










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkRevista mexicana de ciencias geológicas
On-line version ISSN 2007-2902Print version ISSN 1026-8774
Rev. mex. cienc. geol vol.28 n.1 Ciudad de México Apr. 2011
Microfossils, paleoenvironments and biostratigraphy of the Mal Paso Formation (Cretaceous, upper Albian), State of Guerrero, Mexico
Microfósiles, paleoambiente y bioestratigrafía de la Formación Mal Paso (Cretácico, Albiano superior), Estado de Guerrero, México
Harry F. Filkorn1,* and Robert W. Scott2
1 Physics and Planetary Sciences Department, Los Angeles Pierce College, 6201 Winnetka Avenue, Woodland Hills, California 91371 USA.
2 Precision Stratigraphy Associates and University of Tulsa, 149 West Ridge Road, Cleveland, Oklahoma 74020, USA. * filkornh@piercecollege.edu
Manuscript received: April 6, 2010
Corrected manuscript received: October 14, 2010
Manuscript accepted: December 13, 2010
ABSTRACT
Microfossils from an outcrop of the coral reef and rudist–bearing calcareous upper member of the Mal Paso Formation just north of Chumbítaro, State of Michoacán, Mexico, indicate a deepening trend and transition from nearshore through outer shelf depositional environments upward through the sampled stratigraphic interval. The microbiota is mostly composed of species of calcareous algae and foraminifera. The identified calcareous algae are: Pseudolithothamnium album Pfender, 1936; Cayeuxia kurdistanensis Elliott, 1957; Acicularia americana Konishi and Epis, 1962; and Dissocladella sp. cf. D. savitriae Rama Rao and Pia, 1936. The species of foraminifera are: Nezzazata sp. cf. N. isabellae Arnaud–Vanneau and Sliter, 1995; Buccicrenata subgoodlandensis (Vanderpool, 1933); Cuneolina parva Henson, 1948; Pseudolituonella sp.; Praechrysalidina sp.; and Rotalipora appenninica (Renz, 1936). In addition, a species of stromatoporoid is illustrated and an indeterminate tube–shaped calcitic microorganism is described as an incertae sedis. From the base of the section upward, four biofacies are defined by the co–occurrences of these taxa: a benthic foraminiferal assemblage, a coral assemblage, a caprinid – dasycladacean assemblage, and a coral – miliolid assemblage. This report documents the first detailed examination of the microbiota of the calcareous upper member of the Mal Paso Formation. Data from this analysis of the microbiota supplement earlier paleoenvironmental interpretations based on studies of macrofossils, mainly scleractinian corals, rudists, and other mollusks, and carbonate facies relationships.
The combined stratigraphic ranges of the microfossil species identified from this measured section of the Mal Paso Formation support an age determination of late Albian. The occurrence of Rotalipora appenninica (Renz, 1936), a planktic foraminiferan, in the uppermost portion of the exposed stratigraphic section is especially significant because its presence indicates a marked deepening of the depositional environment which can be correlated with the onset of the global late Albian marine transgression and drowning of Tethyan carbonate platforms that is known as the R. appenninica – event. A late Albian age was also suggested by previous studies of other taxonomic groups that have been discovered in the same stratigraphic section, particularly the species of the rudist bivalve genus Mexicaprina Coogan, 1973.
Key words: foraminfera, algae, paleoenvironment, Mal Paso Formation, Cretaceous, Albian, Mexico.
RESUMEN
Microfósiles del miembro superior calcáreo de la Fomación Mal Paso colectados en un afloramiento justo al norte de Chumbítaro, Estado de Michoacán, indican una tendencia la profundización y una transición de un ambiente de depósito de plataforma interna a uno de plataforma externa hacia la porción superior del intervalo estratigráfico muestreado. La microbiota se compone mayoritariamente de especies de algas calcáreas y foraminíferos. Las algas calcáreas identificadas son: Pseudolithothamnium album Pfender, 1936; Cayeuxia kurdistanensis Elliott, 1957; Acicularia americana Konishi y Epis, 1962; y Dissocladella sp. cf. D. savitriae Rama Rao y Pia, 1936. Las especies de foraminíferos son: Nezzazata sp. cf. N. isabellae Arnaud–Vanneau y Sliter, 1995; Buccicrenata subgoodlandensis (Vanderpool, 1933); Cuneolina parva Henson, 1948; Pseudolituonella sp.; Praechrysalidina sp.; and Rotalipora appenninica (Renz, 1936). Adicionalmente, se ilustra una especie de estromatopórido y se describe como incertae sedis un organismo calcítico tubular no determinado. Desde la base y hacia la porción superior de la sección se definen cuatro biofacies por la concurrencia de estos taxa: un ensamble de foraminíferos, un ensamble de corales, un ensamble de caprínidos– dasycladáceas, y un ensamble de corales–miliólidos. Este reporte documenta el primer examen detallado de la microbiota del miembro superior calcáreo de la Formación Mal Paso. Los datos derivados de este análisis de la microbiota complementan interpretaciones paleoambientales previas basadas en estudios de microfósiles, principalmente corales escleractinios, rudistas, y otros moluscos, así como en relaciones de facies carbonatadas.
Los alcances estratigráficos combinados de las especies microfósiles identificadas en la sección medida de la Formación Mal Paso apoyan la determinación de una edad del Albiano tardío. La presencia de Rotalipora appenninica (Renz, 1936), un foraminífero planctónico, en la porción más alta de la sección estratigráfica expuesta es especialmente significativa dado que indica una marcada profundización del ambiente de depósito, el cual puede ser correlacionado con el inicio de la trasgresión marina global del Albiano tardío y con la inundación de las plataformas carbonatadas del Tethys conocido como evento R. appenninica. Una edad correspondiente al Albiano tardío había sido también sugerida en estudios previos de otros grupos taxonómicos que fueron descubiertos en la misma sección estratigráfica, particularmente las especies del género de rudistas Mexicaprina Coogan, 1973.
Palabras clave: foraminíferos, algas, paleoambiente, Formación Mal Paso, Cretácico, Albiano, México.
INTRODUCTION
The Albian Mal Paso Formation is a significant, thick lithostratigraphic unit in southwestern Mexico (Pantoja–Alor, 1959, 1992). The Mal Paso Formation is divided informally into two parts, a lower siliciclastic and volcaniclastic unit and an upper carbonate interval. The upper carbonate interval has yielded diverse and abundant corals, mollusks, echinoids, foraminifera, and calcareous algae (García–Barrera and Pantoja–Alor, 1991). It was deposited in a tectonically active island–arc setting during the late Albian relative sea–level rise. A similar microfossil assemblage is known from Albian shallow–water carbonates in Ahuacatlán, Querétaro (Buitrón–Sánchez et al., 1995). Aptian benthic foraminifera have been reported from the nearby El Cajon locality (Omaña–Pulido and Pantoja–Alor, 1998) which is stratigraphically below the Mal Paso Formation. This section is a data point relevant to hypotheses about the replacement of coral communities by rudist communities.
We describe for the first time the microfossil assemblage of the Mal Paso Formation. The microfossils are from a fossiliferous outcrop of the calcareous upper member of the Mal Paso Formation that is located in the State of Guerrero, on the western limb of the Mal Paso syncline, approximately 0.75 km north of the rural village of Chumbítaro, Michoacán (Figure 1). A measured stratigraphic section at this locality is composed of a continuous sequence of limestone strata about 200 m thick. This interval contains horizons with significant in situ accumulations of oysters, reef corals, stromatoporoids, and rudist bivalves (Figure 2). The strata of this section have been assigned numbers from the base upward, units MP1 through MP18, in order to facilitate lithologic descriptions and as a reference frame for the positions of fossiliferous horizons and lithologic samples. Thin sections prepared from lithologic samples taken throughout this section have yielded a diverse microbiota that complements the paleontological and paleoecological data derived from previous studies of other macrofaunal groups of the Mal Paso Formation including scleractinian corals (Filkorn and Pantoja–Alor, 2004, 2009), rudist bivalves (Filkorn, 2002a), gastropods (Buitrón–Sánchez and Pantoja–Alor, 1994, 1996, 1998), and echinoids (García–Barrera and Pantoja–Alor, 1991). The 23 thin sections examined in this study (Table 1) also included three from samples of beds below the base of the upper member, in the clastic lower member of the formation (samples labeled HFLMP1, 2, and 3), but these thin sections are barren of microfossils. All 20 of the thin sections of samples from beds in the upper member contained identifiable microfossils (Table 1). Because facies of the Mal Paso change greatly laterally and vertically at other localities (García–Barrera and Pantoja–Alor, 1991), the conclusions presented herein pertain to this site only.


PALEOECOLOGY
The studied section of the upper member of the Mal Paso Formation records a major late Albian marine transgression and drowning of the area from the base of the section upward through the highest sampled bed. The shift from continental siliciclastics and volcaniclastics in the top of the lower member to predominantly calcium carbonate sediments in the basal part of the upper member marks a significant shift in depositional setting and begins the transition of environments upward through the section from lagoonal to back reef, coral–stromatoporoid reef, caprinid rudist forereef, distal reef bioclastics, and ultimately deposits in deeper water. The presence of small colonies of corals (Ovalastrea sp., Preverastraea sp.) and stromatoporoids, rudist bivalves, miliolids and other benthic foraminifera suspended in a fine–grained matrix in the lower part of the section indicate normally quiet and relatively shallow marine conditions of a lagoonal or inner shelf setting. A bed with abundant recrystallized fragments of a branching dendroid coral species and a species of red alga (unit MP7) occurs just below an in situ deposit of oyster shells (unit MP8). Above this horizon, the sedimentology and diverse biota of small, isolated coralla of massive and phaceloid colonial coral species, species of the rudist Radiolites, and species of other mollusks such as nerineid gastropods and the pectinid bivalve Neithea [Pecten] roemeri Hill, 1889, indicate an increased affinity with a coral reef environment. The microbiota from this same part of the section (units MP9 – MP12) is in the caprinid – dasyclad biofacies and mainly composed of a diverse assemblage of species of calcareous green algae (Cayeuxia and Dissocladella), encrusting red algae (Pseudolithothamnium) and benthic foraminifera (Table 1). Thus, the biota clearly supports the interpretation of deposition in a shallow marine environment that was well within the depth limits of the photic zone. The rudist species Mexicaprina alata has its lowest recognized occurrence in this stratigraphic section of the upper member in unit MP12 where it occurs with Neithea roemeri and abundant quantities of other molluscan bioclastics.
A 19 m–thick coral reef horizon (unit MP13) overlies and was initially established on the molluscan bioclastic debris of unit MP12. The fauna of the coral reef interval includes a diverse assemblage of at least 15 in situ colonial scleractinian coral species with encrusting, massive, ramose, foliaceous and phaceloid growth forms, a species of stromatoporoid (Actinostromaria sp., which is most abundant in the upper part of this unit), species of the rudist Radiolites and Mexicaprina, large articulated specimens of Neithea roemeri, and nerineid gastropods (Filkorn and Pantoja–Alor, 2009). At the scale of the outcrop and in hand sample, the sediments deposited between the larger coral colonies and mollusks in the coral reef horizon could be classified as an echinoid spine wackestone or floatstone with local packstone. Notably, echinoid tests were not observed in the same bed and the obvious presence of the majority of the larger taxa in the reef facies is not evident from examination of the fine–grained bioclastics of the intra–reef rock. Thus, the microbiota of the coral reef horizon is within the caprinid – dasyclad biofacies and the algal – caprinid microfacies, similar to that of the stratigraphically adjacent beds, and the identified elements are mostly caprinid rudist fragments, calcareous green algae (Dissocladella) and benthic foraminifera (miliolids, Buccicrenata, Cuneolina).
The coral reef horizon is conformably overlain by a bed of Mexicaprina floatstone (unit MP14, samples M48 and M49 in Table 1) that is approximately 6.5 m–thick. Large colonial corals are absent from this stratum and the macrofossil assemblage is almost exclusively composed of valves of the rudist species Mexicaprina alata. The microbiota is primarily composed of calcareous green algae (Cayeuxia and Dissocladella), therefore the observed algal – caprinid microfacies and petrographically determined caprinid – dasyclad biofacies are the same as those of the underlying coral reef horizon. Thus, the stratigraphic relationship coupled with the identified biota indicate that the Mexicaprina floatstone was deposited in a forereef setting adjacent to the coral reef environment and that the physical conditions of the habitat also were within the limits of the essential ecological requirements (light intensity, water temperature and bathymetry) of calcareous green algae. The distribution of the Mexicaprina valves and colonial corals in the stratigraphic interval which spans units MP12 – MP14, i.e., Mexicaprina valves transported into and behind the coral reef, but no corals transported in the opposite direction and forward of the reef front into the forereef facies, suggests that the prevailing water currents and wave energy were from an offshore direction and toward the reef front, as would be expected in most coastal settings.
Above the top of unit MP14, in unit MP15, the taxonomic diversity decreases, calcareous algae are absent, and the microbiota is primarily composed of benthic foraminifera (miliolids and Cuneolina). In addition, the bedding changes noticeably in the strata above unit MP14, where strata become thin– to medium–bedded and an intercalated argillaceous mudstone and wackestone component is introduced, concurrent with a decrease in macrofossils and an increase in the degree of sorting of fine–grained bioclastics. The top of unit MP14 is the highest observed occurrence of Mexicaprina valves in this stratigraphic section.
Higher up the stratigraphic section, in a 1.2 m–thick wackestone bed (unit MP17), six species of colonial corals occurred as isolated coralla along with other species of caprinid rudists, but the macrofauna is not entirely the same as that of the underlying units. The coral fauna of this unit is predominantly composed of species with a ramose growth form and relatively thin branches. Although four of these six coral species also occurred in the main coral reef horizon (unit MP13), the other two coral species were not observed anywhere lower in the stratigraphic section. The scattered distribution of isolated ramose coralla within this interval suggests that the substrate may have been undergoing an initial stage of coral colonization, but evidently the environmental conditions were not optimal for the growth of coral thickets or further development into a coral reef. Notably, unit MP17 contains the highest stratigraphic occurrence of colonial reef–building coral species in this stratigraphic section. Benthic foraminifera and rare calcareous alga also occur in unit MP17, an indication that conditions may have been favorable for coral growth for a relatively brief time. This upper part of unit MP17 likely represents deposition in the outer zone of the middle shelf and at the lower limits of the bathymetric ranges suitable for the existence of zooxanthellate colonial reef corals and calcareous green algae.
The uppermost part of unit MP17 contains an argillaceous horizon with abundant whole de–spined tests of regular echinoids as well as irregular echinoid tests, but the spines are not preserved in the same bed. In contrast, echinoid spines are common in the intra–reef rock of the coral reef horizon (unit MP13), but no echinoid tests were found in the same bed. The combination of these kinds of occurrences has been interpreted as evidence that the regular echinoids lived in the shallow–water coral reef habitat where they were de–spined and that their whole tests were later transported into deeper water by weak bottom currents flowing in an offshore direction.
The stratigraphically highest bed exposed in this measured section of the upper member is a 1 m–thick deposit of fine–grained wackestone, unit MP18. This bed has yielded two species of scleractinian corals: Blastozopsammia guerreroterion Filkorn and Pantoja–Alor, 2004, a thin–branched, ramose, colonial dendrophylliid, and Paracycloseris effrenatus Filkorn and Pantoja–Alor, 2009, a small, disc–shaped or cupolate, solitary cunnolitid. The dendrophylliid species B. guerreroterion belongs to a group of scleractinian corals that is recognized as primarily inhabiting deep– or cold–water environments, therefore this species presumably lacked the kind of algal symbionts that are typical of modern reef–building corals (i.e., it was azooxanthellate) and lived beneath the lower limit of the photic zone. The small, solitary coral species P. effrenatus also was not a reef–building form and it is likely that it too was azooxanthellate and existed in deeper water. The microbiota identified from unit MP18 is particularly significant because it is predominantly composed of benthic foraminifera (Cuneolina, Ne zzazata, miliolids), calcareous algae are absent, and one species of planktic foraminifera, Rotalipora appenninica (Renz, 1936) makes its first appearance in the section (see Table 1). The lack of calcareous algae, a common biotic component of the underlying strata in this section, suggests that the depositional environment had become deeper upward into this horizon of the upper member and that the substrate was now below the photic zone. The presence of the planktic foraminiferan species R. appenninica in unit MP18 is extremely significant because it adds further support to this interpretation. The Rotalipora appenninica Zone spans an interval of carbonate platform drowning in the Tethys typically referred to as the R. appenninica–event (Grötsch et al., 1993). On carbonate platforms in the Mediterranean region this stratigraphic interval overlies a karstic contact that represents a sequence boundary. Thus, the entire fossil assemblage from unit MP18 supports the interpretation that the environment of deposition was in a deeper–water setting. This interpretation completes the stratigraphic transition of depositional environments upward through the upper member of the Mal Paso Formation at this locality and supports the overall interpretation that this section records a major marine transgression.
To summarize, the four biofacies in the Mal Paso Formation are defined by the co–occurrence of fossil groups. In stratigraphic order from the base they are: the benthic foraminiferal assemblage; the coral assemblage; the caprinid–dasyclad assemblage; and the coral–miliolid assemblage (Table 1). These biofacies document a progression from nearshore to offshore depositional environments at this locality. The benthic foraminiferal assemblage represents the inner shelf miliolid assemblage (Sliter and Baker, 1972) and the middle to external carbonate shelf (Tronchetti, 1984). In the Middle East the genera range in depth from 10 to 65 m (Banner and Simmons, 1994). The calcareous algal assemblage of Pseudolithothamnium album, Cayeuxia kurdistanensis, Acicularia americana, and Dissocladella without udoteacean algae compose the Dasyclad–Rhodophyte assemblage and represents depths of 15 to 30 m (Banner and Simmons, 1994). The same model projects planktic foraminifera appearing at outer platform depths below about 60 m. The stratigraphic succession in species diversity and composition reflects a progressive change in environment from nearshore with quartz sand influx to shallow shelf followed by late stage deepening to approximately 60 to 65 m.
The co–occurrence of rudists and colonial corals and their formation of buildups can be used to test hypotheses posed to explain the apparent replacement of coral paleocommunities by rudist paleocommunities. One hypothesis proposes that late Albian rudists out–competed corals in a high salinity "Supertethys" zone (Kauffman and Johnson, 1988). Another hypothesis, so–called "Strangelove" pump with niche exclusion, proposes that Albian corals occupied deeper parts of the shelf than rudists and were stressed repeatedly by Albian anoxic events OAE 1b, 1c, and 1d (Scott, 1995). If the present position of this section relative to northern Mexico is the same as suggested by the Late Aptian plate reconstruction of Mann (1999, fig.16), it was well within the "Supertethys" hypersaline zone. Thus corals were not excluded by the local salinity, which was approximately 'normal' as suggested by the diverse fossil assemblage. Likewise no direct evidence of periodic low oxygen waters supports the niche exclusion by successive dysaerobic conditions. However, the progressive deepening and vertical succession of corals after rudists supports the idea that corals tended to occupy deeper substrates than many rudist assemblages. In this case, the apparent community replacement may be a result of superposition of laterally adjacent carbonate facies (Filkorn, 2002b). Additional well exposed Upper Albian – Lower Cenomanian carbonate sections in the so–called "Supertethys" zone need to be documented to understand the apparent replacement of coral communities by rudist communities.
BIOSTRATIGRAPHY
The calcareous algae and foraminifera species from this section of the calcareous upper member of the Mal Paso Formation support a late Albian age determination. The planktic foraminiferal Rotalipora appenninica Zone in the upper part of the section (unit MP18) has been correlated with latest Albian ammonite zones (Premoli Silva and Sliter, 2002; Scott, 2009). This zone is defined as the interval above the first occurrence (FO) of the nominate species up to the FO of Rotalipora brotzeni (Sigal) that is at the base of the Cenomanian Stage. The species Buccicrenata subgoodlandensis (Vanderpool, 1933), Cuneolina parva Henson, 1948, Nezzazata sp. cf. N. isabellae Arnaud–Vanneau and Sliter, 1995, Pseudolituonella sp., and Praechrysalidina sp. of the benthic foraminiferal assemblage all have longer stratigraphic ranges, but they also are consistent with a late Albian age. Thus, the stratigraphic ranges of the foraminifera support the previous age determination of the strata at this locality that was mainly based upon the occurrence of species of the caprinid rudist Mexicaprina Coogan, 1973 (Filkorn, 2002a). This genus characterizes the uppermost Albian rudist zone in the Gulf Coast (Scott and Filkorn, 2007).
The upper Mal Paso Formation at this site records the latest Albian relative rise in sea level and a marine transgression which correlate with a widespread sea–level cycle. In north Texas, the Pawpaw and Main Street formations in the upper part of the Washita Group, cycle WA 5, represent a siliciclastic flooding and shoaling into a carbonate shelf (Scott et al., 2003). This latest Albian sea–level cycle is also recorded in Europe (Amédro, 2008; Amédro and Robaszynski, 2008). However, as with many other areas in southwestern Mexico, deposition of the Mal Paso Formation was also affected by local tectonics (García–Barrera and Pantoja–Alor, 1991).
SYSTEMATIC PALEONTOLOGY
The thin sections which contain the fossils described herein are in the collections of the Museo de Paleontología, Instituto de Geología (= IGM), Universidad Nacional Autónoma de México, Mexico City. The illustrated specimens are denoted by IGM specimen numbers in the figure captions. The suprageneric classification of calcareous algae is as yet developing (Saunders and Hommersand, 2004). Thus, we use the classification of Wray (1978). For foraminifera, the suprageneric classification of Loeblich and Tappan (1988) is followed.
Phylum Rhodophyta
Class Rhodophyceae
Order Cryptonemiales
Family Squamaraceae Hauck, 1885
Genus Pseudolithothamnium Pfender, 1936
Pseudolithothamnium album Pfender, 1936
1936 Pseudolithothamnium album Pfender, 1936, p. 304–308, pl. 19, figs. 1–5.
1963 Ethelia alba (Pfender), Elliott, p. 293–294, pl. 47, figs. 1–3.
1969 Ethelia alba (Pfender), Johnson, p. 31–32, pl. 19, figs. 1–2 (previous synonymy provided).
1986 Pseudolithothamnium album Pfender, 1936, Kuss, p. 232–233, fig. 6b.
1991 Pseudolithothamnium album Pfender, 1936, Kuss and Conrad, p. 878.
Remarks. Pseudolithothamnium album encrusts micrite nodules and large bioclasts in the basal bed of the upper member of the Mal Paso Formation where it occurs with the rudist Mexicaprina cornuta. The thalli are 0.089–0.179 mm thick and composed of curved tubes 7 to 21 µm in diameter. Full descriptions are provided by Johnson (1969) and Kuss and Conrad (1991).
Stratigraphic range. This species has been reported from Upper Albian strata in Texas (Johnson, 1969) and Maastrichtian to Paleocene strata in Egypt, Sinai and Jordan (Kuss and Conrad, 1991). In the upper member of the Mal Paso Formation, this species has been recognized in several horizons in the lower part of the section, below the main coral reef interval (MP13).
Phylum Chlorophyta
Class Chlorophyaceae
Order Siphonales
Family Codiaceae Kützing, 1843
Genus Cayeuxia Frollo, 1938
Cayeuxia kurdistanensis Elliott, 1957
1957 Cayeuxia kurdistanensis Elliott, p. 790–791, pl. 25, figs. 8 – 10.
1969 Cayeuxia kurdistanensis Elliott, Johnson, p. 37–38, pl. 26, fig. 1.
1970 Cayeuxia kurdistanensis, Dragastan, p. 120.
1995 Cayeuxia kurdistanensis Elliott, Buitrón–Sánchez et al., p. 150, pl. 3, fig. 1.
2005 Cayeuxia (Rivularia) kurdistanensis Elliott, Palma et al., p. 128, fig. 6C.
Description. Thallus a nodular mass, height 1 mm, width 0.804 mm, and composed of slender oval radial tubes 0.018–0.036 mm (18–36 microns) in diameter. Radial tube walls 0.007–0.014 mm (7–14 microns) thick and composed of microcrystalline calcite. Tubes filled with calcite spar; horizontal partitions absent, although calcite spar exhibits horizontal fractures and faces. Tubes diverge toward distal margin of nodule and bifurcate at acute angles from preceding tubes. Tubes branch at angles approaching 45° only near tuft margin (as described by Johnson, 1969).
Stratigraphic range. The dimensions of the specimen from the Mal Paso Formation are most similar to those of Elliott's (1957) species, which has been reported from Iraq, France, and Texas (Johnson, 1969). This species also is known from other Cretaceous localities in Mexico (Albian; Buitrón–Sánchez et al., 1995, and references therein), Europe (Dragastan, 1970) and the Middle East (Johnson, 1969). It also has been reported from Middle Jurassic nearshore carbonates in Argentina (Palma et al., 2005).
Order Dasycladales
Family Dasycladaceae Kützing, 1843
Genus Acicularia d'Archiac, 1843
Acicularia americana Konishi and Epis, 1962
1962 Acicularia americana Konishi and Epis, p. 71–72, pl. 1, figs. 7 – 9, 11, 14.
Description. Spicule stem solid, diameter 0.232 mm. Sporangia chambers 12 in number, pore diameter 0.0179–0.027 mm.
Remarks.The diameter of the spicule and the diameter of the sporangial chambers are near the range of those described for Acicularia americana, 0.088–0.122 mm and 0.025–0.040 mm, respectively. This species may be placed in Terquemella (B. Granier, personal communication, June 25, 2010).
Stratigraphic range. This species is common in the Albian of Texas and Arizona (Johnson, 1969; Scott and González–León, 1991).
Genus Dissocladella Pia in Rama Rao and Pia, 1936
Dissocladella sp. cf. D. savitriae Rama Rao and Pia, 1936
Description. Thallus elongate, cylindrical, with subparallel margins; wall much thinner than thallus diameter; diameter of axial cavity about 60% of total thallus diameter. Inner primary branches numerous, vase–shaped, tapered inward toward axial cavity. Each primary branch divided into 2 to 4 secondary branches. Outer secondary branches typically slightly v–shaped outwards. Skeleton composed of blocky calcite spar, presumably replacing original aragonite.
Remarks. The dimensions and the large number of primary branches of the specimens from the Mal Paso Formation are very similar to those of D. savitriae, which is known from Maastrichtian – Danian beds in India. However, the Mexican species seems to have a greater number of primary branches than the Indian species and its primary branches are smaller. Also, the Mexican species is much larger than the Cretaceous species Dissocladella undulata (Raineri) (Johnson, 1969; Bassoullet et al., 1978; Kuss and Conrad, 1991).
Dimensions. Table 2. Measurements are given in mm.
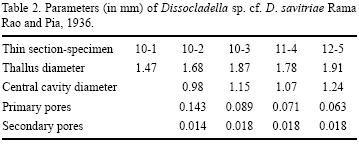
Stratigraphic range. This species occurs throughout a fairly thick interval of the Upper Albian calcareous upper member of the Mal Paso Formation, from unit MP9 to unit MP17, and it is common in unit MP14, the horizon which contains the main accumulation of the rudist Mexicaprina alata.
Incertae Sedis
Tube–within–a–tube fossil
Description. Elongate, curved tube with an internal tube or axis, circular to oval in cross section; diameter: outer tube, 0.232–0.214 mm, inner tube, 0.125 mm; walls of each tube composed of equant microgranular calcite and perforated by minute oval pores. Specimens which lack a central tube possess a wall composed of fine radial calcite fibers that originate from central axis; darker concentric growth rings closely spaced.
Remarks. Specimens of this taxon were recognized in thin section samples from units MP 9, MP10 and MP12 of the calcareous upper member of the Mal Paso Formation.
Order Foraminiferida Eichwald, 1830
Suborder Textulariina Delage and Hérouard, 1896
Superfamily Haplophragmiacea Eimer and Fickert, 1899
Family Nezzazatidae Hamaoui and Saint–Marc, 1970
Subfamily Nezzazatinae Hamaoui and Saint–Marc, 1970
Genus Nezzazata Omara, 1956
Type species. Nezzazata simplex Omara, 1956, p. 887.
Nezzazata sp. cf. N. isabellae Arnaud–Vanneau and Sliter, 1995
1995 Nezzazata isabellae Arnaud–Vanneau and Sliter, p. 552, figs. 7A–7D, pl. 2, figs. 11–24 (for comparison).
1995 Nezzazata isabellae Arnaud–Vanneau and Sliter, Arnaud–Vanneau and Premoli Silva, p. 206, pl. 2, figs. 1–3 (for comparison).
Description. Test low trochospiral, planoconvex; six to eight chambers per whorl; periphery rounded; diameter up to 0.60 mm. Short projections on inner edges of septa directed into preceding chamber. Wall microgranular calcite, imperforate.
Remarks. These planoconvex specimens are larger in diameter than the Aptian to Lower Albian Nezzazata isabellae Arnaud–Vanneau and Sliter, 1995, which has a biconvex test. The specimens from the Mal Paso Formation differ from Nezzazata gyra (Smout, 1956) and other Cenomanian species (Fleury, 1971) by their rounded periphery and unforked septal plate. This may be a new transitional species related to N. isabellae.
Stratigraphic range. All of the specimens are from unit MP18, near the top of the measured section of the calcareous upper member of the Mal Paso Formation. This morphotype of Nezzazata is part of the upper Albian microbiota assemblage which overlies species of the rudist Mexicaprina.
Superfamily Loftusiacea Brady, 1884
Family Cyclamminidae Marie, 1941
Diagnosis. Test enrolled, involute to uncoiled; wall agglutinated, outer layer imperforate, inner layer alveolar; septal structure differentiated from outer wall; aperture interio–marginal near septal base.
Remarks. Loeblich and Tappan (1988) and Banner and Highton (1990) placed the family Cyclamminidae in the superfamily Loftusiacea because of the alveolar inner wall layer. However, BouDagher–Fadel (2001) placed this family in the superfamily Lituolacea de Blainville.
Subfamily Buccicrenatinae Loeblich and Tappan, 1985
Diagnosis. Few chambers per whorl, wall and septa of early whorls may be solid, later stage with thin wall with large ovate alveolae; base of septa thickened with triangular chomata; chamber cavity open; aperture slit–like to zig–zag in coiling plane.
Genus Buccicrenata Loeblich and Tappan, 1949
Type species. Ammobaculites subgoodlandensis Vanderpool, 1933, p. 407.
Diagnosis. Test compressed, inner wall alveolae may bifurcate; septa solid or locally perforate; zig–zag aperture; other features of the subfamily.
Remarks. Buccicrenata is characterized by less than a complete whorl in its planispirally coiled stage and its chambers are arcuate reniform; the agglutinate outer wall has few tubular alveolae separated by large calcite and bioclastic grains; chamber septa are unperforated to locally perforated; base of septa next to preceding whorl are thickened triangle; the zigzag aperture spans the face of the last septum.
Everticyclammina Redmond, 1964, also in the subfamily Buccicrenatinae, is more lenticular and less compressed than Buccicrenata, has two whorls in its planispirally coiled stage and its chambers are wedge–shaped; the agglutinate outer wall has small alveolae and septa are unperforated; and the single simple aperture is wide (Loeblich and Tappan, 1988; Banner and Highton, 1990).
Pseudocyclammina Yabe and Hanzawa, 1926, in the subfamily Choffatellinae Maync, is characterized by its type species P. lituus (Yokoyama, 1890), which has two to three whorls in its planispirally coiled stage with narrow arcuate chambers; numerous closely spaced alveolae transect both the agglutinate wall and the septa; and the cribrate aperture fills the face of the last septum (Loeblich and Tappan, 1988).
Buccicrenata subgoodlandensis (Vanderpool, 1933)
1933 Ammobaculites subgoodlandensis Vanderpool, p. 407, pl. 49, figs. 1–3;
1944 Ammobaculites subgoodlandensis Vanderpool, Lozo, p. 540, pl. 1, figs. 2–3, pl. 4, fig. 1, text–figs. 15A–G.
1949 Buccicrenata subgoodlandensis (Vanderpool), Loeblich and Tappan, p. 253, pl. 47, figs. 5–15b.
1954 Lituola subgoodlandensis (Vanderpool), Frizzell, p. 66.
1965 Lituola subgoodlandensis (Vanderpool), Applin and Applin, pl. 1, fig. 11.
1966 Pseudocyclammina aff. hedbergi (Maync), Banner, pl. 3, figs. 5a–5b.
1970 Pseudocyclammina aff. hedbergi (Maync), Banner, pl. 5, figs. 7, 7a.
1985 Buccicrenata subgoodlandensis (Vanderpool), Loeblich and Tappan, p. 100, pl. 2, figs. 4–10.
1987 Buccicrenata subgoodlandensis (Vanderpool), Simmons and Hart, pl. 10.5, fig. 1.
1988 Buccicrenata subgoodlandensis (Vanderpool), Loeblich and Tappan, p. 99, pl. 96, figs. 1–9.
1990 Buccicrenata subgoodlandensis (Vanderpool), Banner and Highton, p. 12.
1991 Pseudocyclammina hedbergi Maync, Scott and González–León, p. 58;
1999 Pseudocyclammina hedbergi Maync, Scott and Finch, 1999, fig. 4D, E.
2001 Buccicrenata subgoodlandensis (Vanderpool), BouDagher–Fadel, p. 168–169, pl. 1, figs. 5–7.
Diagnosis. Test planispiral to uniserial, four to five hemispherical chambers in coil; uniserial part with one to three ellipsoidal chambers; simple terminal aperture.
Remarks. The five other species of Buccicrenata are described by BouDagher–Fadel (2001). Measurements of specimens from the Walnut Formation: length 0.71 to 4.73 mm, coil diameter 1.38 mm to 2.69 mm, test thickness 0.58 mm (Vanderpool, 1933; Loeblich and Tappan, 1949). The specimens from the Mal Paso Formation range in length from 0.98 mm to 1.79 mm, axial width 0.64 to 1.27 mm, and proloculus diameter 0.125 mm.
Stratigraphic range. The type specimens are from the Middle Albian Walnut Formation in southern Oklahoma (Vanderpool, 1933). The species ranges throughout the Albian in Tethyan sections from North America to the Middle East. In Texas the species occurs in the Lower Albian Glen Rose Formation and in the Lower to Upper Albian Walnut and Goodland formations of the Fredericksburg Group. In Florida the species was identified as Lituola subgoodlanensis where it occurs with Coskinolinoides texanus Keijzer in the Fredericksburg Group (Applin and Applin, 1965). In Mexico and Honduras the species was reported in Middle to Upper Albian strata as P. hedbergi by Scott and González–León (1991) and Scott and Finch (1999). In the United Arab Emirates and Oman it ranges throughout the Lower to Upper Albian Nahr Umr Formation (Simmons and Hart, 1987; Banner and Highton, 1990; BouDagher–Fadel, 2001). In Spain the species is reported from Upper Albian to Lower Cenomanian strata (González Fernández et al., 2004).
Superfamily Ataxophragmiacea Schwager, 1877
Family Cuneolinidae Saidova, 1981
Subfamily Cuneolininae Saidova, 1981
Genus Cuneolina d'Orbigny, 1839
Type species. Cuneolina pavonia d'Orbigny, 1846, p. 253.
Emended Diagnosis. "Test free, somewhat compressed parallel to the plane of biseriality and conical to flabelliform. The embryonic apparatus consists of a globular to oval protoconch covered by a deuteroconch that is divided by beams and rafters. The following biserial stage consists of broad, low chambers that are divided by radial beams extending from the outer wall toward the plane of biseriality and, sometimes, by horizontal rafters. The wall is agglutinated or microgranular and keriothecal. The aperture consists of a row of pores at the base of the septal face" (Arnaud–Vanneau and Sliter, 1995, p. 554).
Cuneolina parva Henson, 1948
1948 Cuneolina pavonia d'Orbigny var. parva var. nov. Henson, p. 624–627, pl. XIV, figs. 1–6, pl. XVII, figs. 7–12, pl. XVIII, figs. 12–14.
1991 Cuneolina walteri Cushman and Applin, 1947, not p. 30, pl. 10, figs. 4, 5a, b; Scott and González–León, p. 58, fig. 5O.
1995 Cuneolina parva Henson, Arnaud–Vanneau and Sliter, p. 554, pl. 4, figs. 6–9.
1995 Cuneolina parva Henson, Arnaud–Vanneau and Premoli Silva, p. 207.
1999 Cuneolina walteri Cushman and Applin, Scott and Finch, fig. 4A–C.
Diagnosis. Test moderate size, biserial flabelliform to triangular, apical angle generally less than 90 °. Wall agglutinated and composed of uniform micrite grains with small alveolae. Proloculus at test apex slightly eccentric and oval; up to twelve biserial arcuate concentric chambers expand from apex; chambers divided into chamberlets by radial septa that align with septa in adjacent chambers, but septa inserted between older septa in younger abapical chambers; one or two short lateral partitions project from outer wall of concentric chambers and insert a narrow concentric line between chamber septa; septal faces curved toward median wall between chambers.
Remarks. A stratigraphic succession of species ranges from Aptian to Maastrichtian and has been discriminated by test sizes, apical angles and sutures (Henson, 1948; Arnaud–Vanneau and Sliter, 1995). However, the two species reported most often are Cuneolina pavonia d'Orbigny, 1846, and Cuneolina parva Henson, 1948 (Arnaud–Vanneau and Sliter, 1995; Arnaud–Vanneau and Premoli Silva, 1995). Cuneolina pavonia is flabelliform and tends to be slightly larger and have a greater apical angle than C. parva, which is conical to flabelliform (Table 3; Figure 5, graphs a, b). The mean apical angle of paratypes of C. parva is 48 ° and ranges from 18 ° to 96 ° (Henson, 1948). The chambers of C. pavonia are wider than those of C. parva and C. pavonia has two to three transverse rafters in the chambers compared to one to two in C. parva (Arnaud–Vanneau and Sliter, 1995; Arnaud Vanneau and Premoli Silva, 1995). Cuneolina parva is reported from Albian – Cenomanian strata and C. pavonia from uppermost Albian to Santonian. The Aptian species, Cuneolina sliteri Arnaud Vaneau and Premoli Silva, 1995, has a small triangular to conical test and lacks the horizontal rafters in its first chambers.

The Middle Cenomanian Florida species Cuneolina walteri Cushman and Applin, 1947 (pl.10, figs. 4, 5a, 5b) is also illustrated by Applin and Applin (1965, pl. 2, fig. 2). This name was used for specimens in Mexico (Scott and González–León, 1991) and in Honduras (Scott and Finch, 1999). The dimensions of the types of C. walteri are in the range of topotypes of the older name C. pavonia (Figure 5, graphs A, B) and if the internal structure of C. walteri is like that of C. pavonia, then the former would be a junior synonym of the latter. The sizes of the Mexican and Honduran specimens are in the size range of C. parva and overlap with the smaller specimens of C. pavonia. However, measurements of Cuneolina specimens taken from thin sections tend to be incomplete because most specimens are unoriented and the cross sections are oblique to the growth axes. Thus, distinguishing species of Cuneolina by sizes of thin section specimens has some degree of uncertainty.
Cuneolina conica d'Orbigny, 1846, in the Coniacian– Santonian is a narrow cylindrical species and Cuneolina fleuriausiana d'Orbigny, 1839, has a very acute apical angle (Henson, 1948). Cuneolina cylindrica Henson, 1948, in the Maastrichtian has an apical angle of 7 ° to 14 °. Clearly the sizes and apical angles do not readily separate these species and a study of the types and collections of free specimens is needed to clarify the taxonomy of Cuneolina species.
Stratigraphic range. The type specimens of C. pavonia are from the Cenomanian of France and the species ranges from uppermost Albian to Santonian in the European and North African Tethys (Arnaud–Vanneau and Premoli Silva, 1995). The types of C. parva are from Egypt in presumed Albian strata, not Santonian as stated by Henson (Arnaud–Vanneau and Premoli Silva, 1995). This species is known from Albian to Cenomanian strata in the Tethys, including Upper Albian strata in Mexico and Honduras. The types of C. walteri are from the lower member of the Atkinson Formation in the subsurface of Florida, which is correlated with the Middle Cenomanian Woodbine Group in north Texas (Applin and Applin, 1965). Specimens of C. parva were recognized in most of the sampled beds of the calcareous upper member of the Mal Paso Formation.
Family Coskinolinidae Moullade, 1965
Genus Pseudolituonella Marie, 1955
Type species. Pseudolituonella reicheli Marie, 1955.
Pseudolituonella sp.
Description. Test uniserial, conical, with a trochospiral proloculus, two juvenile ovate chambers and four arcuate chambers. Test length 0.52 mm, width 0.34 mm. Wall of test microcrystalline calcite and solid. Proloculus a single oval chamber with a thick prismatic wall; inner diameter 0.053 mm, outer diameter 0.120 mm.
Remarks. The species description is based on a specimen from the calcareous upper member of the Mal Paso Formation. The specimen is much smaller than and differs from Pseudolituonella reicheli by the absence of tubular pillars on the adapical side of the apertures. It is very similar to a specimen in the Upper Albian section in the Pyrenees identified as Pseudolituonella sp.? (Peybernes, 1976, pl. 34, fig. 12), which is 1.62 mm in length and 0.85 mm in width. The older and larger Pseudolituonella gavonensis Foury, 1968, has a streptospiral early stage.
Stratigraphic range. According to Loeblich and Tappan (1988) the genus ranges from Cenomanian to Campanian in the Mediterranean part of the Tethys. However, P. gavonesis is a Barremian species. In Europe, Pseudolituonella sp.? has been reported with the Upper Albian Neorbitolinopsis conulus (Douvillé) (Peybernes, 1976). The Mexican specimen is from unit MP10 of the calcareous upper member of the Mal Paso Formation.
Superfamily Textulariacea Ehrenberg, 1838
Family Chrysalidinidae Neagu, 1968
Subfamily Chrysalidininae Neagu, 1968
Genus Praechrysalidina Luperto Sinni, 1979
Type species. Praechrysalidina infracretacea Luperto Sinni, 1979.
Praechrysalidina sp.
Description. Test small, conical, with overlapping apertural faces. Test length 0.119–0.385 mm, width 0.098–0.210 mm. No interior marginal flaps or other structures present. Proloculus not observed.
Remarks. The species description is based on specimens from the calcareous upper member of the Mal Paso Formation. Banner et al. (1991) placed Praechrysalidina in the family Chrysalidinidae Neagu because of its inferred phylogenetic relationship with Chrysalidina rather than in the family Ataxophragmiidae Schwager, as did Loeblich and Tappan (1988). The Mal Paso specimens are similar to the juvenile portion of a specimen from the Albian of Iraq (Banner et al., 1991, fig. 9). The Mexican specimens do not show the characters of Praechrysalidina infracretacea and they are smaller; additional material is needed in order to make a more detailed species description and a more accurate species determination.
Stratigraphic range. The species occurs in units MP6 and MP10 of the calcareous upper member of the Mal Paso Formation. The overall stratigraphic range of the species referred to this genus is Valanginian to Albian in Tethyan strata (Banner et al., 1991).
Suborder Globigerinina Delage and Hérouard, 1896
Superfamily Rotaliporacea Sigal, 1958
Family Rotaliporidae Sigal, 1958
Subfamily Ticinellinae Longoria, 1974
Genus Rotalipora Brotzen, 1942
Type species. Rotalipora turonica Brotzen, 1942 (= Globorotalia cushmani Morrow, 1934).
Rotalipora appenninica (Renz, 1936)
1936 Globotruncana appenninica Renz, p. 20, pl.6, figs. 1–11; pl. 7, fig. 1; pl. 8, fig. 4; text–figs. 2, 7a.
1977 Rotalipora appenninica (Renz, 1936), Masters, p. 497–501, pl. 30, figs. 1, 3, not fig. 2 (prior synonymy provided).
2002 Rotalipora appenninica (Renz, 1936), Premoli Silva and Sliter, p. 71, pl. 8, figs. 1a–1c, 2a–c, 3a–c, pl. 9, figs. 1a–1c, 2a–c, 3a–c.
Description. Test chambers trapezoidal in outline; keel well–developed along peripheral margin. Dimensions of two specimens observed in thin section M10: test diameter 0.378 mm, height 0.126 mm; and test diameter 0.252 mm, height 0.105 mm.
Remarks. The brief description is based on two specimens observed in thin section M10 from unit MP18. The overall form of the Mexican specimens is most similar to that of R. appenninica, except the wall is thicker than normal, especially in the early whorls (Mark Leckie and Brian Huber, personal communications, December 2009). The thickened wall may be the result of recrystallization.
Stratigraphic range. The Mexican specimens are from unit MP18, near the top of the measured section of the calcareous upper member of the Mal Paso Formation. The full range of R. appenninica is Upper Albian to Lower Cenomanian. The Rotalipora appenninica Zone is defined as the interval above the first occurrence (FO) of the nominate species and the FO of Rotalipora brotzeni (Sigal) and it correlates with latest Albian ammonite zones (Premoli Silva and Sliter, 2002).
ACKNOWLEDGMENTS
The authors thank the two anonymous reviewers for their useful comments on the manuscript; Dr. Francisco Vega for the Spanish version of the abstract and key words; and Dra. Maria del Carmen Perrilliat for assistance with cataloging the illustrated specimens in the IGM collections.
REFERENCES
Applin, P.L., Applin, E.R., 1965, The Comanchean Series and associated rocks in the subsurface in central and south Florida: United States Geological Survey Professional, 447, 84 pp. [ Links ]
Amédro, F., 2008, Support for a Vraconian Stage between the Albian sensu stricto and the Cenomanian (Cretaceous System): Carnets de Géologie / Notebooks on Geology, Brest, Memoir 2008/02 (CG2008_M02), 83 pp. [ Links ]
Amédro, F., Robaszynski, F., 2008, Zonation by ammonites and foraminifers of the Vraconian–Turonian interval: A comparison of the Boreal and Tethyan domains (NW Europe / Central Tunisia): Carnets de Géologie / Notebooks on Geology, Memoir 2008/02 (CG2008_L02), 5 pp. [ Links ]
Archiac, A., d', 1843, Description géologique du départment de l'Aisne: Mémoires de la Société Géologique de France, 5(3), 129–420. [ Links ]
Arnaud–Vanneau, A., Sliter, W.V., 1995, Early Cretaceous shallow–water benthic foraminifers and fecal pellets from Leg 143 compared with coeval faunas from the Pacific Basin, Central America, and the Tethys, in Winterer, E.L., Sager, W.W., Firth, J.V., Sinton, J.M. (eds.), Proceedings of the Ocean Drilling Program, Scientific Results: Texas, College Station, Ocean Drilling Program, 143, 537–564. [ Links ]
Arnaud–Vanneau, A., Premoli Silva, I., 1995, Biostratigraphy and systematic description of benthic foraminifers from Mid–Cretaceous shallow–water carbonate platform sediments at sites 878 and 879 (MIT and Takuyo–Daisan Guyots, in Haggerty, J.A., Premoli Silva, I., Rack, F., McNutt, M.K. (eds.), Proceedings of the Ocean Drilling Program, Scientific Results: Texas, College Station, Ocean Drilling Program, 144, 199–219. [ Links ]
Banner, F.T., 1966, Morfologiya, klassificatsiya i stratigraficheskoe znachenie spirotsiklinid: Voprosi Micro–paleontologii, 10, 201–234. [ Links ]
Banner, F.T., 1970, A synopsis of the Spirocyclinidae: Revista Española Micropaleontologia, 2(3), 243–290. [ Links ]
Banner, F.T., Highton, J., 1990, On Everticyclammina Redmond (Foraminifera), especially E. kelleri (Henson): Journal of Micropaleontology, 9(1), 1–14. [ Links ]
Banner, F.T., Simmons, M.D., Whittaker, J.E., 1991, The Mesozoic Chrysalidiinidae (Foraminifera, Textulariacea) of the Middle East: the Redmond (Aramco) taxa and their relatives: Bulletin British Museum of Natural History (Geology), 47, 101–152. [ Links ]
Banner, F.T., Simmons, M.D., 1994, Calcareous algae and foraminifera as water–depth indicators: an example from the Early Cretaceous carbonates of northeast Arabia, in Simmons, M.D., (ed.), Micropalaeontology and hydrocarbon exploration in the Middle East: London, Chapman and Hall, 243–252. [ Links ]
Bassoullet, J.P., Bernier, P., Conrad, M., Deloffre, R., Jaffrezo, M., 1978, Les algues Dasycladales du Jurassique et du Crétacé: Geobios, mémoire spécial 2, 330 pp. [ Links ]
BouDagher–Fadel, M.K., 2001, The taxonomy and evolution of the foraminiferal genus Buccicrenata Loeblich and Tappan: Micropaleontology, 47(2), 168–172. [ Links ]
Brady, H.B., 1884, Report on the foraminifera collected by H.M.S. Challenger during the years 1873–1876 (on line), in Report on the Scientific Results of the Voyage of H.M.S. Challenger during the years 1873–1876, v. 9, Zoology, part 22: Bossard, D.C., publicación mayo de 2004, http://www.19thcenturyscience.org/HMSC/HMSC–Reports/Zool–22/htm/doc.html [ Links ]
Brotzen, F., 1942, Die Foraminiferengattung Gavelinella nov. gen. und die Systematik der Rotaliiformes: Årsbok Sveriges Geologiska Undersökning, 36(8), 1–60. [ Links ]
Buitrón–Sánchez, B.E., Pantoja–Alor, J., 1994, Esponjas perforantes de moluscos del Cretácico Temprano en la región centrooccidental de México: Revista Mexicana de Ciencias Geológicas, 11(2), 222–231. [ Links ]
Buitrón–Sánchez, B.E., Carrillo–Martinez, M., Aguilera–Franco, N., 1995, A Middle Albian biota (algae, foraminifera and gastropoda) from Ahuacatlán, State of Querétaro, Mexico: Revista Mexicana de Ciencias Geológicas, 12(2), 145–156. [ Links ]
Buitrón–Sánchez, B.E., Pantoja–Alor, J., 1996, Albian gastropods of the rudist–bearing Mal Paso Formation of the Chumbitaro region, SW Mexico (resumen), in Quatrième Congrès International sur les Rudistes (Fourth International Conference on Rudists), Université de Provence, Centre de Sédimentologie–Paléontologie : Marseille, France, p. 3. [ Links ]
Buitrón–Sánchez, B.E., Pantoja–Alor, J., 1998, Albian gastropods of the rudist–bearing Mal Paso Formation, Chumbítaro region, Guerrero, Mexico: Revista Mexicana de Ciencias Geológicas, 15(1), 14–20. [ Links ]
Centeno–García, E., 1994, Tectonic Evolution of the Guerrero Terrane, Western Mexico: University of Arizona, Ph.D. Dissertation, 220 pp. [ Links ]
Coogan, A. H., 1973, Nuevos rudistas del Albiano y Cenomaniano de Mexico y del sur de Texas: Revista del Instituto Mexicano del Petróleo, 5, 51–83. [ Links ]
Cushman, J.A., Applin, E.R., 1947, Two new species of Lower Cretaceous foraminifera from Florida: Cushman Laboratory for Foraminiferal Research Contribution, 23(291), 29–30. [ Links ]
Delage, Y., Hérouard, E., 1896, Traité de Zoologie Concrète, vol. 1, La Cellule et les Protozoaires: Paris, Schleicher Frères, 584 pp. [ Links ]
Dragastan, O., 1970, New species of Dasycladaceae (calcareous algae) in the Lower Cretaceous of the eastern Carpathians (Rumania): Review of Palaeobotany and Palynology, 10(2), 117–129. [ Links ]
Dragastan, O., 1981, Mesozoic Dasycladaceae from Romania: Distribution and biostratigraphical importance: Facies, 4, 165–195. [ Links ]
Ehrenberg, C.G., 1838, Über dem blossen Auge unsichbare Kalkthierchen und Kieselthierchen als Hauptbestandtheile der Krediegebirge: Bericht über die zu Bekanntmachung geeigneten Verhandlungen de Königlichen Preussischen Akademie der Wissenschaften zu Berlin, 1938, p. 192–200. [ Links ]
Eichwald, C.E. von, 1830, Zoologia specialis, v. 2: Vilnae, D.E. Eichwaldus, 323 pp. [ Links ]
Eimer, G.H.T., Fickert, C., 1899, Die Artbildung und Verwandtschaft bei den Foraminiferen. Entwurf einer natürlichen Eintheilung derselben. Zeitschrift für Wissenschaftliche Zoologie, v. 65, p. 599–708. [ Links ]
Elliott, G.F., 1956, Further records of fossil calcareous algae from the Middle East: Micropaleontology, 2(4), 327–334. [ Links ]
Elliott, G.F., 1957, Algues calcaires codiacées fossiles d'Iraq, nouvelles et peu connues: Bulletin de la Société Géologique de France, 6(6), 789–795. [ Links ]
Elliott, G.F., 1963, Problematical microfossils from the Cretaceous and Palaeocene of the Middle East: Palaeontology, 6(2), 293–300. [ Links ]
Filkorn, H.F., 2002a, A new species of Mexicaprina (Caprinidae, Coalcomaninae) and review of the age and paleobiogeography of the genus: Journal of Paleontology, 76(4), 672–691. [ Links ]
Filkorn, H.F., 2002b, Rudist bivalves and reef corals in the Lower Cretaceous of southwestern Mexico: tiered coexistence, competition for the same ecologic niche, and facies superposition (resumen), in Geological Society of America, Abstracts with Programs, 34(6), 538. [ Links ]
Filkorn, H.F., Pantoja–Alor, J., 2004, A new Early Cretaceous coral (Anthozoa; Scleractinia; Dendrophylliina) and its evolutionary significance: Journal of Paleontology, 78(3), 501–512. [ Links ]
Filkorn, H.F., Pantoja–Alor, J., 2009, Cretaceous corals from the Huetamo region, Michoacán and Guerrero, southwestern Mexico: Universidad Nacional Autónoma de México, Instituto de Geología, Boletín 116, 168 pp. [ Links ]
Fleury, J.J., 1971, Le Cénomanian a foraminifères benthoniques du massif du Varassove (zone du Gavrovo, Akarnanie, Grece continentale): Revue de Micropaléontologie, 14, 181–194. [ Links ]
Foury, G., 1968, Le Crétacé inférieur des Alpilles contribution a L'étude stratigraphique et micropaléontologique: Geobios, 1, 119–164. [ Links ]
Freydier, C., Lapierre, H., Briqueu, L., Tardy, M., Coulon, C., Martínez–Reyes, J., 1997, Volcaniclastic sequences with continental affinities within the Late Jurassic – Early Cretaceous Guerrero intra–oceanic arc terrane (western Mexico): Journal of Geology, 105(4), 483–502. [ Links ]
Frizzell, D.L., 1954, Handbook of Cretaceous Foraminifera of Texas: University of Texas, Bureau of Economic Geology, Report of Investigation No. 22, 232 pp. [ Links ]
Frollo, M.M., 1938, Sur un Nouveau genere de Codiacée du Jurassique Supérieur des Carpates Orientales: Société Géologique de France Bulletin, 5(8), 269–271. [ Links ]
García–Barrera, P., Pantoja–Alor, J., 1991, Equinoides del Albiano tardío de la Formación Mal Paso, de la región Chumbítaro, estados Guerrero y Michoacán, México: Revista de la Sociedad Mexicana de Paleontología, 4, 23–41. [ Links ]
González Fernández, B., Menéndez Casares, E., Gutiérrez Claverol, M., García–Ramos, J.C., 2004, Litoestratigrafía del sector occidental de la cuenca cretácica de Asturias: Trabajos de Geología, Universita de Oviedo, 24, 43–80. [ Links ]
G Grötsch J., Schroeder, R., Noé, S., and Flügel, E., 1993, Carbonate plattforms as recorders of high–amplitude eustatic sea–level fluctuations: the late Albian appenninica–event: Basin Research, 5, 197–212. [ Links ]
Hamaoui, M. and Saint–Marc, P., 1970, Microfaunes et microfaciès du Cénomanien du Proche–Orient: Bulletin Centre Recherches Pau–SNPA, v. 4, p. 257–352. [ Links ]
Hauck, F., 1885, Die Meeresalgen, in Rabenhorst, L. (ed.), Kryptogamen.― Flora von Deutschland, Österreich und der Schweiz: Leipzig, Kummer, 334–410. [ Links ]
Henson, F.R.S., 1948, New Trochamminidae and Verneuilinidae from the Middle East: Annals and Magazine of Natural History, 11(14), 605–636. [ Links ]
Johnson, J.H., 1969, A review of the Lower Cretaceous algae: Colorado School of Mines, Professional Contributions, 6, 180 pp. [ Links ]
Kauffman, E.G., Johnson, C.C., 1988, The morphological and ecological evolution of middle and Upper Cretaceous reef–building rudistids: Palaios, 3(2), 194–216. [ Links ]
Konishi, K., Epis, R.C., 1962, Some Early Cretaceous calcareous algae from Chochise County, Arizona: Micropaleontology, 8(1), 67–76. [ Links ]
Kuss, J., 1986, Upper Cretaceous algae from the Eastern Desert of Egypt: Neues Jahrbuch für Geologie and Paläontologie, Monatshefte, 1986, 223–238. [ Links ]
Kuss, J., Conrad, M.A., 1991, Calcareous algae from Cretaceous carbonates of Egypt, Sinai, and southern Jordan: Journal of Paleontology, 65(5), 869–882. [ Links ]
Kützing, F.T., 1843, Phycologia generalis: Leipzig, Perthes, 458 pp. [ Links ]
Loeblich, A.R.Jr., Tappan, H., 1949, Foraminifera from the Walnut Formation (Lower Cretaceous) of Northern Texas and Southern Oklahoma: Journal of Paleontology, 23(3), 245–266. [ Links ]
Loeblich, A.R.Jr., Tappan, H., 1985, Some new and redefined genera and families of agglutinated foraminifera I: Journal of Foraminiferal Research, 15(2), 91–104. [ Links ]
Loeblich, A.R., Jr., Tappan, H., 1988, Foraminiferal genera and their classification: New York, Van Nostrand Reinhold Company, 970 pp. [ Links ]
Longoria, J.F., 1974, Stratigraphic, morphologic and stratigraphic studies of Aptian planktonic foraminiferida: Revista Española de Micropaleontologia, Numero extraordinario, 1–107. [ Links ]
Lozo, F.E.Jr., 1944, Biostratigraphic relations of some North Texas Trinity and Fredericksburg (Comanchean) foraminifera: American Midlands Naturalist, 31(3), 513–382. [ Links ]
Luperto Sinni, E.,1979, Praechrysalidina infracretacea n. gen., n. sp. (Foraminiferida) del Cretaceo Inferiore delle Murge Baresi: Bari, Università degli Studi di Bari, Studi Geologici e Morphologici sulla Regione Pugliese V, Istituto di Geologia e Paleontologia, 1–16. [ Links ]
Mann, P., 1999, Caribbean sedimentary basins: classification and tectonic setting from Jurassic to Present, in Mann, P. (ed.), Caribbean Basins. Sedimentary Basins of the World. 4: Amsterdam, Elsevier Science B.V., 3–31. [ Links ]
Marie, P., 1941, Les foraminifères de la Craie à Belemnitella mucronata du Bassin de Paris: Mémoires du Museum Nationale d'Histoire Naturelle, 12(1), 1–296. [ Links ]
Marie, P., 1955, Quelques genres nouveaux de foraminifères du Crétacé à facies récifal: Compte Rendus 19th Congrès Géologique International, Alger 1952, 13(15), 117–124. [ Links ]
Masters, B.A., 1977, Mesozoic planktonic foraminifera, in Ramsay, A.T.S. (ed.), Oceanic Micropaleontology, vol. 1: London, Academic Press, 301–731. [ Links ]
Morrow, A.L., 1934, Foraminifera and ostracoda from the Upper Cretaceous of Kansas: Journal of Paleontology, 8, 186–205. [ Links ]
Moullade, M., 1965, Contribution au problème de la classification des Orbitolinidae (Foraminiferida, Lituolacea): Compte Rendus Hebdomadaires des Séances, Académie des Sciences, Paris, 260, 4031–4034. [ Links ]
Neagu, T., 1968, Andersenia rumana n. gen., n. sp., and some taxonomic observations on the subfamily Valvulininae: Contributions from the Cushman Foundation for Foraminiferal Research, 19, 120–122. [ Links ]
Omaña–Pulido, L., Pantoja–Alor, J., 1998, Early Aptian benthic foraminifera from the El Cajón Formation, Huetamo, Michoacán, SW México: Revista Mexicana de Ciencias Geológicas, 15(1), 64–72. [ Links ]
Omara, S., 1956, New foraminifera from the Cenomanian of Sinai, Egypt: Journal of Paleontology, 30, 883–890. [ Links ]
d'Orbigny, A., 1839, Foraminifères, in Sagra, R. (ed.), Histoire physique, politique et naturelle de l'île de Cuba: Paris, Arthus Bertrand. [ Links ]
d'Orbigny, A., 1846, Foraminifères fossiles du Bassin Tertiaire de Vienne (Autriche): Paris, Gide et Comp, 4. [ Links ]
Palma, R.M., Lo Forte, G.L., Medhli, M., Piethé, R.D., 2005, High frequency subtidal–peritidal cycles of the Callovian Calabozo Formation (Neuquén Basin, western Argentina): preliminary approach: Geologica Acta, 3, 119–132. [ Links ]
Pantoja–Alor, J., 1959, Estudio geológico de reconocimiento de la región de Huetamo, Estado de Michoacán: Consejo de Recursos Naturales No–Renovables (Mexico), Boletín 50, 36 pp. [ Links ]
Pantoja–Alor, J., 1992, La Formación Mal Paso y su importancia en la estratigrafía del sur de México: Sociedad Geológica Mexicana, Convención Geológica Nacional, 11, 121–123. [ Links ]
Peybernès, B., 1976, Le Jurassique et le Crétacé inférieur des Pyrénées Franco–Espagnoles, entre la Garonne et la Méditerranée: Tulouse, Thèse de Doctorat Sciences Naturelles, 459 p. [ Links ]
Pia, J., 1936, Calcareous Green algae from the Upper Cretaceous of Tripoli (North Africa): Journal of Paleontology, 10(1), 3–13. [ Links ]
Pfender, J., 1936, Sur un organisme constucteur des calcaires cretaces et nummulitiques, Pseudolithothamnium album, n.g., nov. sp.: Bulletin Societe geologique France, 5, 303–308. [ Links ]
Premoli Silva, I., Sliter, W.V., 2002, Practical manual of Cretaceous planktonic Foraminifera: Perugia, University of Perugia, Dipartimento di Scienze della Terra, 462 pp. [ Links ]
Rama Rao, L., Pia, J., 1936, Fossil algae from the uppermost Cretaceous beds (the Niniyur Group) of the Trichinopoly District, S. India: Calcutta, Palaeontologica Indica, New Series, 21(4), 1–49. [ Links ]
Redmond, C.D., 1964, Lituolid foraminifera from the Jurassic and Cretaceous of Saudi Arabia: Micropaleontology, 10(4), 405–414. [ Links ]
Renz, O., 1936, Stratigraphische und mikropaläontologische Unterschung der Scaglia (Obere Kreide–Tertiär) im zentralen Apennin: Eclogae geolgicae Helvetiae, 29, 1–149. [ Links ]
Saidova, Kh.M., 1981, O sovremennom sostoyanii sistemy nadvidovykh taksonov Kaynozoyskikh bentosnykh foraminifer [On an up–to–date system of supraspecific taxonomy of Cenozoic benthonic foraminifera]: Moscow, Institut Okeanologii P.P. Shirshova, Akademiya Nauk SSR. [ Links ]
Saunders, G.W., Hommersand, M.H., 2004, Assessing red algal supraordinal diversity and taxonomy in the context of contemporary systematic data: American Journal of Botany, 91, 1494–1507. doi:10.3732/ajb.91.10.1494. [ Links ]
Schwager, C., 1877, Quadro del proposto sistema di classificazione dei formaniferi con guscio: Bolletino R. Comitato Geologico d'Italia, 8, 18–27. [ Links ]
Scott, R.W., 1995, Global environmental controls on Cretaceous reefal ecosystems: Palaeogeography, Palaeoclimatology, Palaeoecology, 119(1–2), 187–199. [ Links ]
Scott, R.W., 2009, Uppermost Albian Biostratigraphy and Chronostratigraphy: Carnets de Géologie / Notebooks on Geology, Article 2009/03 (CG2009_A03), 16 pp. [ Links ]
Scott, R.W., Finch, R.C., 1999, Cretaceous Carbonate Biostratigraphy and Environments in Honduras, in Mann, P. (ed.), Caribbean Basins. Sedimentary Basins of the World, 4: Amsterdam, Elsevier Science B.V., 151–165. [ Links ]
Scott, R.W., González–León, C., 1991, Paleontology and biostratigraphy of Cretaceous rocks, Lampazos area, Sonora, Mexico: Geological Society of America Special Paper 254, 51–67. [ Links ]
Scott, R.W., Filkorn, H.F., 2007, Barremian–Albian Rudist Zones, U.S. Gulf Coast, in Scott, R.W. (ed.), Cretaceous Rudists and Carbonate Platforms: Environmental Feedback: SEPM Special Publication 87, 167–180. [ Links ]
Scott, R.W., Benson, D.G., Morin, R.W., Shaffer, B.L., Oboh–Ikuenobe, F.E., 2003. Integrated Albian–Lower Cenomanian Chronostratigraphy and Paleoecology, Texas and Mexico: Perkins Memorial Volume: GCSSEPM Foundation, Special Publications in Geology, No. 1, CD book, p. 277–334. [ Links ]
Sigal, J., 1958, La classification actuelle des familles de foraminifères planctoniques du Crétacé: Compte Rendu des Séances, Société Géologique de France, 1958, 262–265. [ Links ]
Simmons, M.D., Hart, M.B., 1987, The biostratigraphy and microfacies of the Early to mid–Cretaceous carbonates of Wadi Mi'aidin, Central Oman Mountains, in Hart, M.B. (ed.), Micropalaeontology of Carbonate Environments: Chichester, Ellis Horwood Ltd., 176–207. [ Links ]
Sliter, W.V., Baker, R.A., 1972, Cretaceous bathymetric distribution of benthic foraminifers: Journal of Foraminiferal Research, 2, 167–183. [ Links ]
Smout, A.H., 1956, Three new Cretaceous genera of Foraminifera related to the Ceratobuliminidae: Micropalentology, 2, 335–348. [ Links ]
Sokac, B., Nikler, L., 1973, Calcareous algae from the Lower Cretaceous of the environs of Niksic, Crna Gora (Montenegro): Paleontologie jugoslavica, Zagreb, 13, 1–57. [ Links ]
Tronchetti, G., 1984, Influence du substrat et de la salinité dans la répartition des foraminifères benthiques: example du Crétacé moyen et supérieur de Provence (SE France): Pau and Bordeaux, Benthos '83, 2nd International Symposium on Benthic Foraminifera, 567–571. [ Links ]
Vanderpool, H.C., 1933, Upper Trinity microfossils from southern Oklahoma: Journal of Paleontology, 7(4), 406–411. [ Links ]
Wray, J.L., 1978, Calcareous algae, in Haq, B.U., Boersma, A. (eds.), Introduction to Marine Micropaleontology: New York, Elsevier, 171–187. [ Links ]
Yabe, H., Hanzawa, S., 1926, Choffatella Schlumberger and Pseudocyclammina, a new genus of arenaceous foraminifera: Sendai, Science Reports of the Tohoku University, series 2, Geology 9, 9–11. [ Links ]
Yokoyama, M. , 1890, Foraminiferen aus dem Kalksteine von Torinosu und Kompira: Wien, Denkschriften der Kaiserlichen Akademie der Wissenschaften, Mathematisch–Naturwissenschaftliche Classe, 57, 26–27. [ Links ]

