










Serviços Personalizados
Journal

Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
 Citado por SciELO
Citado por SciELO Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkRevista mexicana de ciencias geológicas
versão On-line ISSN 2007-2902versão impressa ISSN 1026-8774
Rev. mex. cienc. geol vol.26 no.3 Ciudad de México Dez. 2009
Los tiburones Lamniformes (Chondrichthyes, Galeomorphii) del Plioceno inferior de la Formación Arenas de Huelva, suroeste de la cuenca del Guadalquivir, España
The lower Pliocene lamniform sharks (Chondrichthyes, Galeomorphii) from the Arenas de Huelva Formation, southwestern Guadalquivir basin, Spain
Edith Xio Mara García1 *, Miguel Telles–Antunes2, 3 , Ausenda Cáceres–Balbino2, 4 , Francisco Ruiz–Muñoz 5 y Jorge Civis–Llovera 6
1 Departamento de Botánica y Zoología, Centro Universitario de Ciencias Biológicas y Agropecuarias, Universidad de Guadalajara, Km 15.5 carretera Guadalajara–Nogales, Apartado postal 1–139, 45110 Las Agujas, Zapopan, Jalisco, México. * edithxiomara@gmail.com
2 Centro de Investigação em Ciência e Engenharia Geológica, Departamento de Ciências da Terra, Faculdade de Ciências e Tecnologia da Universidade Nova de Lisboa, Quinta da Torre, 2829–516 Caparica, Portugal.
3 Academia das Ciências de Lisboa, Rua da Academia das Ciências, 19, 1249–122 Lisboa, Portugal.
4 Departamento de Geociências, Universidade de Évora, Apartado 94, 7002–554 Évora, Portugal.
5 Departamento de Geodinámica y Paleontología, Facultad de Ciencias Experimentales, Universidad de Huelva, Av. de las Fuerzas Armadas s/n, 21071 Huelva, España.
6 Área de Paleontología, Departamento de Geología, Universidad de Salamanca, Plaza de la Merced, s/n, 37008 Salamanca, España.
Manuscrito recibido: Agosto 15, 2008
Manuscrito corregido recibido: Julio 27, 2009
Manuscrito aceptado: Agosto 10, 2009
RESUMEN
Se estudiaron los dientes de Lamniformes provenientes de ocho localidades de la Formación Arenas de Huelva, Plioceno inferior del suroeste de la cuenca del Guadalquivir, España. Se registraron los géneros Carcharias, Alopias, Isurus, Carcharocles y Parotodus. Se observó un predominio de restos de Carcharias taurus, los demás taxones son raros. Las asociaciones de seláceos indican que estos sedimentos fueron depositados en un ambiente litoral–nerítico, con una temperatura de aguas subtropicales a templadas.
Palabras clave: lamniformes, Paleoecología, Plioceno inferior, Formación Arenas de Huelva, España.
ABSTRACT
Lamniform teeth from eight localities of the lower Pliocene Arenas de Huelva Formation were studied in the southwestern part of the Guadalquivir basin. The genera Carcharias, Alopias, Isurus, Carcharocles and Parotodus were recognized. Carcharias taurus remains prevail, the other taxons being scarce. The selachian assemblage points out to littoral–neritic, subtropical to temperate environments.
Key words: Lamniforms, Paleoecology, lower Pliocene, Arenas of Huelva Formation, Spain.
INTRODUCCIÓN
El Neógeno del suroeste de la cuenca del Guadalquivir ha sido estudiado por el equipo de investigación en Paleontología de la Universidad de Salamanca, así como por investigadores de la Universidad de Huelva y Sevilla, abordando el análisis de diversos grupos bien representados en este sector de la cuenca.
Civis et al. (1987) definieron tres formaciones geológicas ricas en restos fósiles dentro del Neógeno de la Cuenca del Guadalquivir, estas son: Formación Calcarenita de Niebla (Tortoniense superior), Formación Arcillas de Gibraleón (Tortoniense superior/Messiniense) y Formación Arenas de Huelva (Plioceno inferior). Posteriormente, Mayoral y Pendón (1987) definieron una formación aún más moderna, la cual fue nombrada Formación Arenas de Bonares (Plioceno–Cuaternario), que marca el tránsito marino–continental para el área (Figura 1).
La Formación Arenas de Huelva, que es en donde se enfoca este estudio, inicia con un nivel de glauconita de 2 a 4 m de espesor. Este nivel, rico en dientes de seláceos, es considerado como el marcador Mioceno–Plioceno (5.33 Ma) para el suroeste de la cuenca del Guadalquivir (González–Delgado et al., 2004). Suprayacente a este nivel, se encuentra una serie de arenas limosas masivas y bioturbadas con frecuentes intercalaciones lumaquélicas de moluscos, la génesis de estas acumulaciones de moluscos ha sido atribuida a niveles de tormentas (Dabrio et al., 1988).
Entre los trabajos paleontológicos realizados en la década de 1980, destacan las tesis doctorales en foraminíferos (Sierro, 1984; González–Regalado, 1986), malacofauna (Andrés, 1982; González–Delgado, 1983) y nanoplancton calcáreo (Flores, 1985).
Entre los estudios que quedaban pendientes en el Neógeno del suroeste de la cuenca del Guadalquivir estaba el estudio de los Condrictios y Osteíctios, por lo que se elaboró una tesis doctoral abordando estos temas (García, 2008). En ella se determinaron un total de 31 taxones de seláceos a partir de restos dentarios, distribuidos en seis órdenes. La mayor diversidad se encontró en el orden Carcharhiniformes (32%), seguida de los Lamniformes (26%), Myliobatiformes (23%), Rajiformes (10%), Hexanchiformes (6%) y Squaliformes (3%).
El presente trabajo está basado en datos previamente presentados en la Tesis Doctoral de García (2008).
METODOLOGÍA
El material fósil que se estudió en este trabajo se encuentra depositado en la Colección de Paleontología de la Universidad de Salamanca, España, su acrónimo es PALUSAL.
Las determinaciones y descripciones se realizaron a través del estudio de restos dentarios. Se compararon con dientes de especies actuales de Carcharias taurus (Rafinesque, 1810), Alopias superciliosus (Lowe, 1841), Alopias vulpinus (Bonnaterre, 1788) e Isurus oxyrinchus (Rafinesque, 1810), que se localizan en la colección de Paleontología de la Universidad de Évora, Portugal. Asimismo, se compararon con dientes fósiles de Carcharias taurus (Rafinesque, 1810), del Plioceno de Angola, que pertenecen a la colección de Paleontología de la Academia de Ciencias de Lisboa; el estudio de estos dientes se encuentra detallado en Antunes (1978). Además, se compararon con ejemplares de dientes fósiles del Mioceno terminal de la Cuenca de Alvalade, Portugal, como: Isurus desori (Sismonda, 1849), Isurus hastalis (Agassiz, 1843) y Carcharocles megalodon (Agassiz, 1843), los cuales fueron determinados y descritos en la tesis doctoral de Balbino (1995); este material se encuentra resguardado en el Laboratorio de Paleontología de la Universidad de Évora. Se contrastó con figuras de Parotodus benedeni (Le Hon, 1871), de los trabajos de Leriche (1926), Antunes (1978), Cappetta (1987), Ward y Galea (2001) y Cocke (2002).
Se colectaron muestras en ocho localidades del Neógeno de la provincia de Huelva, de las cuales se levigaron más de 300 kg de sedimentos utilizando tamices de 0.5 y 1 mm de luz de malla. Estos sitios de colecta se localizan en el sur de Huelva. Dentro de la población de Niebla se encuentran las localidades de Niebla–Cabezo de las Cruces (37°21'4.85''N; 6°36'44.68''W) y Cabezo de San Cristóbal (37°20'41.73''N; 06°39''41.65''W). En la población de Bonares están las localidades de Bonares–Casa del Pino (37°20'0.40''N; 6°40'33.63''W), Bonares–Ambulatorio (37°19'38.66''N; 6°40'37.39''W) y Bonares–B (37°19'7.63'' N; 6°41'8.53''W). En el pueblo de Moguer se encuentra la localidad Moguer–A (37°16'16.60''N; 6°50'33.53''W) y en Cartaya, la localidad El Rompido (37°13'2.77''N; 7°6' 4.80''W), (Figura 2).

PALEONTOLOGÍA SISTEMÁTICA
La clasificación adoptada se ha tomado de Cappetta (1987, 2006) y de Compagno (1984a, 1984b, 2005).
Clase Chondrichtyes Huxley, 1880
Subclase Elasmobranchii Bonaparte, 1838
Cohorte Euselachii Hay, 1902
Subcohorte Neoselachii Compagno, 1977
Superorden Galeomorphii Compagno, 1973
Orden Lamniformes Berg, 1958
Familia Odontaspididae Müller y Henle, 1839
Género Carcharias Rafinesque, 1810
Carcharias taurus (Rafinesque, 1810) (Figuras 3a–3g y 4a–4d)
Odontaspis taurus Rafinesque. Antunes, 1978, p. 67, lám. 2, figs. 16–25; Mañé, Magrans y Ferrer, 1996, p. 24, lám. 1, figs. 17–21; Mas, 2000, figs. 8.5.
Carcharias taurus Rafinesque. Cappetta y Cavallo, 2006, p. 41, lám. 2, fig. 2.
Descripción. Tres dientes anteriores (PALUSAL, P–C21, P–C63 y PC122); un diente superior–izquierdo, primera fila (PALUSAL, P–C5); un diente inferior–izquierdo, cuarta fila (PALUSAL, P–C105); un diente superior–derecho, tercera fila (PALUSAL, P–C62–2) y un diente lateral (PALUSAL, P–C14).
Los dientes anteriores se encuentran deteriorados del esmalte. Poseen unas delgadas cúspides, bordes cortantes en vista lateral y una forma sigmoidal. La cara lingual es convexa. Apenas se alcanzan a apreciar las estrías verticales debido al deterioro. La cara labial es algo convexa. En los tres ejemplares se han perdido las raíces y los dentículos laterales.
El diente superior procede de la primera fila izquierda y presenta el esmalte bien conservado, así como una cúspide delgada. Presenta bordes cortantes y perfil sigmoidal. La cara lingual es convexa con estrías verticales y la cara labial es convexa. La raíz se encuentra constituida por dos lóbulos separados. La protuberancia lingual esta muy desarrollada, con canal nutricio. Los bordes de los lóbulos se encuentran deteriorados. Los dentículos laterales se han perdido.
El esmalte del diente inferior izquierdo de la cuarta fila está deteriorado. La corona es de forma triangular. En la cara lingual, ligeramente convexa, apenas se aprecian las estrías. La cara labial es tenuemente convexa. En la vista lateral, se pueden ver los bordes cortantes. Los dentículos laterales son inclinados lingualmente y de base circular. La raíz posee los dos lóbulos bien separados. En la cara lingual de la raíz, la protuberancia lingual está muy desarrollada y presenta el canal nutricio. El lóbulo mesial es un poco más largo que el lóbulo distal.
El diente superior derecho de la tercera fila posee la cara lingual de la corona ligeramente convexa, con estrías bien marcadas. En la vista lateral, se aprecian los bordes cortantes. La cara labial es ligeramente convexa. La corona se encuentra ligeramente inclinada en dirección a la comisura. Existe un par de dentículos laterales inclinados lingualmente. La raíz posee unos lóbulos bien marcados con una protuberancia lingual muy desarrollada y un canal nutricio patente. El lóbulo mesial es más largo que el lóbulo distal.
El diente lateral posee la corona triangular, inclinada hacia la comisura y con una base ancha. En la cara lingual aparece estriación. En la vista lateral se pueden ver los bordes cortantes. Se ha perdido uno del par de dentículos laterales. La raíz se encuentra deteriorada, pero se aprecian dos lóbulos.
Material. Siete dientes. PALUSAL, P–C5, P–C14, P–C21, P–C62–2, P–C63, P–C105 y P–C122.
Localidad. Bonares–Ambulatorio, Bonares–Casa del Pino, Cabezo de San Cristóbal y Moguer–A, Formación Arenas de Huelva, Cuenca de Guadalquivir, España.
Discusión. Los fósiles arriba descritos fueron comparados con dientes de un ejemplar actual de Carcharias taurus. Lo anterior permite sugerir la posición dentro de las mandíbulas que algunos de los dientes fósiles debieron tener.
Se compararon con dientes fósiles de C. taurus del Plioceno de Angola, de la colección de paleontología de la Academia de Ciencias de Lisboa, determinados por Antunes (1978). Además se compararon con dientes fósiles de Carcharias acutissima de Esbarrondadoiro y Vale de Zebro (Mioceno terminal de la Cuenca de Alvalade, Portugal), de la Colección de Paleontología de la Universidad de Evora, descritos y determinados por Balbino (1995).
Se determinó que estos dientes pertenecen a la especie C. taurus por la forma sigmoidal de algunos de los ejemplares, por la estriación vertical en la cara lingual y además por la posesión de dentículos laterales.
Familia Alopiidae Bonaparte, 1838
Género Alopias Rafinesque, 1810
Alopias aff. vulpinus (Bonnaterre, 1788) (Figuras 4e–4g)
Alopias cf. vulpinus Bonnaterre. Antunes y Jonet, 1970, lám. 7, fig. 26; lám. 8, figs.30–39, fig. 3 del texto; Reinecke, Moths, Grant y Breitkreutz, 2005, p. 36, lám. 21, 22; Cappetta y Cavallo, 2006, lám. 2, fig. 1.
Descripción. El ejemplar referido es un diente superior, lateral–derecho de tamaño pequeño. Su forma es triangular, comprimido, y con la cúspide inclinada hacia la comisura. Los bordes cortantes son lisos. El ápice de la corona no está conservado. La cara labial de la corona es plana; la cara lingual es convexa. El talón distal no presenta cúspide. La raíz se encuentra un poco deteriorada, es baja con dos lóbulos, su contorno basal es notablemente curvo.
Material. Un diente. PALUSAL, P–C118.
Localidad. El Rompido, Formación Arenas de Huelva, Cuenca del Guadalquivir, España.
Discusión. El ejemplar fue comparado con dientes de las mandíbulas de las especies actuales Alopias superciliosus y Alopias vulpinus.
Según Cappetta (1987), los dientes de A. superciliosus Lowe, 1841 son relativamente grandes, hasta 1.5 centímetros de altura, con una cúspide alargada y delgada en la línea anterior e inclinada hacia la comisura en los dientes laterales. Los dientes de A. vulpinus Bonnaterre, 1788, tienen la cúspide baja, más ancha, con la cara labial plana. A la raíz le falta el canal medio. Por otra parte A. pelagicus Nakamura, 1935 presenta dientes con cúspides pequeñas e inclinadas hacia la comisura, estos dientes se distinguen por presentar pequeñas cúspides en el talón distal de los dientes laterales.
Se comparó además con las figuras de Alopias cf. vulpinus de Antunes y Jonet (1970, lám. VII, fig. 26; lám. VIII, figs. 30–39; fig. 3 del texto) y de Cappetta y Cavallo (2006, lám. 2, fig. 1).
El diente estudiado posee mucha semejanza con Alopias cf. vulpinus por ser un diente medianamente pequeño, de cúspide baja y ancha, de bordes cortantes lisos. La cara labial es plana; presenta ausencia del canal medio y sin cúspides en el talón distal de la corona.
Familia Lamnidae Müller y Henle, 1838
Género Isurus Rafinesque, 1810
Isurus desori (Sismonda, 1849) (Figuras 5a–5b)
Oxyrhina xiphodon Agassiz. Rocabert, 1934 p. 82, lám 2, figs. 12–13.
Isurus (Oxyrhina) desori Sismonda. Zbyszewski y Almeida, 1950, p. 329–330, lám.1, figs. 31–33).
Isurus cf. oxyrhinchus Rafinesque. Antunes y Jonet, 1970, p. 137–139, figs. 14–18.
Isurus desori Agassiz. Cappetta, 1987, p. 96; Nolf, 1988, p. 152; lám. 48, figs. 1–10; Mendiola y Martínez, 2003, p. 34, lám. 1, figs. 12–13.
Isurus desori Sismonda. Balbino, 1995, p. 55, lám. 6, fig. 2–4; Mendiola y López, 2005, p. 18–20, lám. 2, figs. 12–14b.
Descripción. Un diente superior, izquierdo, segunda fila. El esmalte ha sido afectado por una corrosión química. La corona es alta y esbelta, volcada hacia la comisura. La cúspide es afilada y de perfil ligeramente sigmoidal. La cara labial es plana. Se percibe el filo de corte hasta la base de la corona, a pesar de la corrosión. La raíz está regularmente conservada, presenta dos lóbulos bifurcados, siendo el lóbulo mesial más largo. La protuberancia lingual es abultada. No presenta dentículos laterales.
Material. Un diente. PALUSAL, P–C71.
Localidad. Bonares–Casa del Pino, Formación Arenas de Huelva, Cuenca del Guadalquivir, España.
Discusión. Los dientes de Isurus desori son difíciles de distinguir de I. oxyrhinchus. En I. oxyrinchus la raíz está menos desarrollada en relación con la corona y en I. desori la raíz es más potente. Los dientes anteriores superiores de I. desori se diferencian de los correspondientes de jóvenes I. hastalis en que los primeros presentan una corona sigmoidal (Arambourg, 1927 in Mendiola y López, 2005). El diente PALUSAL , P–C71, es similar a aquellos ilustrados en la figura 14 del trabajo de Antunes y Jonet (1970), mismo que es identificado como I. cf. oxyrhinchus del tipo "desori".
El ejemplar se comparó con material fósil y se determinó, por la similitud de la corona con perfil ligeramente sigmoidal y por la semejanza de la potencia de la raíz con dientes determinados por Balbino (1995), como I. desori de las localidades de Esbarrondadoiro y Vale de Zebro del Mioceno terminal de la Cuenca de Alvalade, Portugal.
Isurus escheri (Agassiz, 1843) (Figuras 5c–5d)
Especie tipo. Isurus oxyrinchus Rafinesque, 1810.
Isurus cf. oxyrhinchus var. escheri Rafinesque. Antunes y Jonet, 1970, p. 140–143, lám. 4, fig. 21.
Isurus escheri Agassiz. Cappetta, 1987, p. 96.
Descripción. Un diente inferior derecho, tercera fila. El diente se encuentra bien conservado, con una corona alta, esbelta y ligeramente volcada hacia la comisura. El ápice de la misma se proyecta ligeramente en dirección a la cara labial. La cara lingual de la corona es convexa, apreciándose algunas estrías cercanas a la base de la corona. La cara labial es plana. Se observa una tenue crenulación en los bordes de la corona. No presenta dentículos laterales. La raíz es abultada, con dos lóbulos bifurcados. La protuberancia lingual de la raíz está presente y el canal nutricio aparece en el centro de la protuberancia.
Material: Un diente. PALUSAL, P–C1.
Localidad: Bonares–Ambulatorio, Formación Arenas de Huelva, Cuenca del Guadalquivir, España.
Discusión. Este diente se ha comparado con la lámina VI, fig. 21 del trabajo de Antunes y Jonet (1970), donde se ilustra uno de la especie Isurus hastalis variación escheri. Además se comparó con dientes de una mandíbula actual de I. oxyrinchus.
Según Cappetta (1987), la especie I. escheri se caracteriza por mostrar crenulación en los bordes de corte; por lo tanto, el ejemplar aquí referido es determinado como I. escheri por la tenue crenulación que posee su corona en los bordes de corte, esta crenulación es una característica importante destacada por Antunes y Jonet (1970) y Cappetta (1987).
Isurus hastalis (Agassiz, 1843) (Figuras 5e–f y 6a)
Especie tipo. Isurus oxyrinchus Rafinesque, 1810.
Oxyrhina desori Agassiz. Rocabert, 1934, p. 83, lám. 2, fig. 16.
Isurus cf. oxyrhinchus Agassiz. Antunes y Jonet, 1970, p. 137, lám. 6, figs. 14–21; lám. 7, figs. 22 y 24.
Isurus hastalis Agassiz. Cappetta, 1970, p. 18, lám. 5, figs. 1–13; Cappetta, 1987, p. 96; Nolf, 1988, p. 166, lám. 55, figs. 1–7; Balbino, 1995, p. 56, lám. 6, figs. 5–6; lám. 7, fig. 1; Marín, 1999, p. 19, lám. 1, fig. 11; Mendiola y Martínez, 2003, p. 31, lám. I, figs. 4–11, 14–16 y lám. 4, figs. 20–21.
Oxyrhina hastalis Agassiz. Bauzá Rullán y Plans, 1973, p. 80, lám. 3, figs. 20–21; Obrador y Mercadal, 1973, p. 116, 119, fig. 3, n° 3–4; Ruiz et al., 1996, p. 135, lám. 1, fig. 5.
Isurus oxyrhinchus Rafinesque. Mañé, Magrans y Ferrer, 1996, p. 25, lám. 2, fig. 18.
Descripción. Un diente superior derecho, correspondiente a la sexta fila, se encuentra bien conservado, con un esmalte liso y una base de la corona ancha y de forma triangular. La cara lingual de la corona es ligeramente convexa. La cara labial es plana, con un ápice proyectado ligeramente en dirección a la cara labial. El filo de corte de los bordes de la corona alcanza hasta la base de la corona. La raíz es abultada, con los lóbulos grandes prácticamente unidos. La protuberancia lingual está poco marcada.
El otro diente es inferior anterior, y se encuentra en general bien conservado. La corona es alta, de esmalte liso. La base de la corona es ancha y de forma triangular. La cara lingual de la corona es ligeramente convexa y la cara labial es plana. Los bordes de la corona son lisos. La raíz es abultada, con dos lóbulos grandes y prácticamente unidos. La protuberancia lingual está poco marcada. En su centro se puede apreciar el canal nutricio. Ninguno de los dientes presenta dentículos laterales.
Material. Dos dientes. PALUSAL, P–C62–3 y P–C62–10.
Localidad. Bonares–Casa del Pino, Formación Arenas de Huelva, Cuenca del Guadalquivir, España.
Discusión. Los ejemplares fueron comparados con dientes fósiles de Isurus hastalis de la localidad de Santa Margarida, en el Mioceno terminal de la Cuenca de Alvalade, Portugal. La determinación de los dientes fósiles como pertenecientes a la especie Isurus hastalis se debe a la presencia de una corona triangular, de base ancha y bordes lisos. Al mismo tiempo, esta especie se caracteriza por la raíz abultada, con dos lóbulos grandes prácticamente unidos. La ubicación sugerida de los dientes fósiles arriba descritos es apoyada por la comparación de su forma con la de aquellos observados en la mandíbula de ejemplares recientes de I. oxyrinchus.
Isurus sp. (Figuras 6b–c)
Descripción. Un diente inferior, lateral derecho, cuarta fila. Es un diente mal conservado, ha perdido el esmalte y la mayor parte de la raíz. La corona es de forma triangular, la cara lingual de la corona es ligeramente convexa; la cara labial es plana. El filo de corte de los bordes se ha perdido. No presenta dentículos laterales.
Material. Un diente. PALUSAL, P–C107.
Localidad: El Rompido, Formación Arenas de Huelva, Cuenca del Guadalquivir, España.
Discusión. El ejemplar fue comparado con dientes de Isurus hastalis de la localidad de Santa Margarida, del Mioceno terminal de la Cuenca de Alvalade (Portugal). Se determinó como Isurus sp. debido a que con el mal estado del diente, el avanzado deterioro del esmalte y de la raíz, se han perdido características importantes para una determinación más precisa.
Familia Otodontidae Glückman, 1964
Género Carcharocles Jordan y Hannibal, 1923
Carcharocles megalodon (Agassiz, 1843) (Figura 6d)
Carcharodon megalodon Agassiz. Zbyszewski y Almeida, 1950, p. 322, lám. 4, figs. 60 a 69, lám. 5, figs. 72, 76–78.
Procarcharodon megalodon Agassiz. Antunes y Jonet, 1970, p. 143; Cappetta, 1970, p. 26, lám. 6, fig. 2; Antunes, 1978, p. 64, lám. 1, figs. 12–13.
Carcharocles megalodon Agassiz. Bauzá Rullán y Plans, 1973, p. 82, lám. 2, figs. 10–15;
Obrador y Mercadal, 1973, p. 117–119, fig. 3, n° 7; Cappetta, 1987, p. 103, fig. 91; Nolf, 1988, p. 38 lista; lám. 56, figs. 1–6; Balbino, 1995, p. 59, lám. 7, fig. 2; Balbino, 1996, p. 143; Antunes, Balbino y Cappetta, 1999, p. 116; Ward y Galea, 2001, p. 132 lista; Cocke, 2002, p. 14; Mendiola y Martinez, 2003, p. 38, lám. 3, figs. 1–3; Mendiola y López, 2005, p. 22–25, lám. 3, figs. 10–11b.
Descripción. El diente encontrado es relativamente grande y aparece mutilado, conservándose la mitad de la corona y la raíz. La corona es alta con crenulación en el borde. No presenta dentículos laterales. La raíz es prominente, con la protuberancia lingual poco desarrollada.
Material. Un diente. PALUSAL, P–C62–4.
Localidad. Bonares–Casa del Pino, Formación Arenas de Huelva, Cuenca del Guadalquivir, España.
Discusión. El ejemplar se comparó con un diente determinado por Balbino (1995) como C. megalodon perteneciente a Vale de Zebro, Cuenca de Alvalade (Portugal), así como con la figura 91–A de Cappetta (1987), correspondiente a C. megalodon. Los dientes de Carcharocles megalodon son relativamente fáciles de determinar por su gran tamaño. Debido a la corona alta, la crenulación del borde y la prominente raíz se ha podido determinar como C. megalodon.
Género Parotodus Cappetta, 1980
Parotodus benedeni (Le Hon, 1871) (Figura 6e)
Oxyrhina benedeni Le Hon. Leriche, 1926, p. 397–398, figs. 179–181.
Isurus benedeni Le Hon. Antunes, 1978, p. 66, lám. 2, figs, 12–13.
Parotodus benedeni Le Hon. Cappetta, 1987, p. 104–105, fig. 92, E–H; Ward y Galea, 2001, p. 132; Cocke, 2002, p. 33.
Descripción. Un diente superior, derecho. En general se encuentra bien conservado. La cúspide de la corona se encuentra volcada hacia la comisura, el borde de la arista mesial es claramente convexo; el borde de la arista distal es cóncavo. La cara lingual es de forma convexa, en esta cara se puede apreciar un ancho collar. La cara labial de la corona es plana. La textura de la corona es lisa. La raíz es alta y voluminosa, con dos lóbulos largos. La protuberancia lingual está presente, si bien el canal nutricio está ausente.
Material. Un diente. PALUSAL, P–C62–5.
Localidad. Bonares–Casa del Pino, Formación Arenas de Huelva, Cuenca del Guadalquivir, España.
Discusión. Se comparó con las figuras 179–181 del trabajo de Leriche (1926), determinadas como Oxyrhina benedeni, y con la lámina 2, fig. 12, de Antunes (1978), determinada como Isurus benedeni y procedente del Plioceno de Angola.
Cappetta (1987; p. 97) realizó una aclaración sobre algunas especies previamente asignadas como Isurus, asignándolas a otros géneros, como Cretoxyrhina (C. mantelli), Anomotodon (A. sheppeyensis) y Parotodus (P. benedeni).
El diente que se presenta en este trabajo es muy similar al de la fig. 92–H del trabajo de Cappetta (1987), que corresponde a un diente superior lateral, con la corona volcada hacia la comisura y la raíz alta y voluminosa con dos lóbulos largos.
Ward y Galea (2001) realizaron una revisión de los seláceos del Mioceno de Malta y revisan la sistemática de esta especie, asignándole el nombre de Parotodus benedeni.
DISCUSIÓN
Los taxones de Lamniformes encontrados en la Formación Arenas de Huelva, se han registrado también en diferentes puntos del Neógeno de Europa. La especie Carcharias taurus inicio su registro fósil a partir del Plioceno. Se ha encontrado en el Plioceno de Italia (Cappetta y Cavallo, 2006), Francia (Cappetta y Nolf, 1991), España (Mañé et al., 1996; Mas, 2000) y Angola (Antunes, 1978). Se distribuye actualmente en aguas cálidas y tropicales del Atlántico, Mediterráneo e Indo–Oeste del Pacifico; no se incluye el centro y el este del Pacífico; su distribución es costera, generalmente se encuentra entre 15–25 metros de profundidad (Compagno et al., 2005).
La especie Alopias aff. vulpinus se ha registrado en el Oligoceno de Alemania (Reinecke et al., 2005), Mioceno de Lisboa (Antunes y Jonet, 1970) y en el Plioceno de Italia (Cappetta y Cavallo, 2006), esta especie es cosmopolita en la actualidad y se distribuyen en costas y océanos del mundo, desde aguas tropicales, hasta frías.
Isurus desori se ha registrado en el Mioceno de Italia, Cabinda, Zaire y Francia (Cappetta, 1987), en el Mioceno de Portugal (Balbino, 1995; Antunes y Jonet, 1970), Mioceno y Plioceno de España (Mendiola y Martínez, 2003). Isurus escheri, se ha registrado en el Mioceno de Suiza; Terciario de Alemania; Mioceno superior de Bélgica (Cappetta, 1987) y en el Mioceno de Lisboa (Antunes y Jonet, 1970). Isurus hastalis tiene una amplia distribución en el Mioceno de Europa, Norte y Sur América, en África y Australia. Esta especie alcanza por lo menos hasta el Plioceno medio (Cappetta, 1987). En Portugal se ha registrado en el Mioceno de Portugal (Balbino, 1995; Antunes y Jonet, 1970). En España se ha registrado en el Mioceno y Plioceno de Cataluña; Mioceno y Plioceno medio–superior de las Islas Baleares; Mioceno y Plioceno de la Comunidad Valenciana y Andalucía (Mendiola y Martínez, 2003). La especie tipo para tiburones fósiles del género Isurus, es la especie actual Isurus oxyrinchus (Rafinesque, 1810), esta es una especie cosmopolita, que se distribuye en mares tropicales y templados. Puede encontrarse en la costa o en el océano, entre 0–500 metros de profundidad (Compagno et al., 2005).
Según Cappetta (1987), durante el Mioceno y Plioceno se ha registrado Carcharocles megalodon en el Mioceno y Plioceno de Norte América, Europa, Australia, Nueva Zelanda, Japón, India, Oeste de India, en la parte occidental de África y en el Norte de África. Además se ha registrado esta especie en el Mioceno de Portugal (Balbino, 1995; Balbino, 1996; Antunes et al., 1999; Antunes y Jonet, 1970) y en el Plioceno de Angola (Antunes, 1978). En España se ha registrado en el Mioceno de Cataluña, Islas Baleraes, Valenciana, Murcia y en el Mioceno y Plioceno de Andalucía (Mendiola y Martínez 2003). Según Laurito–Mora (1999) Carcharocles megalodon fue una especie de distribución cosmopolita, dado que aparece prácticamente en todos los depósitos sedimentarios del Cenozoico superior que contienen dientes de tiburón alrededor del planeta; eran probablemente potentes tiburones nadadores nectónicos y epipelágicos, que habitaban tanto las plataformas continentales e insulares como las regiones oceánicas y quizás incluso frecuentaban la zona mesopelágica. Habría sido el depredador que probablemente ocupaba la cima de la cadena alimentaria de los océanos del mundo durante el Cenozoico superior.
Según Cappetta (1987) el género Parotodus está presente en el Oligoceno inferior de Bélgica, pero es especialmente frecuente en el Mioceno de Italia, Malta, Bélgica y Suiza. Se ha encontrado además en el Mioceno de Portugal (Zbyszewski y Almeida, 1950), en España en el Mioceno de las Islas Baleares y la Comunidad Valenciada (Mendiola y Martínez, 2003 y Mas, 2003). Su registro ha persistido hasta el Plioceno de Angola (Antunes, 1978). El género Parotodus no es muy abundante, es probable que hayan tenido vida pelágica, observándose excepcionalmente en los yacimientos neríticos ricos en fósiles (Cappetta, 1987).
CONCLUSIONES
El orden Lamniformes se encuentra representado en los yacimientos del Plioceno inferior de la Formación Arenas de Huelva, Cuenca del Guadalquivir, España, por los siguientes taxones (Tabla 1):
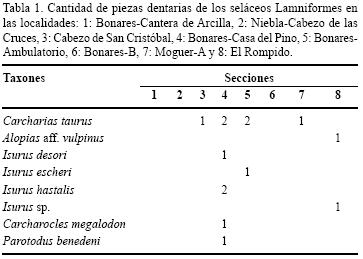
Las especies Carcharias taurus y Alopias vulpinus viven aún en la actualidad y se han registrado en la Formación Arenas de Huelva.
Los taxones de Lamniformes encontrados eran de vida nectónica, en general frecuentaban la zona litoral y nerítica, sin embargo, algunos podrían llegar a tener incidencia en la zona pelágica. Se destaca la mayor frecuencia de dientes de Carcharias taurus, especie típica de distribución litoral–nerítica y de regiones tropicales y subtropicales. El estudio de las asociaciones de seláceos indica un medio de aguas subtropicales a templadas.
Se registran por primera vez para el Neógeno de España: Alopias aff. vulpinus e Isurus escheri.
La distribución bioestratigráfica de los diferentes taxones de Lamniformes encontrados tiene una mayor frecuencia en yacimientos Neógenos. Es notable destacar la presencia de Carcharias taurus, que inició su registro fósil a partir del Plioceno. Por lo tanto, la fauna encontrada en la Formación Arenas de Huelva es compatible con una edad Plioceno s.l.
AGRADECIMIENTOS
Se agradece a los dos dictaminadores que hicieron la revisión del trabajo.
REFERENCIAS
Agassiz, L., 1843, Recherches sur les Poissons fossiles: Neuchâtel and Soleure, 3(7), 32 pp. [ Links ]
Andrés, I., 1982, Estudio malacológico (Clase Bivalvia) del Plioceno marino de Bonares: Salamanca, España, Universidad de Salamanca, tesis doctoral, 410 pp. [ Links ]
Antunes, M.T., 1978, Faunes ichthyologiques du Nèogène supérieur d' Angola, Leur àge, remarques sur le Pliocène Marin en Afrique australe: Ciências da Terra (UNL), 4, 59–90. [ Links ]
Antunes, M.T., Jonet, S., 1970, Requins de L' Helvéntien supérieur et du Tortonien de Lisbonne: Revista da Facultade de Ciências de Lisboa, 16(1), 119–280. [ Links ]
Antunes, M.T., Balbino, A., Cappetta, H., 1999, Sélaciens du Miocène terminal du bassin d'Alvalade (Portugal) Essai de synthèse: Ciências da Terra (UNL), 13, 115–129. [ Links ]
Arambourg, C.,1927, Les poissons fossiles d'Oran: Algeria, Bastide–Jourdau, Matériaux puur la Carte géolgique de l'Alger, 1re sér., Paléontologie 6, 1–218. [ Links ]
Balbino, A.C., 1995, Seláceos (pisces) do Miocénico terminal da Bacia de Alvalade (Portugal) Sistemática, Ecologia, Paleoambientes, Comparação com faunas actuais: Évora, Porgual, Universidade de Évora, tesis doctoral, 188 pp. [ Links ]
Balbino, A.C., 1996, Sharks from the Middle and early Upper Miocene from Lisbon, Portugal A check–list: Comunicações do Instituto Geológico e Mineiro, 82, 141–144. [ Links ]
Bauzá–Rullán, J., Plans, J., 1973, Contribución al conocimiento de la fauna ictiológica del Neógeno Catalano Balear: Boletín de la Sociedad de Historia Natural de Baleares, 18, 72–131. [ Links ]
Berg, L.S., 1958, System der regenten und fossilen Fischartigen und Fische: Berlin, Deutsch Verlag Wiss, 310 pp. [ Links ]
Bonaparte, C.L.J.L., 1838, Synopsis vertebratorum systematis: Nuon Annali delle Scienze Naturali, 2, 105–33. [ Links ]
Bonnaterre, J.P., 1788, Tableau encyclopédique et méthodique des trois règnes de la nature. Ichthyologie: Paris, Chez Panckoucke, 215 pp. [ Links ]
Cappetta, H., 1970, Les Sélaciens du Miocène de la région de Montpellier: Paleovertebrata, Mémoire extraordinaire, 139 pp. [ Links ]
Cappetta, H., 1980, Modification du statut générique de quelques espèces de sèlaciens crètacès et tertiaires: Paleovertebrata, 10(1), 29–42. [ Links ]
Cappetta, H., 1987, Chondrichthyes II , Mesozoic and Cenozoic Elasmobranchii, in Schultze H.–P. (ed.), Handbook of Paleoichthyology: Stuggart–New York, Verlag Dr. Gustav Fischer, 3B, 193 pp. [ Links ]
Cappetta, H., 2006, Elasmobranchii Post–Triadici (Index specierum et generum), in Riegraf, W. (ed.), Fossilium Catalogus I, Animalia pars 142: Leiden, Backhuys Publish, 472 pp. [ Links ]
Cappetta, H., Cavallo, O., 2006, Les Sélaciens du Pliocène de la Region d' Alba (Piémont, Italie Nord–Ouest): Rivista Piemontese di Storia Naturale, 27, 37–76. [ Links ]
Cappetta, H., Nolf, D., 1991, Les Sélaciens du Pliocène inférieur de Le Puget sur Argens (Sud–Est de la France): Palaeontographica A, 218, 49–67. [ Links ]
Civis, J., Sierro, J.F., González–Delgado, J.A., Flores, J.A., Andrés, I., Porta, J., Valle, M.F., 1987, El Neógeno marino de la provincia de Huelva, antecedentes y definición de las unidades litoestratigráficas, in Civis, J. (ed.), Paleontología del Neógeno de Huelva (W del Guadalquivir): Salamanca, Ediciones Universidad de Salamanca, 9–27. [ Links ]
Cocke, J., 2002, Fossil Shark Teeth of the World, United States of America, A collector's guide: Torrance, CA, Lamna books, 170 pp. [ Links ]
Compagno, J.L.V., 1973, Interrelationships of living elasmobranchs, in Greenwood, P.H., Miles, R.S., Patterson, C. (eds.), Interrelationships of fishes: Zoological Journal of the Linnean Society, 53, suppl. 1, 1–37. [ Links ]
Compagno, J.L.V., 1977, Phyletic relationsphips of living sharks and rays: American Zoologist, 17(2), 302–22. [ Links ]
Compagno, L.J.V., 1984a, Sharks of the World. An annotated and illustrated catalogue of shark species known to date; Part 1, Hexanchiformes to Lamniformes: Rome, United Nations Development Programme, FAO fisheries synopsis 125, FAO species catalogue, 4(1), 249 pp. [ Links ]
Compagno, L.J.V., 1984b, Sharks of the World. An annotated and illustrated catalogue of shark species known to date; Part 2, Carcharhiniformes: Rome, United Nations Development Programme, FAO fisheries synopsis 125, FAO species catalogue, 4(2), 251–655. [ Links ]
Compagno, L., 2005, Checklist of living Chondrichthyes, in Hamlett, W.C. (ed.), Reproductive biology and phylogeny of Chondrichthyes: sharks, batoids and chimaeras: Enfield, Plymouth, Science Publishers, 503–548. [ Links ]
Compagno, L., Dando, M., Fowler, S., 2005, Sharks of the World Princeton Field Guides: United States of America, Princeton University Press, 368 pp. [ Links ]
Dabrio, C.J., González–Delgado, J.A., Civis, J., Sierro, F.J., 1988, Influencia de las tempestades en la generación de niveles e interniveles fosilíferos en las Arenas de Huelva (Plioceno), en IV Jornadas de Paleontología: Salamanca, España, Comunicaciones, 34–36. [ Links ]
Flores, J.A., 1985, Nanoplancton calcáreo en el Neógeno del borde noroccidental de la Cuenca del Guadalquivir (SO de España): Salamanca, España, Universidad de Salamanca, tesis doctoral, 714 pp. [ Links ]
García, E.X.M., 2008, Condrictios y Osteíctios del Neógeno de Huelva (Formación "Arenas de Huelva"): Salamanca, España, Universidad de Salamanca, tesis doctoral, 313 pp. [ Links ]
Glückman, L.S., 1964, Sharks of Paleogene and their stratigraphic significance: Moscow–Leningrad, Nauka Press, 229 pp. [ Links ]
González–Delgado, J.A., 1983, Estudio de los Gasterópodos del Plioceno de Huelva, Salamanca: España, Universidad de Salamanca, tesis doctoral, 474 pp. [ Links ]
González–Delgado, J.A., Civis, J., Dabrio, C.J., Goy, J.L., Ledesma, S., Pais, J., Sierro, F.J., Zazo, C., 2004, Cuenca del Guadalquivir, en Vera, J.A. (ed.), Geología de España: Madrid, Sociedad Geológica Española, Instituto Geológico Minero Español, 543–550. [ Links ]
González–Regalado, M.L., 1986, Las asociaciones de foraminíferos bentónicos en las arenas fosilíferas del Plioceno de Huelva: Sevilla, España, Universidad de Sevilla, tesis doctoral, 314 pp. [ Links ]
Hay, O.P., 1902, Bibliograpy and catalogue of the fossil vertebrata of North America: Washington, DC, Bulletin of the United States Geological Survey, 179, 868 pp. [ Links ]
Huxley, T.H., 1880, On the applications of the laws of evolution to the arrangement of the vertebrata and more particularly of the Mammalia: Proceedings of the Zoological Society of London, 43, 649–661. [ Links ]
Jordan, D.S., Hannibal, H., 1923, Fossil sharks and rays of the Pacific slope of North America: Bulletin of the Southern California Academy of Sciences, 22, 27–68. [ Links ]
Laurito–Mora, C.A., 1999, Los seláceos fósiles de la localidad de Alto Guayacán (y otros ictiolitos asociados) Mioceno superior–Plioceno inferior de la Formación Uscari, Provincia de Limón, Costa Rica: San José, Costa Rica, Texto Comunicación S.A, 125 pp. [ Links ]
Le Hon, H.S., 1871, Préliminaires d'un mémoir sur les poissons tertiaires de Belgique: Bruxelles, H. Merzbach, 15 pp. [ Links ]
Leriche, M., 1926, Les Poissons Neògenés de la Belgique: Institut Royal des Sciences Naturelles de Belgique, 32, 397–398. [ Links ]
Lowe, R.T., 1841, On new species of fishes from Madeira: Proceedings of the General Meetings for Scientific Business of the Zoological Society of London, 8, 36–39. [ Links ]
Mañé, R., Magrans, J., Ferrer, E., 1996, Ictiologia fòssil del Pliocè del Baix Llobregat. II. Selacis pleurotremats: Batalleria, 6, 19–33. [ Links ]
Marín, J.F., 1999, Paleoictiología de algunos yacimientos neógenos de la provincia de Alicante: Revista Ilicitana de Paleontología y Mineralogía, 1(1), 4–26. [ Links ]
Mas, G., 2000, Ictiofauna del Pliocè mitjà–superior de la conca sedimentària de Palma lles Balears, Mediterrània Occidental, Implicaciones paleoambientals: Bolletin de la Societat d'Història Natural de les Baleares, 43, 39–61. [ Links ]
Mas, G., 2003, Presència de Parotodus benedeni (Le Hon, 1871) (Pisces: Chondrichthyes: Otodontidae) al Enojen de Marlloca i menoría (Illes Balears, Mediterrània occidental) Consideracions taxonòmiques i paleoambientals: Bolletin de la Societat d'Història Natural de les Baleares, 46, 85–90. [ Links ]
Mayoral, E., Pendón J.G., 1987, Icnofacies y sedimentación en zona costera, Plioceno superior?, litoral de Huelva: Acta Geológica Hispánica, 21–22, 507–513. [ Links ]
Mendiola, C., López., 2005, La ictiofauna fósil (Chondrichthyes, Euselachii) del Serravaliense de Alicante (Sureste de España): Revista de la Societat Paleontològica d'Elx, sección Paleontológica, 14, 1–51. [ Links ]
Mendiola, C., Martínez, J., 2003, La ictiofauna fósil (Chondrichthyes, Euselachii) del Mesozoico y Cenozoico de España: Revista de la Societat Paleontològica d'Elx, sección. Paleontológica, 9, 1–103. [ Links ]
Müller, J., Henle, F.G.J., 1838, On the generic characters of the cartilaginous fishes, with descriptions of the new genera: Magazine of Natural History, 2, 36. [ Links ]
Müller, J., Henle, F.G.J., 1839, Systematische Beschreibung der Plagiostomen: Berlin, Veit, 2, 73. [ Links ]
Nakamura, H., 1935, On the two species of the thresher shark from Formosan water: Memoirs of the Faculty of Science and Agriculture, Taihoku Imperial University, 14(1), 1–6. [ Links ]
Nolf, D., 1988, Fossiles de Belgique. Dents de requins et raies du tertiaire de la Belgique: Belgique, l'Institut Royal des Sciences Naturalles de Belgique, 184 pp. [ Links ]
Obrador, A., Mercadal, B., 1973, Nuevas localidades con fauna ictiológica para el Neógeno menorquín: Acta Geológica Hispánica, 8(4), 115–119. [ Links ]
Rafinesque, C.S., 1810, Caratteri di alcuni nuovi generi e nuove specie di animali e piante della Sicilia: Palermo, Italia, Sanfilippo, 105 pp. [ Links ]
Reinecke, T., Moths, H., Grant, A., Breitkreutz H., 2005, Die Elasmobranchier des Norddeutschen Chattiums, Insbesondere des Sternberger Gesteins (Eochattium, Oberes Oligozän): Paleontos, 8, 134 pp. [ Links ]
Rocabert, L., 1934, Contribució al coneixement de la fauna icitiològica terciària catalana: Butlletí de la Institució Catalana d' Història Natural, 34, 78–106. [ Links ]
Ruiz, M., Clauss, F., González–Regalado, M.L., 1996, Primeras consideraciones sobre los Condrichtios de la Formación "Arenas de Huelva": Stvdia Geológica Salmanticensia, 32, 129–139. [ Links ]
Sierro, F.J., 1984, Foraminíferos planctónicos y Bioestratigrafía del Mioceno Superior–Plioceno del borde occidental de la Cuenca del Guadalquivir (S.O. de España): España, Universidad de Salamanca, tesis doctoral, 391 pp. [ Links ]
Sismonda, E., 1849, Descrizione dei pesci e dei crustacei fossili nel Piemonte: Memoire della Reale Academia delle Scienze di Torino, 10. [ Links ]
Ward, D., Galea, G., 2001, Additions to, and a review of, the Miocene shark and ray fauna of Malta: The Central Mediterranean Naturalist, 3(3), 131–146. [ Links ]
Zbyszewki, G., Almeida, F., 1950, Os peixes miocénicos portugueses: Comunicações dos Servicos Geológicos de Portugal, 31, 311–412. [ Links ]

