










Servicios Personalizados
Revista
Articulo
 Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista mexicana de ciencias geológicas
versión On-line ISSN 2007-2902versión impresa ISSN 1026-8774
Rev. mex. cienc. geol vol.25 no.2 Ciudad de México ago. 2008
Bioestratigrafía de secuencias del Mioceno–Plioceno de la cuenca Macuspana, sureste del Golfo de México
Biostratigraphy of Miocene–Pliocene sequences of the Macuspana basin, southeastern Gulf of Mexico
Jesús Yanina Narváez–Rodríguez1, Javier Helenes–Escamilla1,*, José M. del Moral–Domínguez2, Víctor M. Martínez–Morales2, Consuelo Macías–Ojeda2, Olga G. Castillejos–Zurita2 y María A. Sánchez–Ríos3
1 Centro de Investigación Científica y de Educación Superior de Ensenada (CICESE), Km. 107. Carretera Tijuana–Ensenada, 22860 Ensenada, Baja California, México. * jhelenes@cicese.mx
2 PEMEX Exploración y Producción, Región Sur, Laboratorio de Paleontología y Petrografía, Barranco Sur s/n, Colonia Sur, Zona Industrial en Cobertizo, 86388 Comalcalco, Tabasco, México.
3 Instituto Mexicano de Petróleo, Exploración, Eje Central Lázaro Cárdenas 152, 07730 México, D.F., México.
Manuscrito recibido: Enero 30, 2007
Manuscrito corregido recibido: Diciembre 14, 2007
Manuscrito aceptado: Enero 8, 2008
RESUMEN
Este trabajo presenta el análisis integrado de nanofósiles calcáreos, foraminíferos y registros geofísicos de dos pozos exploratorios en la cuenca Macuspana, sureste del Golfo de México. Esta combinación permite proponer un marco de secuencias estratigráficas para los pozos estudiados. La resolución bioestratigráfica y la confiabilidad de los resultados bioestratigráficos aumentan cuando se integran los resultados de estos dos grupos fósiles. En el pozo A se utilizan las especies Helicosphaera kamptneri, Retículofenestra pseudoumbilica, Minylitha convallis, Discoaster hamatus, Globorotalia obesa y Globorotalia mayeri. Mientras que en el pozo B las especies útiles son: Calcidiscus macintyrei, Retículo fenestra pseudoumbilica, Discoaster quinqueramus y Globorotalia mayeri. En la sucesión sedimentaria del pozo A se reconocen tres candidatos a secuencias estratigráficas, correlacionables con los ciclos de 3er orden: TB–2.6, TB–3.1 y TB–3.5?. Para el pozo B se reconocen cinco candidatos a secuencias estratigráficas, correlacionables con los ciclos de 3er orden: TB–2.6, TB–3.1, TB–3.2, TB–3.3 y TB–3.6?. De acuerdo con los resultados, los sedimentos fueron depositados desde el Mioceno medio hasta el Pleistoceno. Los resultados bioestratigráficos y su relación con la estratigrafía de secuencias permitirán auxiliar en la integración de datos micropaleontológicos para proponer un modelo estratigráfico.
Palabras clave: nanofósiles calcáreos, foraminíferos, bioestratigrafia, Neógeno, cuenca Macuspana, México.
ABSTRACT
This paper presents an integrated analysis of data from calcareous nannofossils,foraminifera and geophysical logs from two exploratory wells in the Macuspana basin, southeastern Gulf of Mexico. This combination permits to propose a sequence stratigraphy frame–work for the studied–wells. Biostratigraphic resolution and reliability increase with the integration of results from both fossil groups. On well A, the following species are used: Helicosphaera kamptneri. Reticulofenestra pseudoumbilica. Minylitha convallis. Discoaster hamatus. Globorotalia obesa and Globorotalia mayeri. On well B, the useful species are: Calcidiscus macintyrei. Reticulofenestra pseudoumbilica. Discoaster quinqueramus and Globorotalia mayeri. In well A, three stratigraphic sequence candidates are recognizable, and they are correlatable with the third order eustatic cycles TB–2.6, TB–3.1 and TB–3.5?. In wellB, five stratigraphic sequence candidates are recognized, which are correlatable with the third order eustatic cycles TB–2.6, TB–3.1, TB–3.2, TB–3.3 and TB–3.6?. Results indicate that the studied sections were deposited from middle Miocene to Pleistocene. The biostratigraphic results, combined with sequence stratigraphy, will help to propose a sequence stratigraphy model.
Key words: calcareous nannofossil, foraminifera, biostratigraphy, Neogene, Macuspana basin, Mexico.
INTRODUCCIÓN
La datación y correlación bioestratigráfica proporcionada por el estudio de grupos planctónicos calcáreos son muy útiles para desarrollar un marco cronoestratigráfico en secciones sedimentarias marinas. Para aumentar la resolución y confiabilidad del marco resultante es recomendable usar bioestratigrafía integrada, la cual consiste en el uso simultáneo de diferentes grupos que puedan suministran datos complementarios. La integración de resultados ayuda a definir con más detalle las líneas de tiempo indicadas por cada grupo taxonómico y permite identificar los eventos paleobiológicos de importancia, susceptibles de ser utilizados en la cronoestratigrafía (Arenillas, 2002). Para fines cronoestratigráficos, entre los principales grupos de microfósiles se encuentran los nanofósiles calcáreos y los foraminíferos planctónicos. Estos grupos son ampliamente reconocidos por ser una herramienta de gran utilidad en las correlaciones bioestratigráficas de carácter regional y global (Brasier, 1980; Raffi, 1999). La bioestratigrafía integrada también se puede usar en estudios estratigráficos de secuencias, principalmente para correlacionar los ciclos depositacionales locales dentro de un marco global (Haq et al, 1988; Haidenbol et al., 1998).
La cuenca Macuspana se encuentra en el extremo suroriental del Golfo de México (Ambrose et al., 2003). Los estratos de facies marinas someras de la cuenca Macuspana se caracterizan por la escasez y hasta ausencia de biomarcadores estratigráficos marinos convencionales. Tampoco poseen minerales autigénicos que permitan datar radiométricamente los estratos involucrados, por lo que es difícil su correlación precisa. A fin de compensar las deficiencias bioestratigráficas mencionadas, realizamos un estudio bio–estratigráfico cuantitativo con varios grupos de microfósiles con la finalidad de cubrir todas las facies sedimentarias e incrementar las posibilidades de recuperación de especies marcadoras.
Como en toda región petrolera, la bioestratigrafía es una importante herramienta en el Golfo de México. Hay algunas publicaciones sobre estudios bioestratigráficos de la parte norte del Golfo de México (Gartner et al, 1983; Weiterman, 1989; Aubry, 1993; Zhang et al., 1993; Wei, 2003). Los trabajos bioestratigráficos publicados de la parte sureste del Golfo de México son pocos, por lo que consideramos importante documentar los conjuntos de microfósiles planctónicos para ayudar a establecer la bioestratigrafía en esta importante región. El análisis bioestratigráfico basado principalmente en foraminíferos planctónicos y bentónicos en la cuenca Macuspana ha permitido conocer especies correlacionares por su distribución de carácter regional (Castillejos et al., 2001).
La escasez de publicaciones sobre estudios bioestratigráficos cuantitativos de la cuenca Macuspana dificulta la elaboración de un marco bioestratigráfico que ayude a modelar su evolución geológica. El presente estudio intenta contribuir a lograr un conocimiento más detallado de la bioestratigrafía y sus relaciones con la estratigrafía de secuencias para la cuenca Macuspana.
ANTECEDENTES
Marco estratigráfico
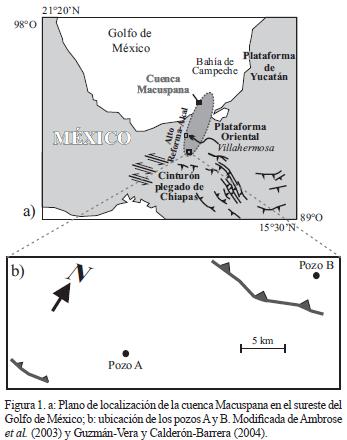
La cuenca Macuspana se localiza en el sureste del Golfo de México, abarcando parte del Estado de Tabasco en el cual se localizan los pozos Ay B (Figura 1). Está conformada por una estructura de tipo graben (Santiago–Acevedo, et al., 1984). Cubre el área de 7,300 km2 continentales y aproximadamente 1,800 km2 marinos (Guzmán–Vera y Aranda–García, 2002). Limita hacia el sur con la Sierra de Chiapas, hacia el este con la plataforma de Yucatán, hacia el oeste con el Alto de Reforma–Akal y al norte se interna en el Golfo de México (Ambrose etal., 2003). Comprende una sucesión sedimentaria de rocas siliciclásticas del Mesozoico y Cenozoico que sobreyace a un basamento de rocas cristalinas de edad precámbrica y paleozoica (Sedlock et al., 1994). La columna litoestratigráfica de la cuenca depositada durante el Neógeno está constituida por las formaciones Belem, Zargazal, Encajonado, Amate superior, Amate inferior, Encarnación y Caliza Macuspana propuestas por Galloway et al. (1991; Figura 2).
Durante la evolución de la cuenca Macuspana intervinieron, desde el noroeste, los eventos de la Orogenia Laramide que afectó al noreste de México y soroeste de Estados Unidos a fines del Cretácico y principios del Cenozoico. En el Mioceno fue sometida nuevamente a esfuerzos tectónicos, esta vez desde el sur, relacionados con el levantamiento de la Sierra de Chiapas (Sedlock et al., 1994). Tectónicamente, la cuenca está ubicada en la parte norte del sistema de fallas Polochic y Motagua del sur de México, y está dominada por la interacción de las placas Norteamérica, Cocos y Caribe.
MATERIALES Y MÉTODOS
El material utilizado corresponde a dos pozos exploratorios que incluyen muestras de canal y de núcleo. Para el análisis de nanofósiles se procesaron 117 muestras del pozo A y 106 muestras del pozo B, siguiendo la técnica de frotis (Martini, 1971; Bowny Young, 1998). Para eliminar el lodo de perforación de emulsión inversa de algunas de las muestras se aplicó la técnica propuesta por Styzen y Reugger (1994), la cual consiste en limpiar las muestras con hipoclorito de sodio para eliminar los polímeros y facilitar la visualización de los nanofósiles. Estos microfósiles fueron observados en un microscopio óptico de luz transmitida, con magnificación de 1000 aumentos. Se realizó el conteo de la población total de especímenes en una lámina por muestra, aunque en láminas con poca recuperación se contaron un mínimo de 300 especímenes. Las identificaciones se basaron en la taxonomía propuesta por Perch–Nielsen (1985), Aubry (1984, 1988, 1989, 1990), Young (1998) y la asistencia de la base de datos compilada por Wise et al. (2004). Los datos de foraminíferos utilizados en este estudio y los registros geofísicos fueron proporcionados por el Laboratorio de Paleontología y Petrografía, Activo de Exploración Macuspana, PEMEX Exploración y Producción (PEP).
RESULTADOS
Los nanofósiles calcáreos son comunes en la mayoría de los niveles, con preservación moderada. Algunos especímenes están afectados por disolución y fragmentación. Algunos autores (Rothy Thierstein, 1972; Roth, 1983) han propuesto que la preservación de los nanofósiles calcáreos está en relación con la profundidad de depositación. Sin embargo, la paleobatimetría de las columnas estudiadas es generalmente somera, por lo que creemos que la disolución en las muestras examinadas se debe en su mayoría a corrosión por los compuestos químicos utilizados en el lodo de perforación. La edad numérica de los eventos estratigráficos utilizada corresponde a la propuesta por Berggren et al. (1995), Gradstein et al. (2004) y la encontrada en la base de datos de Wise et al. (2004). La interpretación bioestratigráfica de los foraminíferos planctónicos fue realizada con base en los rangos bioestratigráficos propuestos por Kennett y Srinivasan (1983) y Bolli y Saunders (1985) para el Neógeno.
Se determinaron los bioeventos de nanofósiles calcáreos y foraminíferos planctónicos para asignar una edad consenso, documentar cambios en los conjuntos de cada grupo fósil y correlacionar las secciones estratigráficas. Debido a que los estudios de nanofósiles y foraminíferos se hicieron en muestras de canal, hay la posibilidad de contaminación dentro del pozo, por derrumbe de material joven (caído) en niveles más antiguos. Para minimizar la posibilidad de contaminación por material caído, los bioeventos utilizados son principalmente las últimas apariciones evolutivas (cimas), aunque en el pozo B también se usó la primera aparición evolutiva (base) de una especie. Los datos de nanofósiles calcáreos son el resultado de un estudio cuantitativo, mientras que los datos de foraminíferos planctónicos están basados en estudios cualitativos. A continuación se describen las evidencias para asignar las edades consenso de ambos pozos. Para cada especie se incluyen entre paréntesis los alcances estratigráficos datados radiométricamente.
Para las edades consenso del pozo A se utilizaron las siguientes especies:
Helicosphaera kamptneri (23.9–0.0 Ma); nanofósil (en el nivel 1 m).
Reticulofenestra pseudoumbilica (11.8–3.8 Ma); nanofósil (en el nivel 1,040 m).
Minylitha convallis (10.8–5.6 Ma); nanofósil (en el nivel 1,500 m).
Discoaster hamatus (10.8–9.4 Ma); nanofósil (en el nivel 2,160 m).
Globorotalia obesa (21.5–10.9 Ma); foraminífero planctónico (en el nivel 3,380 m).
Globorotalia mayeri (27.1–11.4 Ma); foraminífero planctónico (en el nivel 3,640 m).
Reciente–Plioceno (1–1,040 m). Limitado por las cimas de H. kamptneri (1 m) y R. pseudoumbilica (1,040 m). No es posible diferenciar con mayor resolución este intervalo. Aunque se observan escasos especímenes del Plioceno, estos se consideran retrabajados.
Plioceno temprano (1,040–1,500 m). Limitado por las cimas de R. pseudoumbilica (1,040 m) y M. convallis (1,500 m). Especímenes del género Reticulofenestra (tamaño menor a 7 µm) ocurren desde el inicio de la sección, pero la cima de R. pseudoumbilica (8 µm) se observa a los l,040m. Aunque está incluida en este intervalo, la cima de Globigerinoides obliquus obliquus (3.5 Ma; en el nivel l,490m) no se usó como evidencia del Plioceno temprano, por encontrarse estratigráficamente más abajo que la de R. pseudoumbilica. Sin embargo, apoya la asignación de edad.
Mioceno tardío (1,500–2,160 m). Limitado por las cimas de M. convallis (1,500 m) y D. hamatus (2,160 m). Este intervalo incluye el alcance estratigráfico total de la especie M. convallis. La cima de Globorotaloides variabilis (5.2 Ma; en el nivel 2,100 m) no se usó como evidencia del Mioceno tardío por encontrarse estratigráficamente más abajo que la de M. convallis. Sin embargo, apoya la asignación de edad.
Mioceno tardío al Mioceno medio (2,160–3,380 m). Caracterizado por las cimas de D. hamatus (2,160m) y G. obesa (3,380 m). Este intervalo incluye el alcance estratigráfico total de la especie D. hamatus.
Mioceno medio (3,380–3,640 m). Limitado por las cimas de G. obesa (3,380 m) y G. mayeri (3,640 m). En el nivel 3,380 m se usó la edad radiométrica de la extinción de G. obesa (10.9 Ma), por ser más joven que la de Sphenolithus heteromorphus (13.6 Ma), la cual ocurre en el mismo nivel de G. obesa. Sin embargo, la presencia de S. heteromorphus también apoya esta asignación de edad. Mioceno medio a temprano (3,640–3,650 m). Asignación de edad basada en la presencia de las especies G. mayeri (27.1– 11.4 Ma) y G. obesa (21.5–10.9 Ma) en este intervalo.
Para las edades consenso del pozo B se utilizaron las siguientes especies:
Calcidiscus macintyrei (18.4–1.8 Ma); nanofósil (en el nivel 40m).
Reticulofenestra pseudoumbilica (11.8–3.8 Ma); nanofósil (en el nivel 280m).
Discoaster quinqueramus (8.6–5.6 Ma); nanofósil (en el nivel 440m).
Discoaster berggrenü (8.6–5.6 Ma); nanofósil (en el nivel 3,000m).
Globorotalia mayen (27.1–11.4 Ma); foraminífero planctónico (en el nivel 3,885m).
Globorotalia obesa (21.5–10.9 Ma); foraminífero planctónico (en el nivel 3,895m).
Pleistoceno–Plioceno (40–280 m). Intervalo caracterizado por las cimas de C. macintyrei (40 m) y R. pseudoumbilica (280 m). Pocos fósiles fueron observados en este intervalo.
Plioceno temprano (280–440 m). Intervalo caracterizado por las cimas de R. pseudoumbilica (280m) y D. quinqueramus (440m). En este pozo B, no se consideró la cima del foraminífero G. obliquus obliquus (16.4–3.5Ma; en el nivel 1435m) para establecer la edad consenso debido a que apareció estratigráficamente más abajo que R. pseudoumbilica.
Mioceno tardío (440–3,000 m). Intervalo comprendido entre la cima de D. quinqueramus (440m) y la base (primera aparición evolutiva) de D. quinqueramus (3,000m).
Este intervalo incluye los alcances estratigráficos totales de D. quinqueramus y D. berggrenii.
Mioceno tardío–Mioceno medio (3,000–3,885 m). Intervalo entre la base de D. quinqueramus (3000m) y la cima de G mayeri (3,885 m). Pocos fósiles fueron observados en este intervalo.
Mioceno medio (3,885–3,915 m). Determinado por la presencia de G. mayeri en todo el intervalo (3,885–3,915m), el cual llega a la profundidad total del pozo. La última aparición evolutiva del nanofósil S. heteromorphus (18.1–13.6Ma; en el nivel 3,895 m) por debajo de la tubería de revestimiento a los 2,692 m, indica un ligero aumento de la edad, pero aun en el Mioceno medio. En este intervalo se observa la especie G. mayeri, mientras que la presencia de G. obesa (prof. 3,895 m) apoya la asignación de edad.
La distribución de foraminíferos bentónicos constituyó la base principal de la reconstrucción paleobatimétrica y su diversidad permitió la discriminación de ambientes. Se utilizó la zonación paleobatimétrica propuesta por PEMEX–IMP basada en la clasificación de ambientes marinos y zonas ecológicas de la costa del Golfo de México propuesta por Tipsword (1962) y los rangos paleobatimétricos propuestos por Van Morkhoven et al. (1986), Breard et al. (1993) y Katz y Miller (1993). Siete conjuntos de foraminíferos bentónicos fueron utilizados para establecer las condiciones paleobatimétricas en la cuenca Macuspana, los cuales permitieron determinar los ambientes: transicional, nerítico interno, nerítico medio, nerítico externo, batial superior, batial medio y batial inferior.
Cronoestratigrafía de secuencias
La estratigrafía de secuencias subdivide el registro sedimentario en unidades limitadas por discordancias (secuencias), formadas durante cambios del nivel del mar (Vincent et al, 1998). Con base en los datos bioestratigráficos mencionados anteriormente, y en la interpretación de los registros geofísicos (rayos gamma, potencial espontáneo y resistividad) proporcionados por PEMEX, se proponen candidatos a secuencias estratigráficas y su correlación respectiva con la carta global de ciclos eustáticos propuesta por Haq et al (1988) y Hardenbol et al (1998). Las siguientes son secuencias estratigráficas interpretadas y ciclos de tercer orden para los pozos A y B.
Para el pozo A, se reconocieron tres secuencias estratigráficas (Figura 3):
Secuencia TB–2.6. Se caracteriza por una superficie de inundación máxima en el nivel 3,380 m, de 11.6 Ma de edad. El límite inferior está por debajo de la profundidad total del pozo y el límite superior se localiza a la profundidad de 2,500 m asignado a la edad de 10.5 Ma. Este intervalo contiene las últimas apariciones evolutivas de Globorotalia obesa y G. mayeri. Cronoestratigráficamente, esta secuencia corresponde al Mioceno medio y tardío.
Secuencia TB–3.1. Se caracteriza por una superficie de inundación máxima en el nivel 2,080 m, de 9.2 Ma de edad. El límite superior se localiza a 900 m, asignado a la edad de 8.2 Ma. Este intervalo contiene las últimas ocurrencias de Minylitha convallis, Discoaster hamatus y Reticulofenestra pseudoumbilica. Por posición estratigráfica, esta secuencia corresponde al Mioceno tardío y Plioceno temprano.
Secuencia TB–3.5? Se caracteriza por una superficie de inundación máxima en el nivel 200 m, de 4.0 Ma de edad. El límite inferior se encuentra en el nivel 900 m, considerándose la edad de 4.2 Ma, y el límite superior probablemente se localizaría en la cima de la sección. Las secuencias TB–3.1 y TB–3.5? están separadas por una discordancia en el nivel de 900 m por lo que se considera la edad de 4.2 y 8.2 Ma en el límite de ambas secuencias. Esta secuencia es asignada al Plioceno temprano.
En el pozo B se reconocieron cuatro secuencias estratigráficas, las cuales se detallan a continuación (Figura 4):
Secuencia TB–2.6. Se caracteriza por una superficie de inundación máxima en el nivel 3850m, de 11.6 Ma de edad. El límite superior se localiza a la profundidad de 3,300 m y es asignado a la edad de 10.5 Ma. La última aparición evolutiva de G. mayeri está contenida en este intervalo. Cronoestratigráficamente, esta secuencia corresponde al Mioceno medio y tardío.
Secuencia TB–3.1. Se caracteriza por una superficie de inundación máxima en el nivel 2,700 m, de 9.2 Ma de edad. El límite superior se localiza a la profundidad de 2,140 m asignado a la edad de 8.2 Ma. Este intervalo contiene la primera ocurrencia de Discoaster quinqueramus.
Secuencia TB–3.2. Se caracteriza por presentar una superficie de inundación máxima en el nivel 1,760 m, de 7.0 Ma de edad. El límite superior se localiza a la profundidad de 1,000 m, asignado a la edad de 6.3 Ma. La edad asignada de esta secuencia es de Mioceno tardío.
Secuencia TB–3.3. Se caracteriza por una superficie de inundación máxima en el nivel 780 m, de 5.8 Ma de edad. El límite superior es considerado a 420 m, correspondiente a la edad de 5.5 Ma. Este intervalo contiene la última aparición evolutiva de Discoaster quinqueramus. La edad que comprende esta secuencia es Mioceno tardío y Plioceno temprano.
Secuencia TB–3.6? Esta secuencia se caracteriza por una superficie de inundación máxima en el nivel 230m, de 3.4 Ma de edad. El límite inferior corresponde al nivel 420 m, de 3.8 Ma. Este intervalo contiene las últimas ocurrencias de Calcidiscus macintyrei y Reticulofenestra pseudoumbilica. Entre las secuencias TB–3.3 y TB–3.6? se identifica una discordancia a la profundidad de 420 m. El límite superior es considerado a 40 m con edad probable de 3.0 Ma Cronoestratigráficamente, esta secuencia es tentativamente asignada a la edad de Plioceno temprano a Pleistoceno.
DISCUSIÓN
Durante el Neógeno en la cuenca Macuspana, la especie Reticulofenestra pseudoumbilica es un fósil índice que tiene su primera ocurrencia en ambos pozos, mientras que la especie Globigerinoides obliquus obliquus aparece estratigráficamente más abajo. La cima del intervalo Mioceno tardío está indicada por la ocurrencia de Minylitha convallis y Discoaster hamatus en el pozo A y el alcance estratigráfico total de Discoaster quinqueramus en el pozo B. Las especies de Globorotalia obesa y Globorotalia mayeri, ocurren en ambos pozos indicando una edad del Mioceno medio. La posición estratigráfica de estas especies en el pozo B varía ya que se tiene la primera ocurrencia de G. mayeri (11.4 Ma) y posteriormente ocurre G obesa (10.9 Ma).
Los candidatos de secuencias estratigráficas y correlaciones con los ciclos de tercer orden (Haq et al., 1988; Hardenbol et al., 1998) determinados en los pozos varían. En los pozos Ay B se observa una discordancia entre las secuencias estratigráficas del Plioceno temprano. Entre las secuencias TB–3.1 y TB–3.5? en el pozo A ocurre una discordancia entre el límite superior del ciclo TB–3.1, de 8.2 Ma, y el límite inferior de TB–3.5?, de 4.2 Ma, resultando 4.0 m.a. de intervalo faltante en la sección analizada, comprendiendo las edades del Reciente al Plioceno. Las secuencias TB–3.3 y TB–3.6? en el pozo B presentan el límite superior de 5.5 Ma y el límite inferior de 3.8 Ma, obteniéndose 1.7 m.a. de intervalo faltante, considerándose entre las edades de Pleistoceno al Plioceno.
CONCLUSIONES
El estudio bioestratigráfico integrado permitió la subdivisión del Neógeno de la cuenca Macuspana en los pozos exploratorios A y B. La sección sedimentaria del pozo A es del Reciente a Mioceno medio, y del pozo B de Pleistoceno a Mioceno medio. Los foraminíferos bentónicos reconocen ambientes someros a profundos, representados por una paleobatimetría transicional, nerítico interno, nerítico medio, nerítico externo a batial superior. Los candidatos a secuencias estratigráficas y ciclos de tercer orden se determinaron en la sucesión sedimentaría del pozo A: TB–2.6, TB–3.1 y TB–3.5?, mientras que, en el pozo B se determinó: TB–2.6, TB–3.1, TB–3.2, TB–3.3 y TB–3.6?. La presencia de la discordancia varía en la estimación de la duración de su ocurrencia en los pozos A y B.
AGRADECIMIENTOS
Esta investigación forma parte del trabajo que se viene desarrollando en los estudios doctorales de la primera autora, auspiciados por la Secretaria de Relaciones Exteriores (S.R.E.) y el Centro de Investigación Científica y de Educación Superior de Ensenada (CICESE). Agradecemos a PEMEX (Activo de Exploración Macuspana) por proporcionar muestras, información litológica, registros geofísicos y base de datos de foraminíferos de los pozos. También agradecemos a la Oceanóloga Edna Collins del CICESE por su apoyo en el procesamiento de muestras de microfósiles.
REFERENCIAS
Ambrose, W.A., Wawrzyniec, T.F., Fouad, K., Talukdar, S.C., Jones, R.H., Jennette, D.C., Holtz, M.H., Sakurai, S., Dutton, S.P., Dunlap, D.B., Guevara, E.H., Meneses–Rocha, J., Lugo J., Aguilera, L., Berlanga, J., Miranda, L., Ruiz, J., Rojas, R., Solís, H., 2003, Geologic framework of upper Miocene and Pliocene gas plays of the Macuspana Basin, Southeastern Mexico: American Association of Petroleum Geologists Bulletin, 87(9), 1411–1435. [ Links ]
Arenillas, I., 2002, Bioestratigrafía integrada y sus aplicaciones, en Molina, E., (ed.), Micropaleontología: Prensas Universitarias de Zaragoza, Colección Textos Docentes, 93, 539–554. [ Links ]
Aubry, M.–P., 1984, Handbook of Cenozoic Calcareous Nannoplankton, Book 1: Ortholithae (Discoasters): New York, Micropaleontology Press, 266 p. [ Links ]
Aubry, M.–P., 1988, Handbook of Cenozoic Calcareous Nannoplankton, Book 2: Ortholithae (Catinasters, Ceratoliths and Rhabdoliths): New York, Micropaleontology Press, 279 p. [ Links ]
Aubry, M.–P., 1989, Handbook of Cenozoic Calcareous Nannoplankton, Book 3: Ortholithae (Pentaliths and Others). Heliolithae (Fasciculiths, Sphenoliths and Others): New York, Micropaleontology Press, 279 p. [ Links ]
Aubry, M.–P., 1990, Handbook of Cenozoic Calcareous Nannoplankton, Book 4: Heliolithae (Helicoliths, Cribriliths, Lopadoliths and Others): New York, Micropaleontology Press, 381 p. [ Links ]
Aubry, M.–P., 1993, Neogene allostratigraphy and depositional history of the De Soto Canyon area, northern Gulf of Mexico: Micropaleontology, 39, 327–366. [ Links ]
Berggren, W.A., Kent, D.V., Swisher, C.C., III, Aubry, M.–R., 1995, A revised Cenozoic geochronology and chronostratigraphy, in Berggren, W.A., Kent, D.V., Aubry, M.–R., Hardenbol, J., (eds.), Geochronology, Time Scales and Global Stratigraphic Correlation: Tulsa, Oklahoma, Society for Sedimentary Geology, SEPM, Special Publication, 54, 129–212. [ Links ]
Bolli, H.M., Saunders, J.B., 1985, Oligocene to Holocene low latitude planktic foraminifera, in Bolli, H.M., Saunders, J.B., Perch–Nielsen, K. (eds.), Plankton Stratigraphy: Cambridge, Cambridge Univ. Press, 155–262. [ Links ]
Brasier, M., 1980, Phyllum Sarcodina–Foraminifera, in Brasier, M., (ed.), Microfossils: London, George Allen & Unwin, 90–121. [ Links ]
Bown, P., Young, J., 1998, Techniques, in Bown, PR. (ed.), Calcareous nannofossil biostratigraphy: Kluwer Academic, British Micropalaeontological Society Series, 16–28. [ Links ]
Breard, S., Callender, A., Nault, J., 1993, Paleoecologic and biostratigraphic models for Pleistocene through Miocene foraminiferal assemblages of the Gulf Coast Basin: Gulf Coast Association of Geological Societies, 43, 493–502. [ Links ]
Castillejos, O., Granados, O., Olivera, C., Hernández, R., Martínez, V., Saynez, A., Beaurregad, S., 2001, Estudio bioestratigráfico de la Cuenca Terciaria de Macuspana: Revista Ingeniería Petrolera Asociación de Ingenieros Petroleros de México, 43–50. [ Links ]
Galloway, W., Bebout, D., Fisher, W., Dunlap, J., Cabrera–Castro, R., Lugo–Rivera, I., Scott, T., 1991, Cenozoic, in Salvador, A., (ed.), The Gulf of Mexico Basin: Boulder, Colorado, Geological Society of America, The Geology of North America, J, 245–324. [ Links ]
Gartner, S., Chen, M.R., Stanton, R.J., 1983, Late Neogene nannofossil biostratigraphy and paleoceanography of the northeastern Gulf of Mexico and adjacent areas: Marine Micropaleontology, 8, 17–50. [ Links ]
Gradstein, E., Ogg, J., Smith, A., 2004, The Neogene Period, in Gradstein, E, Ogg, J., Smith, A. (eds.), A Geologic Time Scale 2004: United Kingdom, Cambridge University Press, 409–440. [ Links ]
Guzmán–Vera, E., Aranda–García, M., 2002, Inversión estructural, un nuevo concepto sobre la evolución de la Cuenca de Macuspana y sus implicaciones en la acumulación de hidrocarburos (resumen), en III Reunión Nacional de Ciencias de la Tierra, Puerto Vallarta, Jal.:Geos, 22(2), p. 214. [ Links ]
Guzmán–Vera E., Calderón–Barrera, J.A., 2004, Direct hydrocarbon indicators and structural inversion in the south of the Macuspana basin, an addition for hydrocarbons search, en America Association of Petroleum Geologists International Conference, October 24–27, Cancún, México. [ Links ]
Haq, B., Hardenbol, J., Vail, P., 1988, Mesozoic and Cenozoic chronostratigraphy and cycles of sea level change, in Wilgus, C., Hastings, B., Posamentier, H., van Wagoner, J., Ross, C., Kendall, C., (eds.), Sea–level Change: an Integrated Approach: Tulsa, Oklahoma, Society for Sedimentary Geology, SEPM, Special Publication, 42,71–108. [ Links ]
Hardenbol, J., Thierry, J., Farley, M., Jacquin, T, De Graciansky, P.–C, Vail, P., 1998, Cenozoic sequence chronostratigraphy, en De Graciansky, P.–C, Hardenbol, J., Jacquin, T, Vail, P. (eds.), Mesozoic and Cenozoic Sequence Stratigraphic of European Basins: Tulsa, Oklahoma, Society for Sedimentary Geology, SEPM, Special Publication, 60, 3–13. [ Links ]
Katz, M., Miller, K., 1993, Latest Oligocene to Earliest Pliocene benthic foraminiferal biofacies of the northeastern Gulf of Mexico: Micropaloeontology, 39, 367–403. [ Links ]
Kennett, J., Srinivasan, M., 1983, Neogene Planktonic Foraminifera: Stroudsburg, Pennsylvania, Hutchinson Ross Publishing Company, 265 p. [ Links ]
Martini, E., 1971, Standard Tertiary and Quaternary calcareous nannoplankton zonation, in Farinacci, A. (ed.), Proceedings of the Second International Conference on Planktonic Microfossils: Roma, Ed. Tecnoscienza, v. 2, 739–785. [ Links ]
Perch–Nielsen, K., 1985, Cenozoic calcareous nannofossils, in Bolli, H.M., Saunders, J.B., and Perch–Nielsen, K. (eds.), Plankton Stratigraphy: Cambridge, Cambridge Univ. Press, 427–554. [ Links ]
Raffi, I., 1999, Precision and accuracy of nannofossil biostratigraphic correlation: Philosophical Transactions of the Royal Society London, A, 357, 1975–1993. [ Links ]
Roth, P., 1983, Jurassic and lower Cretaceous calcareous nannofossils in the western Atlantic (Site 534): Biostratigraphy, preservation and observations on biogeography and paleoceanography, en Sheridan, R.E., Gradstein, F.M., et al., Initial Reports of the Deep Sea Drilling Project: Washington, U.S. Government Printing Office, 76, 587–621. [ Links ]
Roth, P., Thierstein, H., 1972, Calcareous nannoplankton: Leg 14 of the Deep Sea Drilling Project, en Hayes, D.E., Pimm, A.C., et al, Initial Reports of the Deep Sea Drilling Project: Washington, U.S. Government Printing Office, 14, 421–485. [ Links ]
Santiago–Acevedo, J., Carrillo–Bravo, J., Martell–Andrade, B., 1984, Geología Petrolera de México, en Marmissolle–Daguerre, D. (coord.), Evaluación de Formaciones en México: México, D.F., Schlumberger y Petróleos Mexicanos (PEMEX), I–1 – I–36. [ Links ]
Sedlock, R., Ortega, G., Speed, R., 1994, Tectonostratigraphic terranes and tectonic evolution of México: Geological Society of America, Special Paper 278, 153 p. [ Links ]
Styzen, M., Reugger, L., 1994, Polymer mud: Problems and solution: Journal of Nannoplankton Research, 16(1), 19–20. [ Links ]
Tipsword, H., 1962, Tertiary foraminifera in Gulf Coast petroleum exploration and development, in Rainwater, E., Zingula, R., (eds.), Geology of the Gulf Coast and Central Texas and guidebook of excursions: Houston, Texas, Houston Geological Society, 16–57. [ Links ]
Van Morkhoven, F., Berggren, W., Edwards, A., 1986, Cenozoic cosmopolitan deep–water benthic foraminifera: Bulletin des Centres de Recherches Exploration–Production Elf–Aquitaine, 11, 421p. [ Links ]
Vincent, S., Macdonald, D., Gutteridge, P., 1998, Sequence Stratigraphy, in Doyle, P., Bennett, M., (eds.), Unlocking the stratigraphic record: advances in modern stratigraphy: Chichester, John Wiley and Sons, 299–350. [ Links ]
Wei, W., 2003, Upper Miocene nannofossil biostratigraphy and taxonomy of Exxon core CH30–43–2 from the Gulf of Mexico: Journal of Nannoplankton Research, 25(1), 17–23. [ Links ]
Weiterman, S.D., 1989, Neogene calcareous nannofossil biostratigraphy of the West Florida Carbonate ramp Slope, Northeast Gulf of Mexico: Florida, Florida State University, tesis de maestría. [ Links ]
Wise, S.W., Covington, J.M., Ladner, B.C., Wei, W. (compilers), 2004, Electronic Calcareous Nannofossil, v. 3: International Nannoplankton Association, CD–ROM series, no. 1. [ Links ]
Young, J., 1998, Neogene, in Bown, PR. (ed.), Calcareous Nannofossil Biostratigraphy: London, Kluwer Academic Publ., 225–265. [ Links ]
Zhang, J., Miller, K., Berggren, A., 1993, Neogene planktonic foraminiferal biostratigraphy of the northeastern Gulf of Mexico: Micropaleontology, 39, 299–326. [ Links ]

