










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
 Citado por SciELO
Citado por SciELO Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista mexicana de ciencias geológicas
versión On-line ISSN 2007-2902versión impresa ISSN 1026-8774
Rev. mex. cienc. geol vol.27 no.2 Ciudad de México ago. 2010
Nuevas especies de Quercus (Fagaceae) en el Plioceno de Santa María Amajac, Hidalgo, México
New Quercus (Fagaceae) species from the Pliocene of Santa María Amajac, Hidalgo, Mexico
María Patricia Velasco–de León* y Erika Lourdes Ortiz–Martínez
Facultad de Estudios Superiores Zaragoza, Universidad Nacional Autónoma de México Av. Guelatao 66, Col. Ejercito de Oriente, Iztapalapa, México, D.F. * pativel@servidor.unam.mx
Manuscrito recibido: Septiembre 14, 2009.
Manuscrito corregido aceptado: Abril 24, 2010.
Manuscrito aceptado: Mayo 7, 2010.
RESUMEN
Se describen siete nuevas especies fósiles de la familia Fagaceae para México, utiliiando la arquitectura foliar: Quercus amajensis, Q. cevalli, Q. curvensis, Q. grandensis, Q. hidalgensis, Q. ovatus y Q. rugosus. Las impresiones de hojas fósiles fueron colectadas en las localidades de Sanctorum y Los Baños, Hidalgo, dentro de la Formación Atotonilco El Grande. El análisis fenético realiiado permite concluir que los caracteres importantes que definen a las nuevas especies son: recorrido de las venas terciarias, venación de quinto orden, forma y tamaño de areolas, presencia de venas intersecundarias y recorrido de la vena secundaria dentro del diente. Estos resultados apoyan la idea de la importancia del Cinturón Volcánico Transmexicano como área de especiación.
Palabras clave: arquitectura foliar, taxonomía, Quercus, encino, Plioceno, Hidalgo, México.
ABSTRACT
Using foliar architecture seven new fossil species of the Fagaceae Family are described for Mexico: Quercus amajensis, Q. cevalli, Q. curvensis, Q. grandensis, Q. hidalgensis, Q. ovatus and Q. rugosus. Fossil leaf impressions were collected in the areas of Sanctorum and Los Baños, in the Atotonilco El Grande Formation in Hidalgo. The phenetic analysis performed allow us to conclude which characters are important in defining the new species: ramification of the tertiary veins, fifth order venation, shape and size of the areole, the presence of inter–secondary veins and secondary veins ramification within the tooth. These results support the importance of the Trans–Mexican Volcanic Belt as a speciation area.
Key words: foliar architecture, taxonomy, Quercus, oak, Pliocene, Hidalgo and Mexico.
INTRODUCCIÓN
El Cinturón Volcánico Transmexicano (CVT) ha jugado un papel importante en la diversificación de diferentes grupos de plantas. En esta zona se encuentran enclavadas las localidades fósiles de la región de Santa María Amajac (SMA) donde, en el Plioceno, se localizaba un pequeño lago intermontano (Arellano et al., 2005). En las localidades que lo circundaban (Los Baños y Sanctorum) se han recolectado más de 2000 hojas de angiospermas representando aproximadamente 35 morfotipos, siendo los más abundantes aquéllos con similitud a hojas de encinos. Las hojas del género Quercus se caracterizan por ser alternas, generalmente pecioladas, de láminas simples, enteras o divididas en el margen. La venación primaria es pinnada, la secundaria va de semicaspedódroma, a craspedódroma con dientes mucronados y/o aristados, las venas secundarias entran al diente de forma central o exmedial, en las hojas enteras se presenta venación broquidódroma. Con venas intersecundarias simples y compuestas, que divergen de la vena media en ángulos iguales o mayores que el de las venas secundarias. La venación de alto orden esta bien desarrollada (cinco niveles son comunes) al igual que las areolas, estas ultimas por lo regular son cuadrangulares, medianas y pequeñas, con venillas ramificadas.
La familia Fagaceae se distribuye en todo el mundo con excepción de África tropical. Quercus es el género más grande de esta familia. Actualmente se distribuye principalmente en el hemisferio norte e incluye aproximadamente 500 especies en todo el mundo. En México, el número exacto de especies se desconoce; Trelease (1924) calcula 253 especies para México, mientras que estimaciones de Nixon (1993) arrojan entre 135 y 150 especies, y por último Valencia (2004) propone un listado de 161 especies, de las cuales 109 son endémicas. La alta diversidad de este género en México ha fundamentado que se le proponga, al igual que a Asia (125 spp), como uno de los centros de diversificación del grupo.
El CVT ha jugado un papel importante en la diversificación de este y otros géneros de plantas actuando como barrera o área de especiación (p. ej., Bursera; Espinosa et al., 2006). Históricamente se ha considerado a esta zona como una provincia morfotectónica muy importante desde el punto de vista de riqueza de especies (Ferrusquía–Villafranca, 1993). En la actualidad, en el CVT se distribuyen 36 especies de Quercus, de las cuales 29 son endémicas de México (Valencia, 2007). La riqueza de esta zona abarca también la flora fósil, ya que se han ubicado afloramientos importantes de angiospermas en el municipio de Atotonilco El Grande en donde el género dominante es precisamente Quercus.
La taxonomía del género es complicada debido a la gran variedad morfológica que presentan sus individuos por lo que diferentes autores han buscado caracteres foliares útiles en la delimitación de especies. Hickey (1973) utiliza el término arquitectura foliar para describir la expresión externa de la estructura foliar, incluyendo tipo de venación, configuración marginal, forma de la hoja y posición de las glándulas. El uso de la arquitectura foliar ha demostrado que los diversos taxones de angiospermas tienen hojas que poseen modelos de organización arquitectónica consistentes. En México son pocos los trabajos que han utilizado la arquitectura foliar como una herramienta útil para delimitar a las especies de este género. Valencia (1994) estudia Q. affinis Scheidweiler y Q. laurina Humboldt y Bonpland y encuentra caracteres que permiten diferenciarlas. Otros autores que han realizado estudios en este tema son: Romero–Rangel (2001), Velasco–de León y Romero–Rangel (2001) y Martínez–Cabrera et al. (2003). Los fósiles encontrados en la región de SMA corresponden principalmente a angiospermas, aunque están acompañadas por otros grupos de plantas (Equisetum; Silva–Pineda et al., 2009), restos de vertebrados (peces y anfibios, Fuentes–Guerra, 2005) y de invertebrados (Zaragoza–Caballero y Velasco–de León, 2003). Debido a que las hojas de encinos son las más numerosas, en este trabajo se pretende describir la diversidad de Quercus en la región de Santa María Amajac.
Registro fósil
El registro fósil de este género en el continente Americano abarca el Albiano de Kansas en donde reportan la presencia de las especies: Quercus wardiana (Ward, 1899), Q. stantoni (Berry, 1929) y Q. richardsoni (Bell, 1957). Los registros se incrementan a 21 en el Paleoceno de Estados Unidos; en Europa se menciona para el Mioceno la presencia de 10 especies, mientras que para Japón el registro es de siete especies del Mioceno–Plioceno. Bones (1979) y Manchester (1981) han realizado estudios en madera, hojas y frutos para el Eoceno Medio de la Formación Clarno en Oregón; Daghlian y Crepet (1983) afirman que la división del género en los dos subgéneros se llevó a acabo en el Oligoceno. Jones (1986) considera que los tipos de hojas actuales de la familia Fagaceae aparecieron en el hemisferio norte en diferente tiempo, presentándose primero las hojas enteras en arbustos del Cretácico Superior y Paleoceno, mientras que en el Eoceno se registran dientes redondeados y regulares; las formas lobadas aparecen en el Oligoceno y vienen a ser comunes en el Eoceno. Las hojas de margen dentado y craspedódromas son encontradas desde el Eoceno (Crepet y Nixon 1989).
En México, las localidades donde se presentan megafósiles de este género son escasas. Para el estado de Hidalgo, Geyne et al. (1963) reporta la presencia de Quercus sp. al sureste del pueblo de Tezuantla. Espinoza y Rzedowski (1966) estudiaron las impresiones colectadas en el cerro de la Estrella del Pleistoceno superior, donde identificaron las especies: Q. hahnii, Q. hartewegi, Q. laeta, Q. laurina, Q. repanda, Q. rugosa y Q. rugulosa. De igual forma, Lozano y Cevallos (2007) citan hojas fósiles de este género para el Plioceno del Estado de México y Pleistoceno de Puebla y Morelos. Millán y Velasco–de León (2009) registran para el Pleistoceno de Jalisco (La Primavera) Quercus eduardii, Q. obtusata, Q. desertícola, Q. castanea y Q. crassifolia; finalmente Hernández y Velasco–de León (2009) proponen la presencia en el Plioceno de Michoacán (Charo) de Q. crassipes y Q. hintonii.
MATERIAL Y MÉTODO
Los fósiles fueron colectados en secuencias sedimentarias de la región de SMA, localizadas en la porción Norte–Centro de la provincia morfotectónica del CVT, aproximadamente a 34 km de la ciudad de Pachuca. Los afloramientos estudiados pertenecen a la Formación Atotonilco El Grande (Segerstrom, 1961).
Las impresiones de hojas fueron colectadas en las localidades (Figura 1) de Los Baños, (20° 18' 18'' N, 98° 42' 44.4'' O) y Sanctorum (20°18'18.5'' N, 98°46'52.5''O) . En Los Baños, la secuencia sedimentaria mide 96.9 m de espesor y se caracteriza por la presencia de tres tipos de rocas: conglomerado en la parte superior y por debajo de éstos, estratos alternos y de espesor variable de arenisca y lutita. En la localidad de Sanctorum, la secuencia mide 70 m de espesor, y está formada por arenisca de grano fino, ceniza volcánica, estratos de yeso y lutita lodosa, estas últimas en laminaciones de milímetros. La evidencia fósil, así como las características geológicas de la región señalan la existencia de un ambiente lacustre con poca energía fluvial en la zona de SMA a finales del Neógeno (Salvador–Flores, 2001; Arellano et al., 2005) en donde las hojas fósiles fueron preservadas principalmente como impresiones y en menor cantidad como compresiones carbonosas.
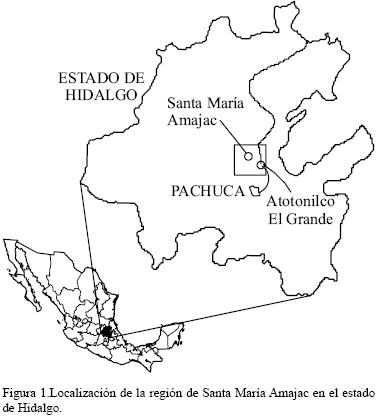
Las dataciones radiométricas (Kowallis et al., 1998) de depósitos lacustres realizadas en la región indican un máximo de edad del Plioceno Tardío.
La observación morfológica detallada de las hojas fósiles se realizó con un microscopio estereoscopio (Nikon, modelo SMZ800), los ejemplares fósiles fueron fotografiados con una cámara digital (Coolpix modelo E995), las imágenes obtenidas fueron procesadas con el programa Adobe Photoshop versión CS2. Para las mediciones se empleó el programa Image J (Abranoff et al., 2004). Los ejemplares se incorporaron a la Colección de la Facultad de Estudios Superiores Zaragoza, UNAM con los acrónimos CFZSMA y CFZST.
Análisis fenético
La selección de los caracteres (Tabla 1) para el análisis fenético se fundamentó en los ejemplares revisados en el Herbario Nacional del Instituto de Biología (MEXU), el Herbario de la Facultad de Estudios Superiores Iztacala, (IZTA) y el Herbario de la División de Ciencias Forestales de la Universidad Autónoma de Chapingo (UACH) (Anexo 1) y en discontinuidades observadas en las especies actuales y fósiles en el trabajo realizado por Velasco–de León y Romero–Rangel (2001), en el cual se validan los caracteres que son menos influenciados por factores ambientales, dejando fuera el tamaño de la lámina foliar, característica fuertemente relacionada con las condiciones ambientales (Velasco–de León, 1999). La terminología empleada es la utilizada por Hickey (1973) y el manual de arquitectura foliar del Leaf Architecture Working Group (1999).
Las hojas fósiles de SMA fueron comparadas con ejemplares de cinco especies fósiles del oeste de Estados Unidos (depositados en el Museo de Paleontología de la Universidad de California, en Berkeley) (Anexo 2), Quercus berryi Trelease, Q. consimilis Newberry, Q. dayana Knowlton, Q. oregoniana Knowlton y Q. simulata Knowlton (la elección de las especies fósiles se fundamentó en la presencia de la mayoría de los caracteres observados en los ejemplares fósiles de SMA) y con ejemplares de 27 especies actuales que se distribuyen a lo largo del CVT (Q. affinis Scheidweiler, Q. candicans Née, Q. castanea Née, Q.crassipes Humboldt et Bonpland, Q. crassifolia Humboldt et Bonpland, Q. deserticola Trelease, Q. dysophylla Bentham, Q. eduardii Trelease, Q. frutex Trelease, Q. glaucoides Martens et Galeotti, Q. greggii Trelease, Q. grisea Liebman, Q. laeta Liebman, Q. lancifolia Cham et Schltdl, Q. laurina Humboldt et Bonpland, Q. liebmani Oersted, Q. mexicana Humboldt et Bonpland, Q. microphylla Née, Q. obtusata Humboldt et Bonpland, Q. potosina Trelease, Q. repanda Humboldt et Bonpland, Q. rugosa Née, Q. salicifolia Neé, Q. sapotifolia Liebmann, Q. sartorii Liebmann, Q. sebifera Trelease, Q. viminea Trelease). Con la información obtenida se corrió un análisis fenético usando el programa NTSYS versión 2.1. Se consideraron 23 caracteres (cuantitativos y cualitativos multiestado) y 40 OTU (unidades taxonómicas operativas, Tabla 2); debido a la naturaleza de la información fue necesario estandarizar la matriz básica de datos con el fin de homogeneizar la escala de valores de las diferentes mediciones. La similitud de las OTU se estimó utilizando el coeficiente de similitud "Distancia Taxonómica Promedio", mientras que el agrupamiento de las OTU se realizó mediante el método de agrupamiento de pares no ponderados usando la media aritmética (Unweighted Pair Group Method Using Arithmetic Averages, UPGMA) graficando por último los resultados. Para determinar la robustez del método aplicado, y determinar por tanto la viablilidad de los caracteres utilizados, se calculó el índice cofenético de correlación (r).
RESULTADOS
A través del análisis de agrupamiento se delimitaron siete nuevas especies fósiles de la región de SMA. En el único fenograma obtenido (Figura 2) se puede distinguir la separación de las OTU en dos grupos (r =0.9045). En el primero (A) se observa el aislamiento de la especie fósil Q. berry. El grupo B, por otro lado, reúne a las 39 OTU restantes y se divide a su vez en dos subgrupos: el subgrupo B.1 que incluye a las especies actuales Q. mexicana y Q. crassifolia (encinos rojos), mientras que el subgrupo B.2 incluye al resto de las OTU incluidas en el análisis. En la parte baja de este subgrupo se puede observar a las especies fósiles de Estados Unidos (Q. simulata, Q. dayana, Q. consimilis) y al ejemplar CFZST 1617 (Q. hidalguensis n. sp.) que se separan de los ejemplares actuales y fósiles de México. El parecido fenético entre estas especies es escaso (1.4). En la parte media de este subgrupo se ubican los ejemplares fósiles CFZST 1864 (Q. cevallosi n. sp.) y CFZSMA 72 (Q. rugosus n. sp.). En el extremo superior del subgrupo B.2 se localiza el ejemplar CFZST 167 que corresponde a la especie actual Q. potosina y por último cuatro ejemplares fósiles de SMA, CFZSMA 43 (Q. ovatus n.sp.), CFZSMA 404 (Q. grandensis n. sp.), CFZSMA 156 (Q. curvensis n. sp.) y CFZSMA 317 (Q. amajensis n. sp.).
PALEONTOLOGÍA SISTEMÁTICA
Clase Magnoliopsida
Suclase Hamamelidae
Orden Fagales
Familia Fagaceae Durmotier
Subfamilia Quercoidae
Género Quercus Linnaeus, 1753
Quercus amajensis "especie nueva"
Diagnosis. Hoja obovada; ápice redondo; base aguda; margen entero en ocasiones con pequeños dientes sin aristas en el tercio superior de la lámina; venación semicraspedódroma; vena media ligeramente curva, masiva; ocho a nueve pares de venas secundarias agudas, con mayor espaciamiento en la parte central de la lámina; terciarias percurrentes alternas y opuestas con curso sinuoso; cuaternarias en retículo poligonal; areolas cuadrangulares pequeñas.
Diagnosis. Obovate leaf; obtuse apex, acute base, entire margin with small teeth on upper third section of the blade, without awn; semicraspedodromous venation; slightly curved mid–vein, massive; eight to nine pairs of acute secondary veins, with a greater distribution in the center of the blade; alternate and opposite percurrent tertiary veins with a sinuous path; quaternary veins in a polygonal reticulum; small square areole.
Descripción. Hojas obovadas; ápice redondo; base aguda; largo 3.2 cm, ancho 2.2 cm, relación largo:ancho de 1.5:1; margen entero a dentado, dientes muy pequeños sin aristas en el tercio superior de la lámina; venación pinada semicraspedódroma; vena media ligeramente curvada; ocho a nueve pares de venas secundarias agudas que se insertan a la vena media en ángulos (25–50°), rectas a ligeramente curvadas, alternas, con espaciamiento mayor en la parte central de la lámina; sin intersecundarias, venas terciarias percurrentes alternas y opuestas; cuaternarias formando areolas cuadrangulares pequeñas.
Etimología. El epíteto específico se refiere a la región donde aflora la localidad fosilífera de donde proviene el material.
Material examinado. Tres impresiones, holotipo: CFZSMA317.
Ocurrencia. Los Baños, Formación Atotonilco El Grande. Plioceno.
Discusión. El fenograma muestra poca similitud con el agrupamiento de Q. curvensis y Q. grandensis, ya que se presentan diferencias en el patrón de venación. Q. amajensis presenta venación semicraspedódroma, margen entero en ocasiones con dientes muy pequeños, mientras que Q. curvensis posee margen dentado con aristas cortas, además posee venas intersecundarias fuertes simples y venación hasta de quinto orden, en Q. amajensis llega hasta cuarto orden; Q. grandensis con margen dentado, vena media recta y venas intersecundarias compuestas.
Quercus cevallosi "especie nueva"
Diagnosis. Hoja elíptica; ápice incompleto; base incompleta; margen dentado, con dientes agudos aristados, seno redondeado, la vena secundaria entra al diente en un recorrido central; venación craspedódroma; secundarias agudas (38–42°); terciarias percurrentes alternas y opuesta, con un ángulo recto perpendicular a la vena primaria; cuaternarias reticuladas, poligonales regulares; quinquenarias; areolas cuadrangulares pequeñas, con venillas sin ramificar.
Diagnosis. Elliptic shaped leaf; incomplete apex; incomplete base; dentate margin, with acute aristate teeth, rounded sinus, the secondary vein enters the teeth through the middle; craspedodromous venation; acute secondary veins (38–42°); alternate and opposite percurrent tertiary veins, with a straight angle perpendicular to the primary vein, quaternary reticulate, regular polygonal shape; quinary venation; small square areole, with veins without branching.
Descripción. Hoja elíptica; ápice incompleto; base incompleta; largo 4.5 cm, ancho 3.0 cm; relación largo ancho 1.2:1; margen serrado, dientes agudos aristados, seno redondeado y profundo (Figura 3c); venación craspedódroma; vena media recta; más de cuatro pares de venas secundarias ligeramente curvas que se insertan en ángulo agudo (38–42°) alternas, con espaciamiento mayor hacia la base; venas terciarias percurrentes opuestas y alternas; venas cuaternarias reticuladas; Las quinquenarias forman areolas cuadrangulares pequeñas (0.08, 0.07 mm) con venillas sin ramificar. (Figura 3d).
Etimología. El epíteto específico se propone como un reconocimiento al Dr. Sergio Cevallos Ferriz por su extenso trabajo en floras de angiospermas cretácicas.
Material examinado. Impresión, holotipo CFZSt 1864.
Ocurrencia. Sanctorum, Formación Atotonilco El Grande. Plioceno.
Discusión. Se une con Q. oregoniana, con muy poca similitud, esta especie puede presentar venación craspedódroma como Q. cevallosii y también semicraspedódroma. Los dientes en Q. cevallosi siempre son agudos, muy espaciados y en Q. oregoniana pueden ser agudos y redondos, están muy juntos, son numerosos (de 15 a 20) y no presentan aristas.
Quercus curvensis "especie nueva"
Diagnosis. Hojas elípticas; ápice agudo, base redonda que tiende a aguda; margen dentado hasta la mitad de la lámina, con aristas cortas; la vena secundaria entra al diente en un recorrido exmedial; venación semicraspedódroma; vena media recta; secundarias agudas; intersecundarias fuertes simples; terciarias percurrentes alternas; cuaternarias reticuladas poligonales regulares; quinquenarias forman areolas cuadrangulares pequeñas.
Diagnosis. Elliptic shape leaf; acute apex; rounded base tends to acute; a dentate margin to the middle of the blade, with short awn; the secondary vein enters the tooth with an exmedial orientation; semicraspedodromous venation, straight mid–vein; acute secondary veins; simple strong intersecondaries; alternate percurrent tertiary veins; regular polygonal reticulate quaternary veins; quinary veins form small square areole.
Descripción. Hojas elípticas; ápice agudo; base redonda que tiende a aguda; largo 6.5 cm, ancho 2.9 cm; relación largo ancho de 2:1; margen serrado, dientes cercanos al ápice con aristas cortas de 0.38 mm de largo; venación pinada semicraspedódroma, arcos agudos; venas secundarias agudas (20–40°) disminuyendo hacia la base; opuestas y alternas, el espaciamiento decrece hacia la base (Figura 3e); la vena entra al diente con un recorrido exmedial; venas intersecundarias; terciarias percurrentes alternas, transversales; cuaternarias; las quinquenarias forman areolas pequeñas y cuadrangulares.
Etimología. El epíteto específico hace referencia al nombre común con el que se conoce esta localidad.
Material examinado. Dos impresiones, holotipo CFZSMA156.
Ocurrencia. Los Baños, Formación Atotonilco El Grande. Plioceno.
Discusión. En el fenograma se une con Q. grandensis, que presenta margen dentado venas terciarias reticuladas mientras que en Q. curvensis son percurrentes alternas y los dientes tienen aristas a diferencia de Q. grandensis donde carecen de estas estructuras.
Quercus grandensis "especie nueva"
Diagnosis. Hojas obovadas; ápice incompleto; base redonda; margen dentado; la vena secundaria entra al diente en un recorrido exmedial; venación semicraspedódroma; dientes redondeados presentes hasta la mitad de la lámina; vena media recta, fuerte; pocas venas secundarias, agudas; intersecundarias compuestas; terciarias reticuladas poligonales regulares; cuaternarias reticuladas poligonales regulares; areolas cuadrangulares.
Diagnosis. Obovate shaped leafs, incomplete apex; rounded base; dentate margin; the secondary vein enters the tooth with an exmedial orientation; semicraspedodromous venation; presence of rounded teeth to the middle of the blade; strong straight mid–vein; few acute secondary veins; compound intersecondary veins, regular reticulate polygonal tertiary veins, regular reticular polygonal quaternary veins; square areole.
Descripción. Hojas obovadas a elípticas; ápice incompleto, puede ser agudo; base redonda; largo 4.73 cm; ancho 3.89 cm; relación largo–ancho de 1.4:1; margen dentado, dientes redondeados, presentes hasta la mitad de la lámina no se puede observa la presencia de arista o mucrón, seno redondeado muy amplio; vena media recta, masiva; venación semicraspedódroma; venas secundarias agudas (25–42°) el ángulo es mayor en la base; el espaciamiento decrece en la base; venas terciarias reticuladas; areolas cuadrangulares pequeñas (0.6).
Etimología. El epíteto específico hace referencia a la formación donde fue colectado el material.
Material examinado. Tres impresiones, holotipo CFZMT404.
Ocurrencia. Los Baños, Formación Atotonilco El Grande. Plioceno.
Discusión. En el fenograma se une con Q. curvensis, que presenta margen dentado hasta la mitad de la lámina; venas terciarias percurrentes alternas mientras que en Q. grandensis son reticuladas, los dientes son redondeados a diferencia de Q. curvensis donde los dientes presentan aristas cortas.
Quercus hidalgensis "especie nueva"
Diagnosis. Hoja elíptica; ápice redondo; base redonda; dentada, con dientes redondos en el tercio apical de la lámina, seno agudo; vena media débil y se curva en el último tercio; venación craspedódroma; venas secundarias agudas en el ápice y en la base rectas, espaciamiento irregular; venas intersecundarias simples y compuestas; terciarias reticuladas; areolas cuadrangulares y pentagonales
Diagnosis. Elliptic leaf; rounded apex; rounded base; dentate, with rounded teeth presents in the apical portion of the blade, acute sinus; weak mid–vein which curves in last third of the leaf; craspedodromous venation; acute secondary veins located at the apex and straight at the base, irregular scattering, simple and compound intersecondary veins; reticulate tertiary veins; square and pentagonal areole.
Descripción. Hoja elíptica; ápice redondo; base redonda; largo 3.2 cm; ancho 2.5 cm; relación largo ancho de 1.28:1; margen dentado (probablemente mucronado); vena media débil y se curva en el último tercio de su longitud; venación craspedodróma; venas secundarias rectas; agudas en el ápice (15–25°) en la base casi rectas 85°; 10 pares de venas intersecundarias; espaciamiento irregular; terciarias reticuladas; areolas cuadrangulares y pentagonales.
Etimología. El epíteto específico hace referencia al Estado de Hidalgo, donde aflora la Formación Atotonilco El Grande.
Ejemplares examinados. Impresión: holotipo CFZST1617.
Ocurrencia. Sanctorum, Formación Atotonilco El Grande. Plioceno.
Discusión. Se une a Q. consimilis con muy poca similitud, Q. hidalgensis presenta venación craspedodroma y Q. consimilis puede presentar venación broquidódroma–craspedódroma, las venas terciarias en Q. hidalgensis son reticuladas y en Q. consimlis se presenta mayor variedad pues pueden ser percurrentes alternas, opuestas y transversales.
Quercus ovatus "especie nueva"
Diagnosis. Hoja elíptica; ápice incompleto; base incompleta; margen entero; vena media recta robusta; broquidódroma; venas secundarias agudas con un ángulo más pequeño hacia el ápice que se incrementa hacia la base, espaciamiento irregular; terciarias mixtas opuestas y transversales; cuaternarias; las quinquenarias forman areolas medianas.
Diagnosis. Elliptic leaf; incomplete apex; incomplete base, entire margin; straight, robust mid–vein; brochidodromous; acute secondary veins with a small angle at the apex which increases towards the base, irregular scattering; opposite and transversal mixed tertiary veins; quaternary veins; quinary veins form medium areole.
Descripción. Hoja elíptica; ápice incompleto; base aguda; largo 5.5 cm; ancho 3.2 cm; relación largo–ancho 1.71:1; margen entero; vena media robusta (0.8 mm); venación pinada, broquidódroma; venas secundarias agudas (20–67°), espaciamiento irregular, mayor en la base; terciarias transversales; cuaternarias; las quinquenarias forman areolas cuadrangulares, medianas.
Etimología. Su epíteto específico hace alusión a la forma de la lámina.
Material examinado. Cuatro impresiones, Holotipo CFZSMA43.
Ocurrencia: Los Baños, Formación Atotonilco El Grande. Plioceno.
Discusión. Se une al agrupamiento formado por Q. grandensis, Q. curvensis, Q. amajensis y difiere de ellos en la venación secundaria que es broquidódroma, no posee dientes y su vena media es robusta.
Quercus rugosus "especie nueva"
Diagnosis. Hojas elípticas; ápice redondo; base redonda; margen entero; venación broquidódroma; vena media recta moderada; venas secundarias agudas, que se curvan en la última parte de su recorrido; venas secundarias muy gruesas; intersecundarias simples en la base; terciarias percurrentes alternas; cuaternarias; quinquenarias forman areolas cuadrangulares pequeñas.
Diagnosis. Elliptic leaves; obtuse apex; rounded base; entire margin; brochidodromous venation; moderate straight mid–vein; acute secondary veins which curve in the last part of their course; very thick secondary veins; simple inter–secondary veins at the base; alternate and thick percurrent tertiary veins; quaternary veins; quinary veins form small square areole.
Descripción. Hojas elípticas; ápice redondo; base redonda; largo 5.1 cm; ancho 2.4 cm; relación largo ancho de 2:1; margen entero; 7–10 pares de venas secundarias muy fuertes, que se insertan a la vena media en ángulo agudo (25–60°), últimos pares rectos, su espaciamiento decrece en la base; intersecundarias simples en la base; terciarias percurrentes, alternas; cuaternarias formando retículo poligonal; quinquenarias forman areolas cuadrangulares pequeñas.
Etimología. Su epíteto específico hace alusión a la venación que da un aspecto rugoso.
Material examinado. Tres impresiones, Holotipo CFZSMA72.
Ocurrencia. Los Baños, Formación Atotonilco El Grande. Plioceno.
Discusión. Se une con Q. eduardii con poca similitud. La venación de Q. rugosus es broquidódroma, puede presentar hasta 10 pares de venas secundarias, la venación de Q. eduardii semicraspedódroma, con siete pares de venas secundarias y dientes agudoscon pequeñas aristas (Figuras 4a y 4b) presentan venación de sexto orden y areolas cuadrangulares pequeñas (Figura 4c).
DISCUSIÓN
Las hojas fósiles descritas tienen caracteres morfológicos que permiten su identificación como miembros del género Quercus. Entre estos caracteres se incluyen hojas generalmente pecioladas, de láminas simples, enteras o divididas en el margen. La venación primaria es pinnada, la secundaria va de semicaspedódromas, a craspedódromas con dientes mucronados y/o aristados, las venas secundarias entran al diente de forma central o exmedial, en las hojas enteras se presenta venación broquidódroma. Con venas intersecundarías simples y compuestas, que divergen de la vena media en ángulos iguales o mayores que el de las venas secundarias. La venación de alto orden esta bien desarrollada (cinco niveles son comunes) al igual que las areolas, estas ultimas por lo regular son cuadrangulares, medianas y pequeñas, con venillas ramificadas.
Nixon (1993) distingue para este taxón dos subgéneros: Quercus y Cyclobalanopsis. Al subgénero Quercus lo divide en tres secciones con presencia en México, Lobatae, Protobalanus y Quercus. Las especies del género con presencia en el CVT y que se integraron al análisis pertenecen únicamente a las secciones Lobatae y Quercus (Valencia, 2007).
El género Quercus ha sido revisado sistemáticamente varias veces durante los últimos años (Martínez, 1952, 1959; Mc Vaugh, 1974; Valencia, 2004) mostrando gran variabilidad morfológica, tanto a nivel interespecífico como intraespecífico, por lo que es un grupo difícil taxonómicamente. Para México no se conoce con exactitud el número de especies. Por ejemplo Trelease (1924), calcula alrededor de 253 especies y Camus (1934–1954) reconoce 238 especies. Años después, Nixon (1993) menciona entre 135 y 150 especies. Desafortunadamente algunos de los caracteres importantes utilizados para describir a las especies actuales no se observan en el material fósil (color de la hoja) o bien pueden estar influenciados por el ambiente (tamaño de la hoja, presencia y abundancia de tricomas), por lo que la identificación del material fósil de SMA está basada en los caracteres de la arquitectura foliar de las hojas observados en los ejemplares de herbario revisados y discontinuidades registradas en las especies (Velasco–de León y Romero–Rangel, 2001). La elección de las especies fósiles se fundamentó en la presencia de la mayoría de los caracteres observados en los ejemplares fósiles de SMA.
De los 22 caracteres cualitativos y cuantitativos empleados para este estudio, haremos mención de aquellos que permitieron el agrupamiento de las OTU: El patrón de venación en las venas secundarias que se presentó fue craspedódromo en Q. cevalli y Q. hidalgensi. semicraspedódromo en Q. amajensi. Q. curvensis y Q. grandensis, mientras que Q. ovatus y Q. rugosus presentan un patrón broquidódromo. Este último carácter está muy relacionado con el tipo de margen, pues las cinco primeras especies tienen margen dentado, mientras que la venación broquidódroma se exhibe en aquellas hojas que muestran margen entero. En el caso de Q. cevalli, los dientes son agudos y con un sinuos redondeado, característica que lo diferencia de Q. hidalgensis, en la cual los dientes son redondos y de sinuos agudo. En la venación semicraspedódroma los dientes son muy pequeños como en Q. amajensis y además con pequeñas aristas en Q. curvensis. En algunas especies actuales, el recorrido de la vena secundaria al entrar al diente ha sido considerado un carácter taxonómico importante (Martínez–Cabrera, 2002; Martínez–Cabrera et al., 2003); en este estudio, en los ejemplares fósiles de Q. cevalli dicho recorrido es central, mientras que en Q. curvensis y Q. grandensis es exmedial. En el fenograma, este carácter (recorrido central) así como el tamaño de las areolas es compartido por Q. candicans (Figuras 4d, 4e y 4f) y Q. potosina (Figuras 4g, 4h, 4i) que forman un subgrupo, junto con el ejemplar CFZ167, el cual presenta recorrido exmedial y diferente tamaño de las areolas. Otros caracteres que se consideran importantes en la separación de especies son el ángulo de divergencia y número de las venas secundarias. Este es el primer reporte de hojas fósiles de encinos con una edad mayor al Pleistoceno para México (Nixon, 1993) por lo que la propuesta de estas nuevas especies puede ayudar a entender la alta diversidad de especies (36) de este género en el CVT. Las condiciones fisiográficas, los cambios climáticos que se han presentado desde el Plioceno y que han provocado la fragmentación y contacto de biotas, la aparición de nuevos hábitats y la alta variabilidad del género, pueden explicar no sólo la diversidad del grupo en las localidades de Los Baños y Sanctorum en el Plioceno, sino también la diversidad actual del grupo (Valencia, 2007).
CONCLUSIÓN
Los caracteres analizados en este trabajo permiten concluir que a partir de la arquitectura foliar es posible diferenciar las especies actuales y fósiles de Quercus y que ya para el Plioceno de México se presentaba una alta diversidad en este género, en las cercanías de un lago en las localidades de Santa María Amajac y Sanctorum en el Estado de Hidalgo.
AGRADECIMIENTOS
Los autores desean agradecer a la Dirección General de Asuntos del Personal Académico por su apoyo al proyecto PAPIIT IN108706 "Paleoecología en dos sectores del Eje Neovolcánico".
REFERENCIAS
Abranoff, M.D., Magelhaes, P. J., Ram, S.J., 2004, Image processing with Image: Journal of Biophotonics International, 11(7), 36–42. [ Links ]
Arellano–Gil, J., Velasco de León, M.P., Silva Pineda, A., Salvador–Flores, R., Beltrán–Romero, F., 2005, Origen y características geológicas del paleo–Lago de Amajac, Hidalgo: Revista Mexicana de Ciencias Geológicas, 22(2), 199–211. [ Links ]
Bell. W.A., 1957, Flora of the Upper Cretaceous Nanaimo Group of Vancouver Island, British Columbia: Geological Survey of Canada Memoir, 293, 1–84. [ Links ]
Berry. E.W., 1929, The flora of the Frontier Formation: United States Geological Survey Professional Paper, 158,129–135. [ Links ]
Bones, T. J., 1979, Atlas of fossil fruits and seeds from north central Oregon: Oregon Museum of Science and Industry, Occasional Papers in Natural Science, 1, 1–23. [ Links ]
Camus, A., 1934–1954, Les chénes: monographie du genre Quercus (et Lithocarpus): Paris, P. Le Chevalier & Fils, Encyclopédie économique de sylviculture, v. 6–8. [ Links ]
Crepet, W.L., Nixon., K.C., 1989, Earliest megafossil evidence of fagaceae: phylogenetic and biogeographic implications: American Journal of Botany, 76(6), 842–855. [ Links ]
Daghlian, C.P., Crepet, W.L., 1983, Oak catkins, leaves and fruits from the Oligocene Catahoula Formation and their evolutionary significance: American Journal of Botany, 70(5), 639–649. [ Links ]
Espinosa, D., Llorente, J., Morrone, J.J., 2006, Historical biogeographical patterns of the species of Bursera (Burseraceae) and their taxonomic implication: Journal of biogeography, 33(11), 1945–1958. [ Links ]
Espinoza, R.J., Rzedowski, J., 1966, Flórula del Pleistoceno Superior del Cerro de la Estrella, próximo a Ixtapalapa D.F. (México): Anales de la Escuela Nacional de Ciencias Biológicas, 16, 9–21. [ Links ]
Ferrusquía–Villafranca, I., 1993, Geology of Mexico: A synopsis, en Ramammorthy, T. P., Bye, R., Lot, A., Fa, J. (eds.), Biological diversity of Mexico: Origins and distribution: Oxford, Oxford University Press, 3–107. [ Links ]
Fuentes–Guerra, E.P., 2005, Estudio de los anfibios pertenecientes al Plioceno tardío en la localidad de Sanctorum en el estado de Hidalgo. Orden Anura: México, D.F., Universidad Nacional Autónoma de México, Facultad de Estudios Superiores Zaragoza, Tesis profesional, 84 pp. [ Links ]
Geyne, A.R., Fries, C., Segerstrom, K., Black, R.F., Wilson, I.F., 1963, Geología y Yacimientos Minerales del distrito de Pachuca–Real del Monte: Consejo de Recursos Naturales no Renovables, Publicación 5E, 30–35. [ Links ]
Hernández, G.O., Velasco de León, M.P., 2009, Estudio paleobotánico del género Quercus en el estado de Michoacán (resumen), en XI Congreso Nacional de Paleontología, Juriquilla, Querétaro, p. 108. [ Links ]
Hickey, L.J., 1973, Classification of the architecture of dicotyledonous leaves: American Journal of Botany, 60(1), 17–33. [ Links ]
Jones, J.H., 1986, Evolution of the Fagaceae: The implications of foliar features: Annals of the Missouri Botanical Garden, 73, 228–275. [ Links ]
Kowallis, B.J., Swisher, C.C., Carranza–Castañeda, O., Miller, W.E., Tingey, D.G., 1998, Fission–track and single–crystal 40 Ar/Ar 39 ions fusion ages from volcanic ash layer in fossil–bearing Pliocene sediments in Central–Mexico: Revista Mexicana de Ciencias Geológicas, 15(2), 157–160. [ Links ]
Leaf Architecture Working Group., 1999, Manual of leaf architecture, morphological description and categorization of dicotyledonous and net–veined monocotyledonous angiosperms: Washington, Smithsonian Institution, 65 pp. [ Links ]
Linnaeus, C., 1753, Species plantarum: Stockholm, Laurentii Salvii, 559 pp. [ Links ]
Lozano, S.M., Cevallos, F.S., 2007, Historia de la Vegetación del centro de México: Evidencias paleobotanicas, en Luna, I., Morrone, J.J., Espinosa, D. (eds.), Biodiversidad de la Faja Volcánica Transmexicana: México, Universidad Nacional Autónoma de México, 273–287. [ Links ]
Manchester, S.R., 1981, Fossil plants of the Eocene Clarno nut beds: Oregon Geology, 43, 75–81. [ Links ]
Martínez, M., 1952, Los encinos de México y Centroamérica, II: Anales del Instituto de Biología, 23, 53–83. [ Links ]
Martínez, M., 1959, Los encinos de México, XII: Anales del Instituto de Biología, 30, 119–140. [ Links ]
Martínez–Cabrera, D., 2002, Variación morfométrica de dos especies de encinos rojos Q. sartorii Liebmann y Q. xalapensis Humboldt y Bondpland (Fagaceae): Montecillo, Texcoco, México, Colegio de Postgraduados, Tesis de Maestría, 86 pp. [ Links ]
Martínez–Cabrera, D, Terrazas, T., Zavala–Chávez, F., 2003. Arquitectura foliar y anatomía de la corteza y madera de Q. sartorii y Q. xalapensis (Fagaceae): Boletín de la Sociedad Botánica de México, 73, 63–72. [ Links ]
Mc Vaugh, R., 1974, Flora Novo–Galiciana (Fagaceae): Contributions from the University of Michigan Herbarium, v.12, parte 1, núm.3, 93 pp. [ Links ]
Millán, E.D.M., Velasco–de León, M.P., 2009, Estudio paleobotánico del género Quercus en el estado de Jalisco empleando arquitectura foliar (resumen), en XI Congreso Nacional de Paleontología, Juriquilla, Querétaro, p. 100. [ Links ]
Nixon, K.C., 1993, The genus Quercus in México, en Ramammorthy, T.P., Bye, R., Lot, A., Fa, J. (eds.), Biological Diversity of México, origins and distribution: New York, Oxford University Press, 447–458. [ Links ]
Romero–Rangel, S., 2001, Estudio taxonómico de la serie acutifoliae (Quercus, Fagaceae): México, D.F., Universidad Nacional Autónoma de México, Tesis Doctoral, 174 pp. [ Links ]
Salvador–Flores, R., 2001, Origen sedimentológico y estratigrafía del Paleolago de Amajac, Hidalgo: México, D.F., Universidad Nacional Autónoma de México, Tesis de Licenciatura, 87 pp. [ Links ]
Segerstrom, K., 1961, Geología del sureste del Estado de Hidalgo y del noreste del Estado de México: Boletín de la Asociación Mexicana de Geología del Petróleo, 34,147–168. [ Links ]
Silva–Pineda, A., Velasco–de León, M.P., Aguilar, F.J., Chacon, E., 2009, An Upper Pliocene Equisetum (Equisetales) from Atotonilco el Grande Formation in Central México: Paleontological Journal, 43(2), 216–225. [ Links ]
Trelease, W., 1924, The American Oaks: Memoirs of the National Academy of Sciences, 20, 1–255. [ Links ]
Valencia, A.S., 1994, Contribución a la delimitación taxonómica de tres especies del género Quercus subgénero Erythrobalanus: Q. laurina Humboldt et Bonpland, Q. affinis Scheidweller y Q. Ghiesbregtii Martens et Galeotti: México, Universidad Nacional Autónoma de México, Tesis de Maestría, 174 pp. [ Links ]
Valencia, S., 2004, Diversidad del Género Quercus (Fagaceae) en México: Boletín de la Sociedad Botánica de México, 75, 33–53. [ Links ]
Valencia, A.S., 2007, Encinos, en Luna, I., Morrone, J.J., Espinosa–Organista, D. (eds.), Biodiversidad de la Faja Volcánica Transmexicana: México, Universidad Nacional Autónoma de México, Comisión Nacional para el Conocimiento y Uso de la Biodiversidad,139–148. [ Links ]
Velasco–de León, M.P., 1999, Estudio paleoecológico de una comunidad del Terciario en el estado de Puebla: México, Universidad Nacional Autónoma de México, Facultad de Ciencias, Tesis doctoral, 120 pp. [ Links ]
Velasco–de León, M.P., Romero–Rangel, S., 2001, Arquitectura Foliar de diez especies de Quercus (Fagaceae) como fuente de información para la identificación de ejemplares fósiles (resumen), en XV Congreso Mexicano de Botánica, Querétaro, México, p.142. [ Links ]
Ward, L.F., 1899, The Cretaceous formation of the Black Hills as indicated by the fossil plants: Cretaceous Formation of the Black Hills: Proceedings of the United States National Museum, 21, 195–229. [ Links ]
Zaragoza–Caballero, S., Velasco–de León, M.P., 2003, Primer registro fósil de Epicauta (Coleoptera Meloidae) de la Formación Atotonilco El grande (Plioceno) del estado de Hidalgo, México: Revista Mexicana de Ciencias Geológicas, 20(2), 154–159. [ Links ]

