










Servicios Personalizados
Revista
Articulo
 Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista mexicana de ciencias geológicas
versión On-line ISSN 2007-2902versión impresa ISSN 1026-8774
Rev. mex. cienc. geol vol.26 no.1 Ciudad de México abr. 2009
Upper lower Eocene calcareous nannoplankton from the Las Pocitas core (Tepetate formation), Baja California Sur, Mexico
Nanoplancton calcáreo de la parte superior del Eoceno inferior en el núcleo Las Pocitas (formación Tepetate), Baja California Sur, México
Erandi García–Cordero and Ana Luisa Carreño*
Instituto de Geología, Universidad Nacional Autónoma de México, Circuito Exterior, Ciudad Universitaria, Delegación Coyoacán, 04510 Mexico D.F., Mexico. *anacar@geol–sun.igeolcu.unam.mx
Manuscript received: September 15, 2007
Corrected manuscript received: September 12, 2008
Manuscript accepted: October 9, 2008
ABSTRACT
Sixty species of calcareous nannoplankton from the Tepetate formation were recovered from a drill hole near Las Pocitas, Baja California Sur. In spite of its low abundance and erratic distribution throughout the sedimentary column, the great amount of reworked species, as well as the absence of traditional index fossils representing the standard tropical biozonation, the co–occurrence of Reticulofenestra dyctioda, Rhabdosphaera crebra, R. pinguis, Micrantholithus flos, Pontosphaera pectinata and Lanternithus minutus situates the studied stratigraphic column at the Discoasteroides kuepperi CP12a Subzone of Okada and Bukry, dated between 49.5 and 49 Ma at the top of the upper lower Eocene.
The population structure as well as the lithological features of the sedimentary package, suggest a temperate open–sea deposit, which confirms interpretations based on other marine fossils at arroyo Datilar, El Conejo, Salada, Colorado y Las Pocitas, where rocks of the lower middle Eocene Tepetate formation crop out.
Key words: biostratigraphy, calcareous nannofossils, Tepetate formation, Eocene, Baja California Sur, Mexico.
RESUMEN
Sesenta especies de nanoplancton calcáreo provenientes de la formación Tepetate fueron recuperadas de un pozo exploratorio perforado en los alrededores de Las Pocitas, Baja California Sur. A pesar de su baja abundancia y distribución errática a través de la columna sedimentaria, la alta frecuencia de especies retrabajadas así como la ausencia de fósiles índice tradicionales de las biozonaciones tropicales estándares, la presencia de Reticulofenestra dyctioda, Rhabdosphaera crebra, R. pinguis, Micrantholithus flos, Pontosphaera pectinata y Lanternithus minutus sitúa la columna estratigráfica estudiada dentro de la subzona CP12a de Discoateroides kuepperi de Okada y Bukry calibrada entre 49.5 y 49 Ma en la parte final del Eoceno inferior.
La estructura de la población, así como las características litológicas de los paquetes sedimentarios sugieren condiciones de depósito de mar abierto en un ambiente templado, confirmando interpretaciones previas basadas en diversos fósiles marinos provenientes de los arroyos Datilar, El Conejo, Salada, Colorado y Las Pocitas, donde afloran rocas del Eoceno inferior de la formación Tepetate.
Palabras clave: bioestratigrafía, nanofósiles calcáreos, formación Tepetate, Eoceno, Baja California Sur, México.
INTRODUCTION
While worldwide the calcareous nannoplakton is a valuable tool in biostratigraphy as well as in other Earth Sciences disciplines, in Mexico it is still poorly known, except for the research made in petroleum exploration, where results remain unpublished as private reports. Several undergraduate theses have been conducted in Mexico but unfortunately, all those remain unpublished.
In order to test the use of standard nannoplankton zonation in temperate areas in Mexico and its application to biostratigraphic studies, as well as to evaluate the resolution of this important group in comparison with other microfossils, the present paper evaluates the distribution of this flora throughout the upper part of the Tepetate formation and, on the basis of its assemblages, an age is proposed. The study forms part of a project consisting in the stratigraphic revision of the Cenozoic marine units that crop out in the Baja California Peninsula, which pretends to obtain enough data in order to best asses the geological evolution of the Baja California Peninsula.
PREVIOUS WORK
The Tepetate formation as an informal lithostratigraphic unit was named by Heim (1922) to include the marine rocks outcropping near rancho El Tepetate in Arroyo Colorado, Baja California Sur, Mexico. It is constituted by slightly folded, well–stratified sandstone and shale, 1,000 m thick, and was interpreted as an Eocene neritic deposit. Beal (1948) assigned a middle Eocene age to this unit concluding that it was deposited in a coastal marine environment. Mina–Uhink (1957), based on the presence of several species of foraminifers, strongly suggested an early Eocene age and considered that the deposits included a coastal to neritic paleoenvironment.
Within a biostratigraphic scope using microfossils, three important studies must be mentioned: the one by Knappe (1974, in Minch and Leslie, 1979), that of Fulwider (1976) using planktonic foraminifers, and Coleman's (1979) with calcareous nannoplankton. In spite of the important contribution of Fulwider's (1976) paper to the lithological and biostratigraphical characterization of the Tepetate formation, the lack of precise location of the composite measured sections, that otherwise are the same used by Coleman (1979) in her study, both master theses remain unpublished.
Ledesma–Vázquez et al. (1999) and Carreño et al. (2000), in a composite measured section at arroyo Colorado and complemented with a section measured at Las Pocitas, assigned a latest early Eocene–earliest middle Eocene age between 51.2 to 48.4 Ma on the basis of planktonic foraminifers. According to sedimentary structures, benthic foraminifers and ostracods association, these authors suggested a deposition from the inner to the outer marine shelf in depths shallower than 150 m. Recently, in the same core here studied, a planktonic foraminifer was recorded by Miranda–Martínez and Carreño (2008) and an early Eocene age between 50.4–49 Ma was assigned to the stratigraphic column. Based on benthic foraminiferal association, a middle upper bathyal deposit was suggested.
STUDY AREA
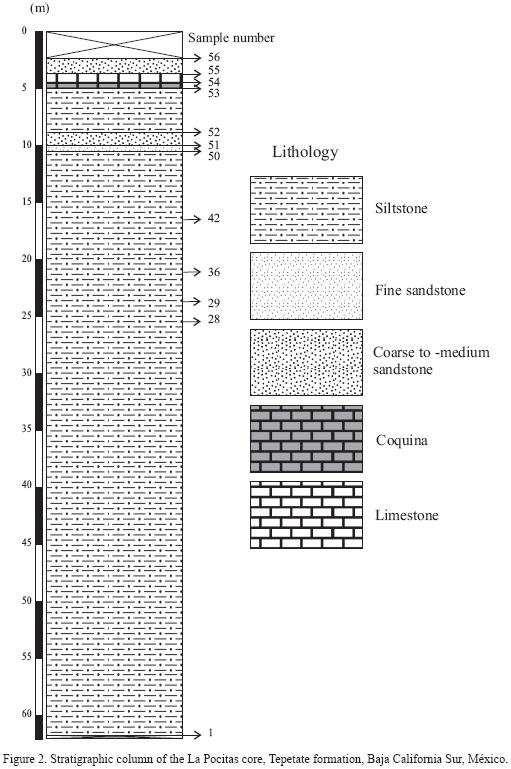
The sedimentary rocks of the Tepetate formation came from a core (Loc. IGM–2984; 24°24'00" N, 111°05'47" W, Las Pocitas Quadrangle, Baja California Sur, Mexico, 1:50,000, G12C79, INEGI, 1983; [Figure 1]), recovered in the surroundings of Las Pocitas town, located approximately 76 km south of Ciudad Constitución, La Paz County, Baja California Sur, east of the Transpeninsular Highway [MEX 1]. The drill hole of 80 m made by Secretaría de Recursos Hidráulicos recovered 62 m of monotonous very light gray, fine–grained sandstone, capped by a limestone (Figure 2), from which 56 samples were obtained.
MATERIAL OF STUDY
For the calcareous nannofossil analyses, the samples were prepared according to the normal procedure (Perch–Nielsen, 1985) including selection, cleaning and pulverization, and subsequent smear–slide preparation. Two smear slides per sample were analyzed and for their observation and fossil identification a polarizing microscope (Olympus BH–2) with crossed nichols and 40X and 100X achromatic objectives DIN standard was used. In some cases, a ½ χ gypsum plate of 30 nm delay of wavelength was used to facilitate the observation of non birefringent nannoliths and discoasterids. The smear–slides were deposited in the Colección Nacional de Paleontología, Museo María del Carmen Perrilliat, Instituto de Geología, Universidad Nacional Autónoma de México with the numbers IGML–1–Mi to IGML–22–Mi, IGML–31–Mi to IGML–106–Mi and IGML–108–Mi to IGML–118–Mi. Thin–sections of samples from the limestone capping the Tepetate formation were prepared, but because no microfossils were found, the thin–sections are not deposited. Micrographs of specimen in polarizing microscope were taken and digital images were also left in the same repository with numbers IGMD–1–Mi to IGMD–60.
MICROPALEONTOLOGY
Taxonomic assignations of the calcareous nannoplankton recovered at the Las Pocitas core are based on Perch–Nielsen (1977, 1985), Bramlette and Sullivan (1961), Bramlette and Wilcoxon (1967), Bukry and Percival (1971), Gartner (1971), Sullivan (1965), and Wise et al. (2002). Sixty species belonging to 26 genera of calcareous nannofossils were identified. Calcareous nannoplankton was absent in the three upper samples of the recovered stratigraphic column, which otherwise might represent a younger unit instead of rocks belonging to the Tepetate formation.
In spite of the relative abundance recorded in several samples of the calcareous nannoplankton, most part of the assemblages correspond to Cretaceous and Paleogene reworked species. In general the nannofossils have moderate to good conservation and no sign of overgrowth is observed. Nevertheless, the absence of many forms, in particular diagnostic age species, suggests strong dissolution. The distribution along the core is not homogeneous for most part of the recorded species. According to ODP Leg 191 Shipboard Scientific Party (2001), the relative abundance is considered as common (one specimen in 2–10 fields of view at 100X immersion) for reworked Mesozoic species as Watznaueria barnesae and W. fossacincta, and Paleocene species as Cruciplacolithus primus, Cr. tenuis, Cyclagelosphaera reinhardtii, Discoaster mohleri, Ericsonia subpertusa, Fasciculithus tympaniformis, Micrantholithus pinguis, Neochiatozygus digitosus and Toweius selandianus.
The species with common frequency throughout the core length were Markalius inversus, Fasciculithus involutus, Coccolithus pelagicus and Sphenolithus radians, whereas species belonging to genera Sphenolithus, S. anarrophus, S. obtusus, S. editus, Rhabdosphaera Haeckel, (R. crebra, R. pinguis), Helicosphaera (H. seminulum, H. lophota) and Pontosphaera (P. pectinata, P. ocellata, P. punctosa), have only one isolated record per sample. From the total species identified, many of them were only recorded once throughout the core: F. tympaniformis, Neochiastozygus digitosus, R. truncata, R. inflata, Transversopontis sigmoidalis, W. fossacincta, Scapholithus rhombiformis, D. mohleri, D. septemradiatus, Cruciplacolithus cruciformis, C. tenius, Pontosphaera versa, Chiasmolithus consuetus, C. bidens, C. solitus, E. subpertusa, Isthmolithus unipons and Micrantholithus pinguis.
Biostratigraphy
Calcareous nannoplankton is scarce and sparse. Also, most of the diagnostic species that could allow the reconnaissance of standard low latitude biozones are diluted or dissolved. Therefore, the stratigraphic range of the species here recorded was examinated (Perch–Nielsen, 1985; Wise et al., 2002) in order to estimate, as far as possible, the span of time covered by the studied sedimentary column.
With the exception of the species that has a restricted range to the Mesozoic and to the Paleocene, and besides the low–ranging Paleogene species, the association represented by Fasciculithus involutus, Sphenolithus editus, Rhabdosphaera truncata, R. crebra, R. inflata, Chiasmolithus solitus, Girsia gammation, Isthmolithus unipons, Discoaster septemradiatus, Toweius callosus, Pontosphaera plana, Helicosphaera lophota, H. seminulum, Reticulofenestra dyctioda, R. pinguis, Micrantholithus flos, Pontosphaera pectinata, and Lanternithus minutus, indicates that the whole sedimentary column belongs to the Eocene (Figure 3).
The erratic distribution of these species in the core (Table 1) does not allow recognition of FO's and LO's and even less of FAD's and LAD's; nevertheless, the co–occurrence of R. dyctioda, R. crebra, R. pinguis, M. flos, P. pectinata and L. minutus situates the studied stratigraphic column in the Discoateroides kuepperi CP12a Subzone of Okada and Bukry (1980), dated between 49.5 and 49 Ma around the boundary between the lower and middle Eocene (Figure 3).
Younger species such as Micrantholithuscf.altus, which has a restricted range in the CP14 Reticulofenestra umbilica Zone of Bukry (1973) and Reticulofenestra scripsae whose FAD occurs within this biozone, strongly suggest a younger age for the sedimentary column; nevertheless, the doubtful assignation of first species and the isolated record at sample 12 of the second species prevent assignment to this biozone.
This age assignment is in agreement with Carreño et al. (2000). Based on planktonic foraminifera, they established, at the Arroyo Colorado locality situated west of the Las Pocitas area, an age between 51.2 and 48.4 Ma, a span of time that includes the latest early Eocene to the early middle Eocene. Planktonic foraminifers recovered from Las Pocitas core at the same levels studied here (Miranda–Martínez and Carreño, 2008), allowed to recognize an assemblage equivalent of Planorotalites palmerae–Acarinina pentacamerata Zone (= P9 Zone of Premoli–Silva et al., 2003), which according to Berggren et al. (1995) situates this part of the unit in the early Eocene, between 50.4 and 49 Ma, age that matches well with that inferred from the calcareous nannoplankton. Diagnostic biostratigraphic species are illustrated in Figures 4, 5 y 6.
Paleoenvironment
The lithology present in the Las Pocitas core, which is formed by sandstone beds composed of fine–to–medium grained sand with siliceous cement, whose grains are composed of angular quartz, feldspars, and lithics, suggests that this part of the sedimentary column belongs to the upper part of the interbedded member of Fulwider (1976). These lithological characteristics, as well as the age assignment, suggest that this sedimentary package corresponds to the upper part of the Tepetate formation.
Following Perch–Nielsen (1985), Sphenolithus species are characteristic species of low latitude, open sea, and warm water, whereas Pontosphaera species are frequent in hemipelagic sediments. Both genera are well represented in the studied sedimentary column. Fulwider (1976) stated that some levels of the interbedded member were deposited under low oxygen conditions, probably in the upper submarine fan or possible canyon mouth deposit. However, throughout the stratigraphic section here studied, opportunistic species associated to stress conditions such as Braarudosphaera bigelowii are scarce; the absence of biogenic structures reinforces the interpretation that this part of the Tepetate formation was deposited in an oceanic paleoenvironment, near the upper slope under normal conditions in temperate water masses. The fact that many of the tropical index species are absent seems to strengthen a less warm or tropical climate.
According to Miranda–Martínez and Carreño (2008), the relative abundance of planktonic foraminifers over the benthonic ones in the Las Pocitas core is almost equal or slightly lower, whereas the high species diversity, as well as its frequency throughout the stratigraphic column suggest upper slope deposits, data that match well with the interpretation on the basis of calcareous nannoplankton. Diagnostic paleoenvironmental species are shown in Figures 4, 5 y 6.
DISCUSSION
The revision of the calcareous nannoplankton data given by Coleman (1979), who uses the biozonation of Bukry (1973) to assign age, suggests that the rocks of the Tepetate formation constitute a continuous deposit that covers a span of time from the Maastrichtian to the lower Eocene. This author mentions no species for the Late Cretaceous, and the age was established by the presence of long ranging calcareous nannoplankton species in rocks outcropping at arroyo Salada. However, for the lower Paleogene, the recognized biozones from the bottom to the top were the Cruciplacolithus tenuis, Fasciculithus tympaniformis, Heliolithus kleinpellii, Discoaster mohlei, Heliolithus riedelii, Discoaster multiradiatus, Discoaster diastypus, Tribrachiatus orthostylus and Discoaster lodoensis.
On the other hand, Fulwider (1976) recorded at the arroyo Salada outcrops the presence of Tetralithus nitidus, Watznaueria barnesae, Watznauria biporta and Arkhangelskiella cymbiformis and, on the basis of this assemblage, assigned a Maastrichtian age to the base of the Tepetate formation. Nevertheless, A. cymbioformis, which otherwise has several definitions attached, in the sense of Bukry (1973) is restricted to the Campanian Eiffelithus augustus Zone, while the other species has a more wide range.
Neither Coleman (1979) nor Fulwider (1976) mentioned the presence of reworked calcareous nannoplankton species throughout the studied lithostratigraphic columns. In the present study, the reworked Cretaceous species are continuously recorded throughout the Las Pocitas core, sometimes in such frequency that they constitute, as was stated earlier, the dominant assemblage in a sample.
Contrary to data presented by Coleman (1979), Fulwider (1976) recognized a hiatus at the base of the Paleogene, evidenced by the lack of at least two foraminiferal biozones, and being the Morozovella trinidadensis Zone the first recognized biozone. The encompassing examination of the species recorded by both authors as well as the updated biozonal schemes, situate the top of the Tepetate formation at around 48.60 ± 0.2 Ma in the late Ypresian (Gradstein et al., 2004), which is close to the age given for the base of the CP12a Zone of Bukry (1973) here assigned.
ACKNOWLEDGMENTS
Authors thank Héctor Ruiz Ruiz from the Laboratorio de Paleontología, Centro Regional de Estudios de Laboratorio, Activo Regional de Exploración, Región Norte, PEMEX Exploración y Producción, and one anonymous reviewer, for their criticisms and suggestions that contributed to improve the present manuscript. Authors are indebted with to Universidad Autónoma de Baja California Sur, which provided the samples studied in this paper and to the Secretaría de Recursos Hidráulicos for permission to study them and to publish the present results. Thanks are extended to H. Hernández, R. Flores and F. Montaño for continuous help and to Magdalena Alcayde for the English revision of the manuscript. A. Zamora from Carl Zeiss was particularly helpful. This study was partially supported by grant PAPIIT–IN116308.
REFERENCES
Báldi–Beke, M., 1971, The Eocene nannoplankton of the Bakony Mountains, Hungary, in Celebration du centenaire de l'Institut Géologique de Hongrie; Colloque sur la stratgraphie de l'eocene, Budapest, Tihany, 6–8 September, 1969: Magyar Állami Földtani Intézet évkönyve (Annals of the Hungarian Geological Institute), 54(4), 11–25. [ Links ]
Beal, H.C., 1948, Reconnaissance of the geology and oil possibilities of Baja California, Mexico: Geological Society of America, Memoir 31, 138 pp. [ Links ]
Berggren, W.A., Kent, D.V., Swisher III, C.C., Aubry, M.–P. 1995, A revised Cenozoic geochronology and chronostratigraphy in: Berggren, W.A., Kent, D.V., Aubry, M.–P., Hardenbol, I. (eds.), Geochronology, time scales and global stratigraphic correlation: SEMP (Society of Economic Mineralogists and Paleontologists) Special Publication 54, 129–212. [ Links ]
Black, M., 1964, Cretaceous and Tertiary coccoliths from Atlantic seamounts: Paleontology, 5, 306–316. [ Links ]
Black, M., 1971, Problematical microfossils from the Gault clay: Geological Magazine, 108, 325–328. [ Links ]
Black, M., Barnes, B., 1959, The structure of coccoliths from the English Chalk: Geological Magazine, 108, 325–327. [ Links ]
Bramlette, M.N., Martini, E., 1964, The great change is calcareous nannoplankton fossils between the Maestrichtian and Danian: Micropaleontology, 10, 291–322. [ Links ]
Bramlette, M.N., Riedel, W.R., 1954, Stratigraphic value of discoasters and some other microfossils related to Recent coccolitophores: Journal of Paleontology, 28, 385–403. [ Links ]
Bramlette, N., Sullivan, F., 1961, Coccolithophorids and related nannoplankton of the early Tertiary in California: Micropaleontology, 7(2), 129–188. [ Links ]
Bramlette, N., Wilcoxon, A., 1967, Middle Tertiary calcareous nannoplankton of the Cipero section, Trinidad, W. I.: Tulane Studies in Geology and Paleontology, 5, 93–131. [ Links ]
Bown, P.R., Cooper, M.K.E., 1989, Conical calcareous nannofossil in the Mesozoic, in Crux, J.A., van Heck, S.E. (eds.), Nannofossils and their Applications: British Micropalaeontology Society, Series 8, 106 pp. [ Links ]
Bukry, D., 1971, Cenozoic calcareous nannofossils from the Pacific Ocean: Transactions San Diego Society Natural History, 16, 303–327. [ Links ]
Bukry, D., 1973, Low–latitude coccolith biostratigraphy zonation, in Edgar, N.T., Saunders, J.B. et al., Initial Reports of the Deep Sea Drilling Project: Washington, D.C., United States Government Printing Office, 15, 685–703. [ Links ]
Bukry, D., Bramlette, M.N., 1969, Some new and stratigraphically useful calcareous nannofossils of the Cenozoic: Tulane Studies in Geology and Paleontology, 7, 131–142. [ Links ]
Bukry, D., Percival, S. F. 1971, New Tertiary calcareous nannofossils: Tulane Studies in Geology and Paleontology, 8(3), 123–146. [ Links ]
Burns, D.A., 1973, Structural análisis of flanged coccoliths in sediments from south west Pacific Ocean: Revista Española de Micropaleontología, 5, 147–160. [ Links ]
Bybell,L.M., Gartner, S., 1972, Provincialism among mid Eocene calcareous nannofossils: Micropaleontology, 18, 319–336. [ Links ]
Carreño, A.L., Ledesma–Vázquez, J., Guerrero–Arenas, R., 2000, Bioestratigrafía e historia deposicional de la formación Tepetate en el Arroyo Colorado (Eoceno medio–temprano), Baja California Sur, México: Ciencias Marinas, 26(1), 177–200. [ Links ]
Coleman, T.A., 1979, Nannoplankton biostratigraphy of the Tepetate formation, Baja California Sur: USA, University of Southern California, M.Sc. Thesis, 62 pp. (unpublished). [ Links ]
Deflandre, G., 1947, Braarudosphaera nov. Gen., type d'une famille nouvelle de Coccolithophoridés actuels à éléments composites: Compte Rendues Académie Sciences, 225, 439–441. [ Links ]
Deflandre, G., 1950, Observations sur les Coccolithophoriés à propos d'un nouveau type de Braarudosphaeridé, Micrantholithus, à éléments clastiques: Compte Rendus Académie of Sciences, 231, 1156–1158. [ Links ]
Deflandre, G., Fert, C., 1954, Observations sur le coccolithophoridés actuels et fossiles en microscopie ordinaire et électronique: Annales de Paléontologie, 40. 115–176. [ Links ]
Fulwider, R.W., 1976, Biostratigraphy of the Tepetate formation, Baja California Sur: USA, University of Southern California, M.Sc. Thesis, 111 pp. (unpublished). [ Links ]
Gartner, S., 1971, Calcareous nannofossils from the Joides Blake Plateau cores and revision of Paleogene nannofossils zonation: Tulane Studies in Geology and Paleontology, 8(3), 101–121. [ Links ]
Gradstein, F.M., Ogg J., Smith A., 2004, A Geologic Time Scale 2004: Cambridge University Press, pp. 610. [ Links ]
Gran, H.H., Braarud, T., 1935, A quantitative study of the phytoplankton in the Bay of Fundy and the Glf of Main (including observations on hydrogrsphy, chemistry and turbidity): Journal of the Biological Board of Canada, 1, 279–467. [ Links ]
Grassé, P.P., 1952, Traité de Zoologie: Anatomie, Systematique, Biologie. t. 1, fasc.1: Phylogénie. Protozoaires: généralités. Flagellés: Paris, Masson, 1071 pp. [ Links ]
Haq, B.U., 1971, Paleogene calcareous nannoflora, Parts I–IV: Stockholm Contributions in Geology, 25, 1–158. [ Links ]
Hay, W.W., Mohler, H.P., 1967, Calcareous nannoplankton from early Tertiary rocks at Pont Labau, France, and Paleocene–Eocene correlations: Journal of Paleontology, 41, 1505–1541. [ Links ]
Hay, W.W., Towe, K.M., 1962, Electronmicroscopic examination of some coccoliths from Donzacq (France): Eclogae geologica Helvetica, 55, 497–517. [ Links ]
Hay, W.W., Mohler, H.P., Roth, P.H., Schmidt, R.R. Boudreaux, J.E., 1967, Calcareous nannoplankton zonation of the Cenozoic of the Gulf Coast and Caribbean–Antillean area, and transoceanic correlation: Gulf Coast Association of Geological Societies, Transactions, 17, 428–280. [ Links ]
Heim, A., 1922, Notes on the Tertiary of southern lower California: Geological Magazine, 59(12), 529–547. [ Links ]
Instituto Nacional de Estadística, Geografía e Informática (INEGI), 1983, Las Pocitas G12C79, Carta topográfica, escala 1:50,000: Mexico, Instituto Nacional de Estadística, Geografía e Informática, 1 map. [ Links ]
Kamptner, E., 1948, Coccolithen aus dem Torton des Inneralpinen Wierner Beckens: Sitzungsberichte der Österreichische Akademie der Wissenschaften, Mathematische–Naturwissenschaftliche Klasse, Abt. 1, 157, 1–16. [ Links ]
Kamptner, E., 1963, Coccolithineen–Skelettreste aus Tiefseeablagerungen des Pazifischen Ozeans: Annalen des Naturhistorisches Museum Wien, 66, 139–204. [ Links ]
Klumpp, B., 1953, Beitrag zur Kenntnis der Mikrofossilien des mittleren und oberen Eozän: Palaeontographica, Abt. A, 103, 377–406. [ Links ]
Ledesma–Vázquez, J., Rendón–Márquez, G., Carreño, A.L., 1999, Ambientes sedimentarios en la sección arroyo Colorado, formación Tepetate (Eoceno temprano–Medio), Baja California Sur, México: GEOS, 19(2), 78–83. [ Links ]
Locker, S., 1967, Neue coccolithoporiden aus dem Altertertiär Norddeutschlands: Geologie, 16, 361–364. [ Links ]
Locker, S., 1968, Biostratigraphie des Altertiärs von Norddeutschland mit Coccolithophoriden: Monatsberichte der Deutschen Akademie der Wissenschaft zu Berlin, 10, 220–229. [ Links ]
Martini, E., 1958, Discoasteriden und verwandte Formen im NW–deutschen Eozän (Coccolithophorida), 1. Taxonomische Untersuchungen: Senckenbergiana Lethaea, 39, 353–388. [ Links ]
Martini, E., 1959, Pemma angulatum und Micrantholithus basquensis, zwei neue Coccolithoporiden–Arten aus dem Eozän: Senckenbergiana Lethaea, 40, 415–421. [ Links ]
Mina–Uhink, F., 1957, Bosquejo geológico del territorio sur de la Baja California: Boletín de la Asociación Mexicana de Geólogos Petroleros, 9, 188–192. [ Links ]
Minch, J., Leslie, T.A., 1979, Geologic Road Log, La Paz to Tijuana, in Abbott P.L., Gordon R.G. (eds.), Baja California Geology, Geological Society of America Annual Meeting, Field guides and Papers: San Diego, San Diego State University, 159–189. [ Links ]
Miranda–Martínez, A.Y., Carreño, A.L., 2008, Foraminifera from the Tepetate Formation (Heim) at Las Pocitas, Baja California Sur, Mexico: Ciencias Marinas, 34(2), 179–195. [ Links ]
Okada, H., Bukry, D., 1980, Supplementary modification and introduction of code numbers to the low latitude cocccolith biostratigraphic zonation (Bukry, 1973; 1975): Marine Micropaleontology, 5, 321–325. [ Links ]
Perch–Nielsen, K., 1968, Der Feinbau und die Klassifikation der Coccolithen aus dem Maastrichtien von Dänemark: Kongelige Danske Videnskabernes Selskabs Biologiske Skrifter, 16, 96 pp. [ Links ]
Perch–Nielsen, K., 1971a, Neue Coccolithen aus dem Paläozän von Dänemark, der Bucht von Biskaya und dem Eozän der Labador:Bulletin of the Geological Society of Denmark, 21, 51–66. [ Links ]
Perch–Nielsen, K., 1971b, Elektronenmikroskopische Untersuchungen an Coccolithen und verwandten Formen aus dem Eozän von Dänmark: Kongelige Danske Videnskabernes Selskabs Biologiske Skrifter, 18, 76 pp. [ Links ]
Perch–Nielsen, K., 1977, Albian to Pleistocene calcareous nannofossils from the western South Atlantic, DSDP Leg 39, in Supko, P.R., Perch–Nielsen, K., et al., Initial. Reports of Deep Sea Drilling Project, 39: Washington, U.S. Government Printing Office, 699–823. [ Links ]
Perch–Nielsen, K., 1979, Calcareous nannofossil zonation at the Cretaceous/Tertiary boundary in Denmark, in Christensen, W.K., Birkelund, T. (eds.), Cretaceous–Tertiary Boundary Events Symposium, Proceedings: Copenhagen, University of Copenhagen, 1, 115–135. [ Links ]
Perch–Nielsen, K., 1984, Validation of new combinations: International Nannoplankton Association Newsletter, 6, 42–46. [ Links ]
Perch–Nielsen, K., 1985, Mesozoic and Cenozoic calcareous nannofossils, in Bolli, H.M., Saunders, J.B., Perch–Nielsen, K. (eds.), Plankton Stratigraphy: Cambridge University Press, 329–554. [ Links ]
Perch–Nielsen, K., Sadeh, A., Barakat, M.G., Taleb, F., 1978, Late Cretaceous and Early Tertiary calcareous nannofossils and planktonic foraminifera zones from Egypt, in Actes du VIe Colloque Africain de Micropaléontologie, Tunis: Annales des Mines et de la géologie, 28(2), 337–403. [ Links ]
Premoli–Silva, I., Rettori, R., Verga D., 2003, Practical Manual of Paleocene and Eocene Planktonic Foraminifera, International School on planktonic foraminifera, 2nd Course: Università degli Study di Perugia, 152 pp. [ Links ]
Romein, A.J.T., 1977, Calcareous nannofossils from the Cretaceous–Tertiary boundary interval in the Barranco del Gredero (Caravaca, Prov. Murcia, S.E., Spain), Parts I and II: Proceedings of the Koninklijke Nederlandse Akademie van Wetenschappen Amsterdam, Series B, 80, 256–279. [ Links ]
Romein, A.J.T., 1979, Lineages in early Paleogene calcareous nannoplankton: Utrecht Micropaleontological Bulletins, 22, 1–231. [ Links ]
Roth, P.H., 1970, Oligocene calcareous nannoplankton biostratigraphy: Eclogae Geologica Helvetica, 63, 799–881. [ Links ]
Roth, P.H., 1973, Calcareous nannofossils – Leg 17, DSDP, in Winterer, E.L., Ewing. J.I., et al., Initial Reports of the Deep Sea Drilling Project: Washington, D.C., U.S. Government Printing Office, 17, 695–793. [ Links ]
Schiller, J., 1930, Coccolithineae, in Dr. L. Rabenhorst's Kryptogamen–Flora von Deutschland, Osterreich und der Schweiz: Leipzig, Akademische Verlagsgesellschaft, 10(2), 89–267. [ Links ]
Sherwood, R.W., 1974, Calcareous nannofossil systematics, paleoecology and biostratigraphy of the Middle Eocene Weches Formation of Texas: Tulane Sudies in Geology, 11, 1–79. [ Links ]
Shipboard Scientific Party, 2001, Leg 191 Summary in Kanazawa, T., Sanger, W. W., Escutia, C., et al., Proceeding of the Ocean Drilling Program: College Station, TX, Texas A & M University, 191, 1–49. [ Links ]
Stradner, H., 1961, Vorkommen von Nannofossilien im Mesozoikum und altertiär: Erdöl–Zeitschrift, 77, 77–88. [ Links ]
Stradner, H., 1962, Über neue und wenig bekannte Nannofossilien aus Kreide und Alttertiär: Sonderdruck Verhandlungen der Geologischen Bundesanstalt (Wien), 2, 363–377. [ Links ]
Stradner, H., Edwards, A.R., 1968, Electron microscope studies on upper Eocene coccoliths from the Oamaru diatomite, New Zeland: Jahrbuch der Geologischen Bundesanstalt (Wien), 13, 1–66. [ Links ]
Sullivan, F., 1965, Lower Tertiary nannoplankton from the California coast ranges. I Paleocene: University of California, Publications in Geological Sciences, 53, 11–75. [ Links ]
Tan, S.H., 1927, Discoasteridae Incertae Sedis: Proceedings of the Koninklijke Nederlandse Akademie van Wetenschappen Amsterdam, 30, 411–419. [ Links ]
Varol, O., 1989, Eocene calcareous nannofossils from Sile (Northwest Turkey): Revista Española de Micropaleontología, 21, 273–320. [ Links ]
Wallich, G.C., 1877, Observations on the Coccosphere: Annals and Magazine of Natural History, series 4, 19, 342–350. [ Links ]
Wise, S.W., 1973, Calcareous nannofossils from cores recovered during Leg 18, Deep Sea Drilling Project: biostratigraphy and observations on diagenesis, in Kulm, L.D., von Huene, R., et al., Initial Reports of the Deep Sea Drilling Project: Washington, D.C., U.S. Government Printing Office, 18, 569–615. [ Links ]
Wise, S.W., Covington, J.M., Ladner, B., Wei, W. (compilers), 2002, Electronic Calcareous Nannofossils. Version 1.0: International Nannoplankton Association. CD–Rom Series. No. 1. [ Links ]

