










Serviços Personalizados
Journal

Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
 Citado por SciELO
Citado por SciELO Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkRevista mexicana de ciencias geológicas
versão On-line ISSN 2007-2902versão impressa ISSN 1026-8774
Rev. mex. cienc. geol vol.27 no.1 Ciudad de México Abr. 2010
Dientes de terópodos del Cretácico Superior del Estado de Coahuila, México
Theropod teeth from the Upper Cretaceous of the State of Coahuila, Mexico
Esperanza Torres–Rodríguez1,*, Marisol Montellano–Ballesteros2, René Hernández–Rivera2 y Mouloud Benammi3
1 Posgrado en Ciencias Biológicas, Universidad Nacional Autónoma de México, 04510 México, D.F., México. *E–mail: alarbol@gmail.com.
2 Departamento de Paleontología, Instituto de Geología, Universidad Nacional Autónoma de México, Cd. Universitaria, Del. Coyoacán, 04510 México, D. F., México.
3 Institut International de Paléoprimatologie, Paléontologie Humaine: Evolution et Paléoenvironnements (IPHEP)–UMR CNRS 6046,SFA–Université de Poitiers, Bât. de Sciences Naturelles (3ème étage), 40 avenue du Recteur Pineau, F86022 Poitiers Cedex, Francia.
Manuscrito recibido: Septiembre 10, 2009.
Manuscrito corregido recibido: Diciembre 8, 2009.
Manuscrito aceptado: Diciembre 12, 2009.
RESUMEN
En México, los terópodos son un grupo de dinosaurios poco conocido, el escaso material descrito ha sido referido solamente a nivel de "familia". Como resultado de la investigación paleontológica realizada en el Estado de Coahuila, se recuperó material dental de terópodos. Se identificaron los géneros Saurornitholestes, cf. Troodon, así como Tyrannosauridae y un terópodo indeterminado. La composición faunística de terópodos es similar a la descrita en el Parque Nacional del Big Bend, Texas, confirmando la existencia de la zona biogeográfica de dinosaurios del sur de las Grandes Planicies de América del Norte. De confirmarse la presencia de Troodon en estas latitudes sugeriría que la distribución geográfica de este género de terópodo era mucho más amplia de lo supuesto anteriormente.
Palabras claves: paleobiogeografía, dinosaurios, dientes, Theropoda, Formación Aguja, Formación Olmos, Cretácico Tardío, México.
ABSTRACT
In Mexico, the theropods represents a poorly known group of dinosaurs, the scarce material had been referred only to "family" level. As a result of paleontological research carried on in the State of Coahuila, dental remains belonging to theropods were recovered. The identified genera are Saurornitholestes , cf. Troodon , as well as Tyrannosauridae and an indeterminate theropod. The theropod diversity of Coahuila is similar to that described for the Big Bend National Park, Texas, confirming the existence of a distinct dinosaurian biogeographic province in the southern Great Plains of North America. If the presence of Troodon at this latitude is confirmed, it would suggest that the geographic distribution of this theropod genus was much wider than previously supposed.
Key words: paleobiogeography, dinosaurs, teeth, Theropoda, Late Cretaceous, Aguja Formation, Olmos Formation, Mexico.
INTRODUCCIÓN
En el norte de México, en la frontera de los estados de Coahuila y Chihuahua con Texas, afloran rocas continentales de edad cretácica tardía. Cabrera et al. (1984) y personal del Instituto Nacional de Estadística Geografía e Informática (com. pers., 2000) mencionaron la presencia de restos de dinosaurios en las áreas conocidas como Altares y La Americana. En los últimos años, personal del Instituto de Geología de la Universidad Nacional Autónoma de México ha desarrollado el proyecto de investigación paleontológica "Cretácico Tardío continental del norte de los estados de Coahuila y Chihuahua" con el propósito de conocer la paleodiversidad de microvertebrados que habitaron la zona y evaluar la hipótesis del gradiente faunístico durante el Cretácico Tardío en América del Norte propuesta por Lehman (1987, 1997, 2001).
Como resultado de este proyecto se localizaron 17 sitios fosilíferos, y se han recuperado restos del ceratópsido Agujaceratops mariscalensis Lehman, 1989 (Andrade–Ramos et al., 2002) y de titanosaurio (Montellano–Ballesteros, 2003), entre otros grupos de dinosaurios, además de restos de tortugas y cocodrilos, entre otros (Montellano–Ballesteros et al., 2000). Entre el material fósil recuperado existen dientes aislados de terópodos que son el objeto de estudio de este trabajo.
Los terópodos conforman un grupo de dinosaurios que fue sumamente diverso durante el Mesozoico y abundante en el Cretácico, representan el 50% de las familias reconocidas de dinosaurios y el 40 % de los géneros válidos (Currie, 1988; Fastovsky et al., 2004). Los restos de terópodos han sido encontrados en todos los continentes incluyendo la Antártica (Fastovsky et al., 2005). Como la mayor cantidad de material de terópodos encontrada la constituyen dientes aislados, diversos autores como Currie et al. (1990), Smith (2005, 2007) y Smith et al. (2005) han propuesto el uso de ciertas características morfológicas de los dientes consideradas como diagnósticas para la identificación de terópodos a nivel de "familia", género e incluso de especie.
De las faunas cretácicas con terópodos registradas para América del Norte, Baszio (1997a), Sankey (2001) y Sankey et al. (2002 y 2005) observaron diferencias en la composición entre las asociaciones de dinosaurios terópodos del norte y las del sur. Por la posición geográfica de las localidades de donde proviene el material de estudio se supondría que la fauna recuperada debe ser similar a la reportada para el Parque Nacional del Big Bend, Texas. Sankey (2001) y Sankey et al. (2005) registraron la presencia de terópodos pertenecientes a Tyrannosauridae, así como los géneros Saurornitholestes , Richardoestesia y cf. Paronychodon recuperados en las formaciones Aguja y Tornillo, en Texas. Cabe mencionar que Sankey (2001) y Sankey et al. (2002, 2005) consideran a Paronychodon como género válido a pesar de que varios autores (ver Currie et al., 1990; Makovicky y Norell, 2004) lo consideran como una patología de dientes de terópodos pequeños.
ÁREA DE ESTUDIO Y MARCO GEOLÓGICO
El material de terópodos aquí mencionado fue recolectado en dos zonas del estado de Coahuila (Figura 1): Ejido "El Mezquite" en la Cuenca de Sabinas y el Rancho "Las Garzas" en el Municipio de Ocampo. El Ejido "El Mezquite" se encuentra dentro de la Cuenca de Sabinas que se ubica dentro de la conocida Región Carbonífera de Sabinas. Las rocas que afloran se han identificado como pertenecientes a la Formación Olmos o también llamada "Series de Carbón" (Dumble, 1982) y se consideran de edad maastrichtiana tardía por la presencia del bivalvo Exogyra costata Say, 1820 y del cefalópodo Sphenodiscus sp. (Stephenson, 1941). La Formación Olmos está constituida por lutita de color gris verdoso y capas de limolita finamente estratificadas; además hay presentes capas de arenisca de grano grueso a fino con estratificación cruzada y hacia la base contiene mantos de carbón. El material proviene de la capa que subyace al primer estrato de la Formación Escondido, la cual sobreyace a la Formación Olmos, y fue donado por el Ingeniero Juan Pablo García, miembro de la Asociación de Paleontólogos Aficionados de Sabinas (PASAC).

El material que proviene del Rancho "Las Garzas" fue recolectado a lo largo de varias temporadas de campo, en dos secciones llamadas Las Garzas y Anizul, desde el año 2000 hasta 2004; esta área se encuentra en la esquina más noroccidental del Municipio de Ocampo y ahí aflora una sucesión de lodolitas y depósitos de conglomerados; se han identificado seis niveles de donde proviene el material fósil (Figura 2). De acuerdo a Benammi y Montellano–Ballesteros (2006) las rocas que ahí afloran se correlacionan con los miembros "Middle Sandstone" y "Upper Shale" de la Formación Aguja (Campaniano–Maastrichtiano). El análisis paleomagnético de las rocas muestra una polaridad normal, que puede ser correlacionado con el chron C32n.2n que corresponde al Campaniano superior (entre 71.5 y 72.9 Ma.). Recientemente, en la base de la sección de "Las Garzas" se recolectó un ejemplar de un molusco gasterópodo identificado como Viviparus cf. V. australis Kues, 1995 (en Lucas et al., 1995; Dra. Carmen Perrilliat, com. pers.), el cual tiene un alcance desde el Santoniano hasta el Maastrichtiano, por lo que se considera que, con la información que se tiene hasta la fecha, toda esta sección comprende desde el Campaniano tardío al Maastrichtiano.
Las coordenadas geográficas de las localidades se encuentran en el catálogo de referencia geográfico del Museo de Paleontología del Instituto de Geología.
MATERIAL Y MÉTODO
El material fósil del Ejido "El Mezquite" fue recolectado en superficie, mientras que el material perteneciente al Rancho "Las Garzas" fue obtenido mediante la técnica del tamizado. Se utilizaron tamices de los números de malla 10 (apertura 0.64 mm) y 18 (apertura 0.0394 mm). Para la identificación del material se siguió la metodología propuesta por Smith (2005, 2007) y Smith et al. (2005). A cada diente de terópodo se le tomaron las siguientes medidas en vista lateral (de acuerdo con la Figura 3): altura de la corona (desde la punta hasta la base del diente), longitud de la base de la corona (medida a lo largo del segmento linear que va entre los puntos AB en la base del diente); anchura de la base de la corona; longitud apical; medición del segmento linear entre los puntos AG (medida para los dientes completos); densidad denticular basal de las carenas mesial y distal; densidad denticular media de las carenas mesial y distal; densidad denticular apical en las carenas mesial y distal. En el caso de los dientes de terópodos pequeños se determinó el número de dentículos cada milímetro, mientras que para los dientes de terópodos grandes se determinó cada 5 milímetros; el conteo se realizó tan cerca como fue posible de la partes basal, media y apical de la corona para obtener las densidades denticulares basales (MB y DB), medias (MC y DC) y apicales (MA y DA) de la corona. Se debe tomar en cuenta que la mayor parte del material aquí descrito corresponde únicamente a fragmentos o puntas de dientes, por lo cual no fue posible obtener la mayor parte de las medidas para cada uno de los ejemplares.

La medición de los dientes mayores a 10 mm se hizo con un vernier, mientras que para los dientes menores a un centímetro y los dentículos se utilizó un microscopio Olympus modelo SZ61. Las medidas se dan en milímetros.
El material se encuentra depositado en la Colección del Museo de Paleontología del Instituto de Geología, Universidad Nacional Autónoma de México.
Abreviaturas: mm, milímetros; IGM, Museo de Paleontología del Instituto de Geología; CH, altura de la corona; CBL, longitud de la base de la corona; CBW, anchura de la base de la corona; AL, longitud apical; MB/MC/MA, densidad denticular basal, media, apical de la carena mesial; DB/DC/DA, densidad denticular basal, media, apical de la carena distal; Ma, millones de años; V–, material reportado por Sankey (2001).
PALEONTOLOGÍA SISTEMÁTICA
Orden Saurischia Seeley, 1887
Suborden Theropoda Marsh, 1881
Familia Dromaeosauridae Matthew y Brown, 1922
Saurornitholestes sp. Sues, 1978
Material. IGM 6199 e IGM 6200. Fragmentos de diente.
Localidad. Rancho "Las Garzas", Municipio de Ocampo, Coahuila, México.
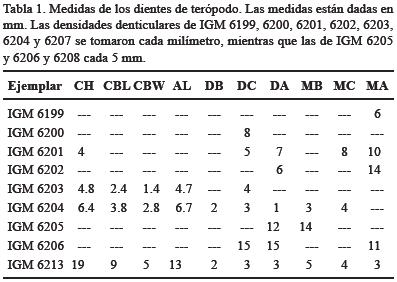
Descripción. Fragmentos de diente pequeños y aplanados labiolingualmente. Presentan dentículos delgados, los cuales pueden ser cortos con puntas redondeadas o bien, alargados y puntiagudos. Los dentículos distales siempre son más grandes que los mesiales. Los dentículos mesiales, cuando están presentes, son diminutos, rectos, y estrechos (Figura 4a–c). Las densidades denticulares de ambos ejemplares se encuentran citadas en la Tabla 1.
Discusión. El género Saurornitholestes fue descrito originalmente por Sues (1978) en la Formación Judith River, en Alberta, Canadá. La especie tipo es Saurornitholestes langstoni. Sankey et al. (2005) reportaron la presencia de este género en la fauna del Parque Nacional del Big Bend y parte del material fue conferido a la especie Saurornitholestes langstoni ; además describieron tres nuevos morfotipos de dientes conferidos a este género: cf. Saurornitholestes n. sp.? A, cf. Saurornitholestes n. sp.? B y cf. Saurornitholestes n. sp.? C.
IGM 6199 y 6200 se asignan a Saurornitholestes ya que presentan una forma de D en vista lateral, se encuentran fuertemente comprimidos labiolingualmente y la carena mesial se encuentran en la línea media del diente (Baszio, 1997b; Brinkman, 2002). Además, se ha observado que los dentículos distales en el género Saurornitholestes presentan un amplio rango de variabilidad en cuanto a su forma (Baszio 1997b) y que dicha variación denticular va desde rectangular, en forma de "U" hasta una forma de gancho. Sin embargo, a pesar de dicha variación morfológica, los dentículos de Saurornitholestessiempre conservan bases con forma rectangular y nunca están redondeadas como las de Troodon (Baszio, 1997b). En el ejemplar IGM 6199, aunque la carena distal se encuentra fracturada, es posible observar que ésta se encuentra caracterizada por la presencia de dentículos muy pequeños, con bases rectangulares y puntas redondeadas.
Currie et al. (1990) observaron que los dientes de Saurornitholestespresentan una fuerte diferencia en el tamaño denticular entre las carenas mesial y distal; siendo los dentículos mesiales diminutos mientras que los distales son más grandes, pero más pequeños que los de Troodon y más alargados y puntiagudos que los de Dromaeosaurus . El ejemplar IGM 6200 presenta dentículos alargados, rectos, estrechos y puntiagudos similares a los dentículos distales observados y descritos para Saurornitholestes . Aunque estos dentículos son considerablemente más alargados y más puntiagudos que los presentes en la carena distal del ejemplar IGM 6199, éstos no son tan largos ni presentan la típica morfología de gancho descrita para el género Troodon (Currie, 1987; Currie et al., 1990; Fiorillo y Currie, 1994; Baszio, 1997a; Peng et al., 2001).
En adición, en ambos ejemplares se observó la presencia de surcos sanguíneos, los cuales son más pronunciados que en Dromaeosaurus Matthew y Brown, 1922, aunque éstos tienen la misma orientación (paralela al eje longitudinal de los dentículos). Sin embargo, los dientes de Saurornitholestesson diferentes a los de Dromaeosaurus ya que nunca muestran la torsión característica de la carena mesial en la superficie lingual (Currie et al., 1990; Fiorillo y Currie 1994; Brikman, 2002).
Saurornitholestes langstoni Sues, 1978
Saurornitholestes langstoni , Sues, 1978, p. 382, figs 1–3. láms. 1–6.
Material. IGM 6201, diente (Figura 4d–e).
Localidad. Rancho "Las Garzas", Municipio de Ocampo, Coahuila, México.
Descripción. El ejemplar IGM 6201 tiene un grado de preservación relativamente bueno, se encuentra fracturado en su parte basal de tal modo que tiene una apariencia triangular en vista lateral y se encuentra labiolingualmente comprimido. Presenta dentículos en ambas carenas que corren a lo largo de la línea media del diente. El tamaño de los dentículos aumenta en ambas carenas conforme éstos se acercan hacia la parte basal del diente. Esto es más evidente en la carena distal. Los dentículos distales tienen puntas en forma de gancho y son el doble de grandes que los de la carena mesial. Se pueden observar espacios interdenticulares a pesar de que los dentículos se encuentran muy juntos entre sí. Por otro lado, los dentículos mesiales están inclinados hacia la punta del diente y presentan una forma de U, pero son más simétricos y redondeados que los de la carena distal y apenas se perciben espacios interdenticulares (Currie et al., 1990: fig. 8.2, 8.3 N; Sankey, 2001). Las medidas de este ejemplar y sus densidades denticulares se citan en la en la Tabla 1.
Discusión. Este ejemplar concuerda con la descripción y tamaño dados para la especie Sues (1978) y descritos por Currie et al. (1990). Presenta el carácter diagnóstico de la especie Saurornitholestes langstoni que es presentar dentículos distales con puntas en forma de gancho, los cuales son el doble de grandes (en promedio presentan un largo de 0.23 mm) que los de la carena mesial (en promedio presentan un largo de 0.12 mm).
Saurornitholestes n. sp.? A Sankey, Standhardt y Schiebout, 2005
Material. IGM 6202. Ápice de diente.
Localidad. Rancho "Las Garzas", Municipio de Ocampo, Coahuila, México.
Descripción. El ejemplar IGM 6202 (Figura 4f) corresponde al ápice de un diente con un estado de preservación relativamente bueno. Es un elemento labiolingualmente comprimido. Tanto la carena mesial como la distal presentan dentículos que se extienden a lo largo de ambas carenas y corren por la línea media del diente.
Los dentículos distales son más grandes que los mesiales; son rectangulares y tienen base en forma de cincel. Los dentículos que se encuentran completos presentan ápices ligeramente puntiagudos. Estos dentículos van haciéndose más grandes conforme se acercan a la base del diente. Además, al igual que los dentículos mesiales, los distales también reducen su tamaño conforme se acercan a la punta. Por otro lado, los dentículos mesiales son considerablemente más pequeños que los distales; son cortos, con bases rectangulares y ápices redondeados. Conforme se acercan a la punta del diente, los dentículos disminuyen en tamaño y se inclinan hacia el ápice del diente. Se observan con claridad espacios interdenticulares aunque éstos son muy estrechos, lo que hace que los dentículos estén muy juntos entre sí. Las densidades denticulares de este ejemplar se citan en la Tabla 1.
Discusión. Sankey et al. (2005) refirieron como cf. Saurornitholestes n. sp.? A a dientes similares a Saurornitholestes cf. langstoni pero que poseen dentículos mesiales, siendo los distales menos puntiagudos. En IGM 6202 los dentículos mesiales son más pequeños en comparación con los distales (Figura 4f), pero están más desarrollados que los de Saurornitholestes langstoni . Los dentículos distales presentan ápices ligeramente puntiagudos, sin embargo, éstos no están tan desarrollados como los de Saurornitholestes langstoni. Sankey et al. (2005) obtuvieron una densidad denticular de 4 a 9 dentículos cada milímetro en la carena distal de los dientes asignados a este morfotipo, los cuales presentan una altura que va de 0.17 a 0.23 mm y una anchura de 0.17 a 0.23 mm. Debido a que el ejemplar IGM 6202 únicamente corresponde al ápice del diente solamente fue posible obtener las densidades denticulares apicales tanto de la carena distal (6 dentículos cada milímetro) como de la mesial (14 cada milímetro). Además se observó que la altura y el ancho en promedio de los dentículos de la carena distal fue de 0.11 mm; siendo en la carena mesial de 0.08 mm y 0.06 mm respectivamente. Si tomamos en cuenta el conteo denticular de la carena distal (seis dentículos por 1 mm en la parte apical) vemos que cae dentro del rango establecido para el morfotipo cf. Saurornitholestes n. sp.? A. Además, los dentículos miden igual de alto que de ancho siendo que los dentículos de las carena distal de los dientes descritos por Sankey et al., (2005) siguen este mismo patrón en cuanto a la proporción de estas medidas. A pesar de que Sankey et al. (2005) no especificaron de cual parte de la carena distal obtuvieron su conteo denticular, en el presente trabajo se asigna el ejemplar IGM 6202 a cf. Saurornitholestes n. sp.? A debido a la similitud morfológica de los dentículos previamente explicada.
Saurornitholestes n. sp.? C Sankey, Standhardt y Schiebout, 2005
Material. IGM 6203. Diente (Figura 4g–h).
Localidad. Rancho "Las Garzas", Municipio de Ocampo, Coahuila, México.
Descripción. El ejemplar IGM 6203 tiene un estado de preservación bueno pero se encuentra fracturado en su ápice (Figs. 4g–h). Es un elemento labiolingualmente comprimido y su sección transversal es redondeada. La carena distal está aserrada mientras que la carena mesial carece de dentículos. En vista lateral se observa un borde en la carena mesial que corre desde el ápice hasta la parte media del diente (se observa como una línea café). Ambas carenas corren por la parte media del diente. Los dentículos distales son largos y delgados, en general se encuentran muy fracturados y únicamente se encuentran preservados en parte media de la carena distal; en uno de ellos se puede observar que la punta poseía un contorno redondeado. Se puede apreciar espacios interdenticulares. Las medidas de este diente y su densidad denticular se citan en la Tabla 1.
Discusión. Sankey et al. (2005) describieron el morfotipo cf. Saurornitholestes n. sp.? C el cual corresponde a un diente labiolingualmente comprimido que carece de dentículos anteriores y cuyos dentículos posteriores son alargados, delgados y con puntas redondeadas características morfológicas que presenta el ejemplar IGM 6203, por lo que este elemento se asigna a este taxón.
Familia Troodontidae, Gilmore, 1924
Género Troodon Leidy, 1856
cf. Troodon sp.
Material. IGM 6204. Diente premaxilar.
Localidad. Rancho "Las Garzas", Municipio de Ocampo, Coahuila, México.
Descripción. IGM 6204 presenta un buen estado de preservación, sin embargo, se encuentra fracturado en el ápice (Figura 4i). Su sección basal es elíptica. Este ejemplar corresponde a un diente premaxilar ya que la cara labial es convexa mientras que la lingual es cóncava; además, ambas carenas se encuentran aserradas y recorridas hacia la cara posterior (lingual) del diente. Los dentículos son largos con bases redondeadas y presentan una forma de gancho. A pesar de que únicamente una de las carenas está completa, es posible apreciar las diferencias entre dentículos mesiales y distales. Con base en el contorno de su carena más completa y en la inclinación de sus dentículos se sugiere que éstos pertenecen a la carena distal. Las medidas de este ejemplar y sus densidades denticulares se citan en la Tabla 1.
Discusión. Los dientes de troodóntidos poseen dentículos muy largos y puntiagudos en la carena distal; los dentículos de la carena mesial pueden tener un desarrollo variable o estar ausentes (Sankey et al., 2002). Los dientes tienen una constricción en la base de la corona; su forma es variable a lo largo de la hilera dental. Los dientes premaxilares y maxilares muestran diferencias en su morfología con lo que es posible distinguirlos. El ejemplar IGM 6204 presenta dentículos fuertemente alargados con forma de gancho, bases denticulares redondeadas, ambas carenas se encuentran en la cara lingual del diente, la sección transversal es triangular y presenta uno a dos dentículos cada milímetro en ambas carenas, características de los dientes premaxilares de Troodon (Currie et al.,1990; Sankey et al., 2002).
Los dentículos de los troodóntidos se caracterizan por ser los más grandes dentro de los terópodos juditianos, con un diámetro basal de hasta de 0.7 mm y hasta dos dentículos por milímetro.
Cabe destacar que los dientes de los troodóntidos, en algunas ocasiones, pueden llegar a confundirse con los dientes de anquilosaurios, ya que estos últimos presentan una serie de cúspides apicales separadas por surcos en el margen de la corona, que pueden ser identificadas erróneamente como dentículos (Baszio, 1997b; Brinkman, 2002); sin embargo, a diferencia de los dientes de Troodon , las cúspides de los anquilosaurios no adoptan la forma de gancho y los surcos corren verticalmente hacia la base del diente.
Familia Tyrannosauridae Osborn, 1905
Material. Ocho ápices de dientes IGM 6205, 6206, 6207, 6208, 6209, 6210, 6211, 6212.
Localidades. IGM 6206, 6207, 6208 y 6209 provienen del Rancho "Las Garzas". Municipio de Ocampo, Coahuila, México; IGM 6205, 6210, 6211, 6212 del El Ejido "El Mezquite", Cuenca de Sabinas, Coahuila, México.
Descripción. Ápices de dientes labiolingualmente comprimidos los cuales pueden presentar la cara lingual aplanada y la cara labial ligeramente convexa (IGM 6207), mientras que otros presentan ambas caras ligeramente convexas (IGM 6205 y 6210). Su sección transversal puede ser elíptica o cuadrangular. Ambas carenas se encuentran aserradas. Las carenas (mesial y distal) pueden correr por la línea media del elemento, o bien, la carena mesial puede comenzar cerca de la línea media del diente; pero a medida que se aleja hacia la base de éste desviarse hacia la cara labial (IGM 6207 y 6208), desviarse hacia la cara lingual (IGM 6205) o simplemente correr más cercana a la cara labial del diente (IGM 6211). Por otro lado, la carena distal puede correr a través de la parte media del diente, o correr más pegada hacia la cara labial del diente y conforme se aleja hacia la base se detiene como es el caso del ejemplar IGM 6207.
Los dentículos, de manera general, son anchos y gruesos, sus bases son cuadradas y sus puntas se encuentran aplanadas. Existen espacios interdenticulares claros, los cuales son más evidentes en la carena mesial (esta característica es más notoria en los ejemplares IGM 6206 y 6208; Figura 5d–e). Es posible observar la presencia de surcos sanguíneos entre los dentículos, los cuales se observan con mayor claridad en la cara lingual de la carena distal. Las densidades denticulares de los ejemplares IGM 6205 y 6206 se citan en la Tabla 1.
Discusión. Los dientes de tiranosáuridos en general se pueden identificar debido a su gran tamaño, y a la presencia de una corona alargada y recurvada. Los dientes de tiranosáuridos juveniles son básicamente versiones a escala de los grandes; de tal manera que éstos son más gruesos que los de los terópodos pequeños que presentan dientes de tamaño equivalente (Peng et al., 2001). Algunos dientes pueden ser bastante robustos como los pertenecientes a Tyrannosaurus rex Osborn, 1905 o bien, pueden ser un poco más gráciles como los de Gorgosaurus libratus (antes Albertosaurus libratus Lambe, 1914). Existen dentículos en ambas carenas y ambos son equivalentes en el tamaño. Son dentículos largos, rectangulares o cuadrados con extremos apicales ligeramente redondeados o aplanados, más anchos labiolingualmente que largos próximo distalmente y se presentan en intervalos de alrededor de tres dentículos por milímetro (Currie et al., 1990; Sankey, 2001).
Dentro de las características diagnósticas que definen los dientes de los tiranosáuridos se encuentran la sección transversal (la cual es la más grande de todos los grupos de terópodos) y la presencia de surcos sanguíneos bien definidos localizados entre los dentículos, los cuales se extienden oblicuamente hacia la base del diente (Currie et al.,1990; Abler, 1992). Estos particularmente se presentan en la cara lingual de la carena distal y es un carácter notorio en los ápices de dientes previamente descritos.
Los ejemplares IGM 6205, 6206, 6207, 6208, 6209, 6210, 6211 y 6212 presentan la morfología típica de dientes de Tyrannosauridae y aunque todos los ejemplares son partes apicales de dientes, las longitudes de estas partes van de 23 a 70 mm; esto sugiere que estos ápices corresponderían a dientes de terópodos de gran tamaño, tales como aquellos encontrados en Tyrannosauridae.
Los géneros de tiranosáuridos que se han encontrado en América del Norte son entre otros Tyrannosaurus, Daspletosaurus, y Gorgosaurus (antes Albertosaurus libratus Lambe, 1914) (Carr y Williamson, 2000; Holtz, 2004). Se ha visto que los dientes de Tyrannosaurus son muy robustos, y pueden superar los 280 mm de longitud, tienen carenas aserradas y una sección transversal redondeada; además los dentículos son muy robustos, cuadrados con puntas aplanadas; presentan seis dentículos por 5 mm (Lambe, 1917; Ostrom, 1969); Russell, 1970), que no es el caso de los dientes de Coahuila. Por otro lado, Albertosaurus y Daspletosaurus presentan dientes gráciles de más de 120 mm de longitud, los cuales poseen una sección transversal elíptica la cual llega a ser rectangular hacia la base de la superficie del diente. Tanto la carena mesial como la distal corren rectas a lo largo de la línea media del diente.
La identificación de taxa dentro de los Tyrannosauridae a través del material dental se ha basado principalmente en la densidad denticular de sus carenas. Sin embargo, esta identificación puede ser problemática (Carr y Williamson, 2000) debido a que los intervalos de densidad denticular pueden traslaparse entre los taxa estudiados. Por ejemplo Russell (1970) encontró que en los dientes de Tyrannosauridae los intervalos de densidad denticular de Daspletosaurus (11.5–13 dentículos cada 5 mm) se traslapaban con los de Albertosaurus (9–11 dentículos cada 5 mm). Así, un diente dado, no puede ser referido a un taxón específico ya que dependiendo de la longitud de la corona o del punto de la carena en donde se lleve a cabo el conteo de los dentículos la identificación puede volverse ambigua (Carr y Williamson, 2000). Por otro lado, Carr y Williamson (2000) estudiaron dientes de Tyrannosauridae diferenciando un conteo denticular apical, medio y basal para cada una de las carenas en un intervalo de 5 mm. Sus resultados arrojaron que las densidades denticulares de Daspletosaurus (10–23 dentículos cada 5 mm) se traslapan con las de Albertosaurus (10–17 dentículos cada 5 mm) al igual que lo que encontró Russell (1970). Sin embargo, Carr y Williamson (2000) propusieron que la densidad denticular puede ser útil para la identificación de dientes de Tyrannosaurus rex y confirieron dientes a esta especie con una densidad denticular apical de 7.5 dentículos cada 5 mm y una densidad denticular media de 8.5 dentículos cada 5mm en dientes del dentario.
Los ejemplares IGM 6206, 6207, 6208, 6209, 6210, 6211 y 6212 presentan densidades denticulares apicales que van de 8 a 12 dentículos cada 5 mm en la carena mesial y densidades denticulares apicales que van de 9 a 15 dentículos cada 5 mm en la carena distal. Así, esto ejemplares caen dentro de los rangos denticulares propuestos para Albertosaurus y Daspletosaurus . Debido a dicha problemática y a que no se tienen las coronas completas de los ejemplares aquí estudiados se decidió asignar estos ejemplares sólo a la familia Tyrannosauridae.
Por otro lado, es importante mencionar que IGM 6205 (Figura 5a–c) comparte una morfología muy similar a la forma descrita como Aublysodon por Molnar (1978) y Molnar y Carpenter (1989) para dientes muy recurvados hacia la punta, donde la carena mesial se desvía hacia la cara lingual conforme se acerca hacia la base del diente (Figura 5b). Sin embargo, la forma de los dentículos de IGM 6205 es un poco diferente a la descrita para Aublysodon, ya que éstos en IGM 6205 son más altos que anchos pero sí tienen forma cilíndrica y se inclinan suavemente hacia la punta del diente como sucede en Aublysodon (Molnar, 1978; Molnar y Carpenter, 1989). Actualmente Aublysodon es considerado como nomen dubium por considerarse que son dientes de organismos juveniles de Tyrannosauridae (Carr y Williamson, 2004; Holtz, 2004), por lo que se sugiere que el ejemplar IGM 6205 pertenece a un terópodo juvenil de Tyrannosauridae.
Familia Indeterminada
Género Indeterminado
Material. IGM 6213, diente.
Localidad. Rancho "Las Garzas" Municipio de Ocampo, Coahuila México.
Descripción. IGM 6213 (Figura 5f–h) es un diente labiolingualmente comprimido que se encuentra fracturado en su parte basal y en el ápice. Presenta dentículos tanto en la carena mesial como en la distal. Las carenas corren por la parte media del diente y conforme se acercan a la base de éste comienzan a desviarse hacia la cara labial de éste. El elemento se encuentra fracturado en el extremo proximal de la cara labial y en su margen distal (en el extremo distal). El margen mesial está más curvado que el margen distal. Su sección basal es elíptica. Los dentículos distales son más largos que los mesiales; son más altos que anchos y presentan una forma rectangular. Se encuentran redondeados en sus puntas. Existe un espacio claro entre un dentículo y otro. Los dentículos mesiales son más pequeños que los distales y al igual que éstos tienen una forma rectangular pero parecen medir igual de altos que de anchos; los espacios interdenticulares mesiales son más estrechos. Las medidas de este diente se mencionan en la Tabla 1.
Discusión. IGM 6213 presenta una morfología muy similar al género Dromaeosaurus (Currie et al., 1990; Baszio, 1997b; Sankey, 2001), es decir, se trata de un diente largo, robusto y recurvado, con dentículos cuyas puntas son redondeadas, sin embargo, a diferencia de Dromaeosaurus , no presenta la carena mesial torcida hacia la región lingual, que es una característica diagnóstica para el género. Por otro lado, IGM 6213 se diferencia del género Saurornitholestesen que los dientes de este género, son más pequeños, más recurvados y aplanados labiolingualmente y poseen dentículos fuertemente puntiagudos.
Sankey (2001) recolectó en sedimentos de la Formación Aguja en el área de Talley Mountain, (en el Parque Nacional del Big Bend, Texas), unos dientes (ejemplares V–5483 y V–6239) que son muy similares a IGM 6213. Los ejemplares V–5483 y V–6239 son fragmentos largos y recurvados cuyos CBL son 9.0 mm y 5.5 mm respectivamente. Sus dentículos son pequeños (0.2 mm de alto y 0.1 mm de ancho) y presentan de 3.5 a 5.5 dentículos por milímetro. El IGM 6213 tiene un CBL de 9.0 mm. Los dentículos son pequeños (0.2 mm a 0.24 mm de altura y de 0.16 a 0.2 mm en ancho) y presentan las densidades denticulares citadas en la tabla 1, por lo cual podemos ver que existe una similitud entre IGM 6213 y los ejemplares V–5483 y V–6239 reportados por Sankey (2001) quien sugirió que éstos podrían corresponder a un nuevo taxón debido a que la morfología de estos dientes no correspondían a ninguno de los géneros de terópodos descritos anteriormente.
Por otro lado, se sabe que los dentículos de los dientes en Tyrannosauridae son equivalentes en tamaño en ambas carenas y que éstos pueden ser largos, rectangulares o cuadrados con extremos apicales ligeramente redondeados (Currie et al., 1990; Sankey, 2001). Puesto que el ejemplar IGM 6213 presenta dentículos con puntas redondeadas podría suponerse que pudiera pertenecer a un organismo juvenil de Tyrannosauridae o bien a un terópodo de tamaño pequeño tal como Saurornitholestessi consideramos que la redondez de los dentículos pudiera deberse al desgaste o a la acción de la intemperie . Esta idea estaría sustentada además por la diferencia del tamaño denticular entre las carenas distal y mesial que existe en IGM6213.
Debido a que no se observan características diagnósticas y que se trata de un solo ejemplar hemos decidido dejar a IGM6213 simplemente como un terópodo indeterminado.
DISCUSIÓN
Baszio (1997a), Currie et al. (1990), Sankey (2001) y Sankey et al. (2002, 2005) realizaron estudios comparando las asociaciones de terópodos a través del tiempo, desde el Campaniano tardío al Maastrichtiano tardío, los dos primeros en las faunas del norte (Alberta, Canadá) y Sankey y colaboradores en el Parque Nacional del Big Bend, en Texas.
En la fauna del norte, Baszio (1997a) encontró dos asociaciones de terópodos: una corresponde a la presente en las formaciones Dinosaur Park y Horseshoe Canyon que abarcan del Campaniano tardío al Maastrichtiano temprano, la cual se caracteriza por una abundancia de terópodos del género Troodon y por una escasa presencia del género Richardoestesia. La otra asociación, presente durante el Maastrichtiano tardío, se encuentra caracterizada por una elevada abundancia de los géneros Richardoestesia y "Paronychodon" (ver observación en párrafos anteriores).
En la fauna del sur, Sankey et al. (2005) también distinguieron dos asociaciones distintas de terópodos; la primera de ellas, corresponde del Campaniano tardío al Maastrichtiano temprano y la segunda pertenece al Maastrichtiano tardío. La asociación campaniana tardía–maastrichtiana temprana (Formación Aguja) se desarrolló en hábitats costeros y está caracterizada por una mayor abundancia de Saurornitholestes cf. langstoni y Richardoestesia isosceles Sankey, 2001. En cambio, la asociación maastrichtiana tardía (Formación Tornillo) se desarrolló en hábitats más terrestres y estaba caracterizada por una abundancia elevada de Saurornitholestes sp. y "Paronychodon".
Durante el Campaniano tardío, la composición de tiranosáuridos también presenta diferencias entre los elementos de las faunas del norte y del sur (Lehman, 1997; Carr y Williamson, 2000); aparentemente Daspletosaurus es el taxón más abundante en las faunas sureñas, mientras que la fauna del norte estuvo dominada por los géneros Gorgosaurus libratus (antes Albertosaurus libratus Holtz, 2004) y Daspletosaurus (Russell, 1970; Horner et al., 1992). En el Maastrichtiano temprano, Albertosaurus fue el tiranosáurido más abundante en la fauna del norte, mientras que en el sur el material de tiranosáurido es muy escaso y los únicos ejemplares recolectados también han sido referidos a Albertosaurus (Carr y Williamson, 2000). Durante el Maastrichtiano tardío, Tyrannosaurus rex se convirtió en el gran depredador residente del oeste de América del Norte (desde Alberta y Saskatchewan al sur de Canadá hasta Nuevo México y Texas en los Estados Unidos). Esta diferenciación de las faunas entre norte y sur también se reconoce en las asociaciones de hadrosaurios y ceratópsidos (Lehman, 2001). Wolfe y Upchurch (1987), Baghai (1996) y Lehman (1997) han propuesto que la diferencia de lluvia y temperatura fueron los factores principales que produjeron este patrón biogeográfico norte–sur.
La diversidad de pequeños terópodos hasta ahora identificada en Coahuila es comparable a la correspondiente del Campaniano tardío–Maastrichtiano temprano de Texas. En Coahuila los dientes de Saurornitholestes fueron los más abundantes; llama la atención que no se ha reconocido todavía la presencia de Richardoestesia que es común en la fauna texana.
La presencia del género Troodon (un diente premaxilar) en la localidad "Las Garzas" resulta importante, ya que dicho género solamente se había registrado en la provincia de Alberta (Canadá) así como en los estados de Montana y Wyoming (Estados Unidos) durante el Campaniano–Maastrichtiano y además era muy abundante (Baszio, 1997a). De esta manera, la presencia del género Troodon en el estado de Coahuila indicaría que este taxón tendría una distribución geográfica mucho más amplia.
En el estado de Coahuila se observa una asociación de dinosaurios terópodos compuesta principalmente por Saurornitholestes y tiranosáuridos, lo cual coincide con lo reportado para el Parque Nacional del Big Bend por Sankey et al. (2005). Es importante continuar con el estudio de dientes de terópodos pequeños en busca de los otros taxa reportados por Sankey et al. (2005) para establecer si la asociación de dinosaurios terópodos presente en el estado de Coahuila, México puede integrarse a la fauna del sur de las Grandes Planicies de América del Norte propuesta por Lehman (1997).
CONCLUSIONES
A pesar de las limitantes que existen, el estudio de dientes aislados de terópodos puede constituir una herramienta para la identificación taxonómica, inclusive hasta el nivel de género y en algunos casos de especie. Esto pudo ser aplicado principalmente a terópodos pequeños tales como Troodon y Saurornitholestespues en el caso de los terópodos grandes referidos a Tyrannosauridae no existe una constante denticular para poder identificar a nivel de género.
La metodología propuesta por Smith (2005, 2007) y Smith et al. (2005) puede ser útil para identificar terópodos y reconocer taxa a partir del estudio de la morfología de dientes aislados mediante la medición de diversas variables como la longitud de la corona, posición de las carenas, la densidad denticular, entre otros. A pesar de que dichas variables habían sido objeto de estudio previamente por diversos autores (ejemplo Farlow y Brinkman, 1987; Currie et al., 1990; Sankey et al., 2002, entre otros) éstas no habían sido estandarizadas para el grupo de terópodos. Se debe tomar en cuenta que la mayor parte de las medidas propuestas para los dientes aislados están diseñadas para coronas completas y en el registro fósil generalmente los dientes completos son raros o escasos y la identificación puede verse limitada principalmente a la morfología de los dentículos. En el caso de terópodos pequeños tales como Saurornitholesteso Troodon esto no sería una limitante pero en el caso de terópodos grandes como Tyrannosauridae la morfología denticular no es un carácter diagnóstico a nivel de género, por lo que es necesario seguir trabajando en una metodología que permita identificar taxa de Tyrannosauridae a partir de fragmentos de dientes.
La asociación de terópodos encontrada en el estado de Coahuila representada por Saurornitholestes , cf. Troodon , Tyrannosauridae y un género que podría corresponder a un nuevo taxón demuestra que dicho estado contaba con una amplia diversidad de terópodos durante el Cretácico Tardío, la cual comparte similitud con la registrada por Sankey et al. (2005) para el Parque Nacional del Big Bend, Texas.
AGRADECIMIENTOS
El sedimento para tamizar y la recolección de los fósiles fueron parte de los proyectos "Cretácico Tardío continental del norte de los estados de Coahuila y Chihuahua" financiado por la DGAPA IN 202802 (2003–2004) y "Exploring for Late Cretaceous vertebrates in northern Coahuila" apoyado por la National Geographic Society durante el año 2000. Parte de este trabajo es el resultado del proyecto de tesis de licenciatura realizado por la Biól. Esperanza Torres–Rodríguez (2006). Agradecemos el apoyo del Sr. Gerardo Álvarez–Reyes durante las temporadas de campo y el trabajo de laboratorio; a la M. en C. Claudia Serrano Brañas, quien fue la responsable de tamizar y revisar el sedimento para la separación de los microvertebrados; y en especial al Ing. Juan Pablo García por la donación del material del Ejido "El Mezquite". Agradecemos a la Dra. María del Carmen Perrilliat por la identificación del molusco recolectado en el área de Las Garzas. Extendemos nuestro agradecimiento a la comunidad de Manuel Benavides, Chihuahua, en especial al Sr. Benjamín Ortiz por la orientación y apoyo recibidos durante las temporadas de campo, así como al Sr. Baldemar Márquez, dueño del Rancho "Las Garzas" por la autorización para realizar las temporadas de campo en su propiedad; al personal del Área Protegida de Flora y Fauna del Cañón de Santa Elena por las facilidades prestadas; y a la Dra. Julia Sankey por la ayuda prestada durante la identificación del material fósil. No podemos dejar de agradecer a los Dres. Mathew Lamanna, José Ignacio Canudo y Robert McCord por la cuidadosa revisión y comentarios hechos al manuscrito.
REFERENCIAS
Abler, W.L., 1992, The serrated teeth of tyrannosaurid dinosaurs, and biting structures in other animals: Paleobiology, 18(2), 161–183. [ Links ]
Andrade–Ramos, P., Montellano–Ballesteros, M., Cevallos–Ferriz, S., Lozano–García, S., 2002, A Late Cretaceous association from Altares, Chihuahua, México: Journal of Vertebrate Paleontology, 22(3) (suppl.), 32A. [ Links ]
Baghai, N., 1996, An analysis of palynomorphs from Upper Cretaceous sedimentary rocks with emphasis on the Aguja Formation, Big Bend National Park, Brewster County, Texas: University of Texas, at Austin, tesis doctoral, 495 pp. [ Links ]
Baszio, S., 1997a, Systematic paleontology of isolated dinosaur teeth from the latest Cretaceous of south Alberta, Canada: Courier Forschungsinstitut Senckenberg, 196, 33–77. [ Links ]
Baszio, S., 1997b, Investigations on Canadian dinosaurs: palaeoecology of dinosaurs assemblages throughout the Late Cretaceous of southern Alberta, Canada: Courier Forschungsinstitut Senckenberg, 196, 1–31. [ Links ]
Benammi, M., Montellano–Ballesteros, M., 2006, New data from the continental Late Cretaceous faunas from northern México: Journal of Vertebrate Paleontology, 26(3) (suppl.), 41A. [ Links ]
Brinkman, D.B., 2002. An illustrated guide to the vertebrate microfossils from the Dinosaur Park Formation: Alberta Paleontological Society, Workshop on Vertebrate Microfossils, 141 pp. [ Links ]
Cabrera, F., Narváez, G., Chávez, J. M, Hernández, R., Alcántara, J., Gómez, F., 1984, Exploración carbonífera en la Cuenca de Ojinaga, Chihuahua: Boletín de la Sociedad Geológica Mexicana, 45(3), 41–61. [ Links ]
Carr, T.D., Williamson, T.E., 2000, A review of Tyrannosauridae (Dinosauria, Coelurosauria) from New Mexico, en Lucas, S.G., Heckert, A.B. (eds.), Dinosaurs of New Mexico: New Mexico Museum of Natural History and Science Bulletin, 17, 113–146. [ Links ]
Carr, T.D., Williamson, T.E., 2004, Diversity of late Maastrichtian Tyrannosauridae (Dinosauria: Theropoda) from western North America: Zoological Journal of the Linnean Society, 142, 479–523. [ Links ]
Currie, P.J., 1987, Bird–like characteristics of the jaws and teeth of troodontid theropods (Dinosauria, Saurischia): Journal of Vertebrate Paleontology, 7(1), 72–81. [ Links ]
Currie, P.J., 1988, Theropoda en Currie, Ph. J., Padian K. (eds.), The Dinosaur Enciclopedia: San Diego, Academic Press, 731–736. [ Links ]
Currie, P.J., Rigby, J.K, Jr. Sloan, R.E., 1990, Theropod teeth from the Judith River Formation of southern Alberta, Canada, in Carpenter, K., Currie, P.J. (eds.), Dinosaur Systematics: Approaches and Perspectives: Cambridge, Cambridge University Press, 107–125. [ Links ]
Dumble, E.T., 1982, Notes on the Geology of the Valley of the Middle Rio Grande: Geological Society of America Bulletin, 3, 219–232. [ Links ]
Farlow, J.O., Brinkman, D.L. 1987. Serration coarseness and patterns of wear of theropod dinosaur teeth: Geological Society of America, Abstracts with Programs, 19, 151. [ Links ]
Fastovsky, D.E., Huang, Y., Hsu, J., Martin–McNaughton, J., Sheehan, P.M., Weishampel, D.B., 2004, Shape of Mesozoic dinosaur richness: Geology, 32, 877–880. [ Links ]
Fastovsky, D.E., Weishampel, D.B., Sibbick, J., 2005, The evolution and extinction of the dinosaurs: Cambridge, Cambridge University Press, Segunda Edición, 550 pp. [ Links ]
Fiorillo, A.R., Currie, Ph.J., 1994, Theropod teeth from the Judith River Formation (Upper Cretaceous) of south–central Montana: Journal of Vertebrate Paleontology, 14(1), 74–80. [ Links ]
Gilmore, C.W., 1924, A new coelurid dinosaur from the Belly River Cretaceous of Alberta: Bulletin Canadian Department of Mines Survey, 38, 1–12. [ Links ]
Holtz T.R., 2004, Tyrannosauroidea, en Weishample, D.B., Dodson, P., Osmolska, H., (eds.), The Dinosauria: Berkeley, University of California Press, Segunda Edición, 111–136. [ Links ]
Horner, J.R., Varricchio, D.J., Goodwin, M.B., 1992, Marine transgressions and the evolution of Cretaceous dinosaurs: Nature, 258, 59–61. [ Links ]
Lambe, L.M., 1914, On a new genus and species of carnivorous dinosaur from the Belly River Formation of Alberta, with a description of the skull of Stephanosaurus marginatus from the same horizon: Ottawa Naturalist, 28, 13–20. [ Links ]
Lambe, L.M., 1917, The Cretaceous theropodous dinosaur Gorgosaurus: Geological Survey of Canada Memoir, 100, 1–84. [ Links ]
Lehman, T.M., 1987, Late Maastrichtian paleoenviroments and dinosaur biogeography in the Western Interior of North America: Palaeogeography, Palaeoclimatology, Palaeoecology, 60(1), 189–217. [ Links ]
Lehman, T.M., 1989, Chasmosaurus mariscalensis, sp. nov. a new ceratopsian dinosaur from Texas: Journal of Vertebrate Paleontology 9(1), 137–162. [ Links ]
Lehman, T.M., 1997, Late Campanian dinosaur biogeography in the Western Interior of North America, in Wolberg, D.L., Stump, E., Rosenberg G. D. (eds.), Dinofest International; Proceedings of a Symposium Sponsored by Arizona State University: Philadelphia, Academy of Natural Sciences, Special Publication, 223–240. [ Links ]
Lehman, T.M., 2001, Late Cretaceous dinosaur provinciality, en Tanke, D.H., Carpenter, K. (eds.), Mesozoic Vertebrate Life: Bloomington, Indiana University Press, 310–328. [ Links ]
Leidy, J., 1856, Notices of remains of extinct reptiles and fishes, discovered by Dr. F.V. Hayden in the bad lands of Judith River, Nebraska Territory: Proceedings of the Academy of Natural Sciences of Philadelphia, 8, 72–73. [ Links ]
Lucas, S.G., Kues, B.S., González de León, C., 1995, Paleontology of the Upper Cretaceous Cabullona Group, northeastern Sonora, en Jacques–Ayala, C., González de León, C., Roldán–Quintan, J., (eds.), Studies on the Mesozoic of Sonora and Adjancent Areas: Boulder, Colorado, Geological Society of America Special Paper, 301, 143–165. [ Links ]
Makovicky, P.J., Norell, M.A., 2004, Troodontidae, en Weishample, D.B., Dodson, P., Osmolska, H., (eds.), The Dinosauria: Berkeley, University of California Press, Segunda edición, 184–195. [ Links ]
Marsh, O.C., 1881, A new order of extinct Jurassic reptiles (Coeluria): American Journal Science (serie 3) 21, 339–340. [ Links ]
Matthew, W.D., Brown, B., 1922, The family Deinodontidae, with notice of a new genus from the Cretaceous of Alberta: Bulletin of the American Museum of Natural History, 46, 367–385. [ Links ]
Molnar, R.E., 1978, A new theropod dinosaur from the Upper Cretaceous of central Montana: Journal of Paleontology, 52(1), 73–82. [ Links ]
Molnar, R.E., Carpenter, K., 1989, The Jordan theropod (Maastrichtian, Montana, USA) referred to the genus Aublysodon: Geobios, 22(4), 445–454. [ Links ]
Montellano–Ballesteros, M., 2003, Titanosaurid sauropod in the Late Cretaceous of Chihuahua, México: Revista Mexicana de Ciencias Geológicas, 20(2), 160–164. [ Links ]
Montellano–Ballesteros, M., Hernández–Rivera, R., Alvarez–Reyes, G., Andrade–Ramos, P., Martín–Medrano, L., López J., 2000, Discovery of Late Cretaceous vertebrate local faunas in northern Mexico: Journal of Vertebrate Paleontology, 20(3) (suppl.), 58A. [ Links ]
Osborn, H., 1905, Tyrannosaurus and other Cretaceous carnivorous dinosaurs: Bulletin of American Museum of Natural History, 21,259–265. [ Links ]
Ostrom, J.H., 1969, A new theropod dinosaur from the Lower Cretaceous of Montana: Postilla, 128, 1–17. [ Links ]
Peng, J., Russell, A.P., Brinkman, D.B., 2001, Vertebrate microsite assemblages (exclusive of mammals) from the Foremost and Oldman formations of the Judith River Group (Campanian) of southeastern Alberta: an illustrated guide: Edmonton, Provincial Museum of Alberta, Natural History Occasional Paper, 25, 54 pp. [ Links ]
Russell, D.A., 1970, Tyrannosaurs from the Late Cretaceous of western Canada: National Museum of Natural Science Publications in Palaeontology, 1, 1–34. [ Links ]
Sankey, J.T., 2001, Late Campanian southern dinosaurs, Aguja Formation, Big Bend National Park, Texas: Journal of Paleontology, 75(1), 208–215. [ Links ]
Sankey, J.T., Brinkman, D.B., Guenther, M., Currie, P. J., 2002, Small theropod and bird teeth from the Judith River Formation Group (late Campanian), Alberta: Journal of Paleontology, 76(4), 751–763. [ Links ]
Sankey, J.T., Standhardt, B.R., Schiebout, J.A., 2005, Theropod teeth from the Upper Cretaceous (Campanian–Maastrichtian), Big Bend National Park, Texas, en Carpenter, K. (ed.), The Carnivorous Dinosaurs: Bloomington, Indiana University Press, 127–152. [ Links ]
Say, T., 1820, Observations on some species of zoophytes, shells, etc. principally fossil: The American Journal of Science and Arts, 2, 234–45. [ Links ]
Seeley, H.G., 1887, On the Classification of the fossil animals commonly named Dinosauria: Proceedings of the Royal Society of London, 43, 165–171. [ Links ]
Smith, J.B., 2005, Heterodonty in Tyrannosaurus rex: implications for the taxonomic and systematic utility of theropod dentitions: Journal of Vertebrate Paleontology, 25(4), 865–887. [ Links ]
Smith, J.B., 2007, Dental morphology and variation in Majungasaurus crenatissimus (Theropoda: Abelisauridae) from the late Cretaceous of Madagascar: Journal of Vertebrate Paleontology, 27(2), 103–126. [ Links ]
Smith, J.B., Vann, D.R, Dodson, P., 2005, Dental morphology and variation in theropod dinosaurs: implications for the taxonomic identification of isolated teeth: The Anatomical Record, Part A 285A, 699–736. [ Links ]
Stephenson, L.W., 1941, The larger invertebrate of the Navarro Group of Texas: The University of Texas Publications, 4101, 1–641. [ Links ]
Sues, H.–D., 1978, A new small theropod dinosaur from the Judith River Formation (Campanian) of Alberta, Canada: Zoological Journal of the Linnean Society, 62, 381–400. [ Links ]
Torres–Rodríguez, E., 2006, Terópodos del Cretácico Superior del Estado de Coahuila, México: México, Universidad Nacional Autónoma de México, Facultad de Ciencias, Tesis de licenciatura 91 pp. [ Links ]
Wolfe, J.A., Upchurch, G.Jr., 1987, North American non marine climates and vegetation during the Late Cretaceous: Palaeogeography, Palaeoclimatology, Palaeoecology, 61,33–77. [ Links ]

