
ACKNOWLEDGEMENTS
We are grateful to Hannes Löser, Mohammed Gameil, Steve Donovan and two anonymous reviewers for their remarks, both in content and language. Bert Hoeksema is thanked for allowing us to study some recent Cycloseris Cyclolites specimens for comparisons.
REFERENCES
Abdel–Gawad, G. I., Gameil, M., 1995, Cretaceous and Palaeocene Coral Faunas in Egypt and Greece: Coral Research Bulletin 4, 1–36, pls 1–21. [ Links ]
Abou Ela, N.M., Abdel–Gawad, G.L., Aly, M.F., 1991, Albian fauna of Gabal Manzour, Maghara Area north Sinai, Egypt: Journal of African Earth Sciences 13, 201–220. [ Links ]
Alloiteau, J., 1952, Madréporaires post–paléozoïques in Piveteau, J. (ed.), Traité de paleontology: Paris, Masson et Cie, éditeurs 1, 539–684. [ Links ]
Alloiteau, J., 1957, Contribution à la systématique des Madréporaires fossils: Paris, C. N. R. S., 462 p. [ Links ]
Baron–Szabo, R.C., 2002, Scleractinian Corals of the Cretaceous: Knoxville, Tennessee, R.C. Baron–Szabo: 539 pp. [ Links ]
Baron–Szabo, R.C., Hamedani, A., Senowbari–Daryan, B., 2003, Scleractinian Corals from the Lower Cretaceous Deposits North of Esfahan (Central Iran): Facies 48, 199–216, pls 36–39. [ Links ]
Beauvais, M., 1982, Révision systématique des Madréporaires des couches de Gosau (Crétacé supérieur, Autriche): Paris, Travaux de laboratoire de paléontologie des invertebres, Universite Pierre et Marie Curie: 5 parts. [ Links ]
Beauvais, L., Zlatarski, V.N., 1966, Actinoseris? alloiteaui sp. n. – Madréporaire Crétacé inférieur de la Bulgarie: Comptes rendus de l'Academie bulgare des Sciences 19(12), 1171–1173, pl. 1. [ Links ]
Bourne, G.C., 1900, The Anthozoa in Lancester, E.R. (ed.), A treatise on zoology. Part II. The Porifera and Coelenterata: London, Adam & Charles Black, 80 p. [ Links ]
Faujas de Saint Fond, B., 1799–1803?, Histoire naturelle de la montagne St. Pierre de Maestricht: Paris, 263 p., 54 pls. [ Links ]
Fromentel, E. de, 1861, Introduction a l'étude des polypiers fossils: Mémoires de la Société d'émulation du Département du Doubs, Troisième série 5(1859–1860), 1–359. [ Links ]
Fromentel, M. de, 1862–1887, VIII. Zoophytes in Orbigny, A.D. d', 1840–1887, Paléontologie française, 1re Série, Terrain crétacé: Paris, chez l'auteur: 512 p., 131 pls [fide Löser (2002: 758–759) for dating of the pages and plates] [ Links ].
Guettard, J.–E., 1770, Mémoires sur différentes Parties des Sciences et Arts. Tome second: Paris, Laurent Prault, 530 p., 71 pls. [ Links ]
Kœnig, C.D.E., 1825, Icones fossilium sectiles. Centuria Prima: London, G.B. Sowerby: 4 p., 8 pls. [fide Weisbord, 1971] [ Links ].
Lamarck, J.B. de, 1801, Systême des animaux sans vertèbres: Paris, Musee d'Histoire Naturelle, 432 p. [ Links ]
Lamarck, J.B. de, 1816, Histoire Naturelle des Animaux sans Vertèbres: Paris, Verdière 2, 568 p. [ Links ]
Linné, C. a, 1767, Systema Naturæ, per Regna Tria Naturae, secundum Classes, Ordines, Genera, species cum Characteribus, Differentiis, Synonymis, Locis. Tomus I. Editio decimal tertia: Stockholm, Ioannis Thomae, 1361 p. [ Links ]
Löser, H., 2002, Catalogue of Cretaceous Corals. Volume 2. List of Citations: Dresden, Cpress Verlag, 784 p. [ Links ]
Löser, H., 2005, Catalogue of Cretaceous Corals. Volume 3. List of Localities: Dresden, CPress Verlag 3, 366 p. [ Links ]
Löser, H., 2009, Fossile Korallen aus Jura und Kreide. Aufbau, Klassification, Bestimmung und Fundmögligkeiten: Dresden, CPress Verlag, 206 p. [ Links ]
Mallada, L., 1887, Sinópsis de las especies fósiles que se han encontrado en España: Tomo III, Terreno mesozóico, Cretáceo inferior: Boletín de la Comisión del Mapa geológico de España, 14, 1–167 [ Links ]
Milne Edwards, H., Haime, J., 1848, Recherches sur les polypiers; deuxième mémoire, Monographie des Turbinolides: Annales des Sciences Naturelles. Zoölogie, III–9, 211–344, pls 7–10. [ Links ]
Milne Edwards, H., Haime, J., 1849, Mémoire sur les polypiers appartenant à la famille des oculinides, au groupe intermédiaire des Pseudoastréides et à la famille des Fongides: Comptes rendus hebdomadaires des séances de l'Académie des Sciences, 29, 67–73. [ Links ]
Mutterlose, J., 1995, Die Unterkreide–Aufschlüsse des Osning–Sandsteins (NW–Deutschland) – ihre Fauna und Lithofacies: Geologie und Paläontologie in Westfalen, 36, 1–85, 10 pls. [ Links ]
Orbigny, A. d', 1849a, Note sur des Polypier fossils: Paris, Victor Masson, 12 p. [ Links ]
Orbigny, A. d', 1849b, Note sur les Polypiers fossils: Revue et Magasin de Zoologie pure et applicée, Série 2, 1, 526–528. [ Links ]
Orbigny, A. d', 1850, Catalogue des espèces fossils de Mollusques Bryozoaires, de Polypiers et d'Amorphozoaires de l'étage néocomien: Revue et Magasin de Zoologie pure et appliqué, Serie 2, 2, 170–181. [ Links ]
Pasteur, J.D., 1802, Natuurlijke historie van den St. Pieters Berg bij Maastricht. Eerste stuk: Amsterdam, Johannes Allart: 186 p., 19 pls. [ Links ]
Pasteur, J.D., 1804, Natuurlijke historie van den Sint Pieters–berg. Tweede stuk: Amsterdam, Johannes Allart, 187–340, pls 20–52. [ Links ]
Russo, A. , Cherchi, A., Schroeder, R., 1996, An example of auto–mobility and host substrate relation ship in "Cycloseris" escosurae Mallada, 1887, scleractinian coral from Lower Aptian of Spain: Bollettino della Società Paleontologica Italiana Special Vol. 3, 191–203. [ Links ]
Stolarski, J., Roniewicz, E., 2001, Towards a new synthesis of evolutionary relationships and classification of Scleractinia: Journal of Paleontology, 75(6), 1090–1108. [ Links ]
Vaughan, T.W., Wells, J.W., 1943, Revisions of the suborders, families, and genera of the Scleractinia: Geological Society of America, special papers, 44, 363 p. [ Links ]
Verrill, A.E., 1865, List of Polyps and corals sent by the Museum of Comparative Zoology to other institutions in exchange, with annotations: Bulletin of the Museum of Comparative Zoology, 1, 29–60. [ Links ]
Weerth, O., 1884, Die Fauna des Neocoomsandsteins im Teutoburger Walde: Palaeontologische Abhandlungen, 2(1), 77 p., 11 pls. [ Links ]
Weisbord, N.E., 1971, Bibliography of Cenozoic Echinoidea including some Mesozoic and Paleozoic Titles: Bulletins of American Paleontology, 59, 314 p. [ Links ]
Wells, J.W., 1934, Some fossil corals from the West Indies: Proceedings of the United States National Museum, 83, 71–110. [ Links ]
Wells, J.W., 1936, The nomenclature and type species of some genera of recent and fossil corals: American Journal of Science, Fifth series, 31(182), 97–134. [ Links ]
Wells, J.W., 1956, Scleractinia in Bayer, F.M., Boschma, H., Harrington, H.J., Hill, D. , Hyman, L.H. , Lecompte, M. , Montanaro–Gallitelli, E. , Moore, R.C. , Stumm, E.C., Wells, J.W., Part F, Coelenterata in Moore, R.C. (ed.), 1953–1969, Treatise on Invertebrate Paleontology: Lawrence, Kansas, The University of Kansas Press and The Geological Society of America, F328–F444. [ Links ]
Yazdi, M., Bahrami, A., Vega, F.J., 2009, Albian decapod Crustacea from Southeast Isfahan, Central Iran–Kolah–Qazi area: Bulletin of the Mizunami Fossil Museum, 35, 71–77. [ Links ]

