










Servicios Personalizados
Revista
Articulo
 Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista mexicana de ciencias geológicas
versión On-line ISSN 2007-2902versión impresa ISSN 1026-8774
Rev. mex. cienc. geol vol.23 no.3 Ciudad de México ene. 2006
Sección especial
The genus Phlyctenodes Milne Edwards, 1862 (Crustacea: Decapoda: Xanthidae) in the Eocene of Europe
Alessandra Busulini1, Giuliano Tessier2, and Claudio Beschin3
1 c/o Museo di Storia Naturale, S. Croce 1730,1 – 30125, Venezia, Italia. busulini@aitin.it
2 via Barbarigo 10,1– 30126, Lido di Venezia, Italia.
3 Associazione Amici del Museo Zannato, Piazza Marconi 15,1 – 36075, Montecchio Maggiore (Vicenza), Italia.
Manuscript received: September 2, 2005
Corrected manuscript received: January 17, 2006
Manuscript accepted: January 25, 200
ABSTRACT
A systematic review of the crab genus Phlyctenodes Milne Edwards, 1862 is carried out. Based on carapace features, this taxon is placed in the subfamily Actaeinae, family Xanthidae MacLeay 1838. Species attributed to this genus are known from Eocene reef environments in Europe. Preservation of crustacean remains in this kind of environment is very rare, and it could explain scarcity of specimens of this genus. For the first time, pictures of types of this genus described during the XIX century and the first decades of the XX century are presented. A study of recently collected specimens from the Eocene of Véneto (Italy) allows to clarify relationships between Phlyctenodes krenneri Lörenthey. 1898 and P. dalpiazi Fabiani, 1911. Presence of P. tuberculosus Milne Edwards, 1862 among the new material is documented. The otherknown species of this genus, P. hantkeni Lörenthey. 1898 is placed in Pseudophlyctenodes new genus on the basis of differences in morphological features.
Key words: Crustacea, Decapoda, Phlyctenodes. systematic review, Eocene, Italy.
RESUMEN
Se presenta una revisión sistemática del género de cangrejo Phlyctenodes Milne Edwards, 1862. Con base en las características del caparazón, este taxon es ubicado en la subfamilia Actaeinae, familia Xanthidae MacLeay, 1838. Las especies de este género son conocidas de ambientes arrecifales en el Eoceno de Europa. La preservación de restos de crustáceos en este tipo de ambiente en muy rara, lo que podría explicar la escasez de especímenes de este género. Se presentan por vez primera fotografías de ejemplares tipo de este género descritos durante el siglo XIXy las primeras décadas del siglo XX. El estudio de especímenes recolectados recientemente en el Eoceno de Véneto (Italia) permite aclarar las relaciones entre Phlyctenodes krenneri Lörenthey, 1898, y P dalpiazi Fabiani, 1911. Se documenta la presencia de P tuberculosus Milne Edwards, 1862 entre los ejemplares recolectados. La otra especie conocida de este género, P. hantkeni Lörenthey. 1898, es ubicada en Pseudophlyctenodes nuevo género, con base en diferencias en las características morfológicas.
Palabras clave: Crustacea, Decapoda, Phlyctenodes. revisión sistemática, Eoceno, Italia.
INTRODUCTION
Fossil crabs housed in the Museo civico "G. Zannato" (Montecchio Maggiore, Vicenza, Italy) and discovered in the Colli Berici (Berici Hills, NE Italy) –precisely at San Feliciano and Campolongo di San Germano (Vicenza)– represent four species referred to the genus Phyctenodes Milne Edwards, 1862, including P. dalpiazi Fabiani, 1911, P. krenneri Lörenthey, 1898, P. steinmanni Lörenthey, 1902, and P. tuberculosas Milne Edwards, 1862. The fossils were collected in Late Eocene limestones rich in corals, algae and crustaceans remains (De Angeli and Garassino, 2002).
The new findings have prompted to carry out a review oí Phlyctenodes Milne Edwards, 1862, represented by a small number of Eocene species, all of them distributed only in Europe (Figure 1). This study is suitable for publication of a photographic testimony of all type specimens referred to species of Phlyctenodes, previously known only by drawings. P. krenneri Lörenthey, 1898 and P. steinmanni Lörenthey, 1902 were reported from several localities, represented by a small number of specimens. Only holotypes are known for P. pustulosus Milne Edwards, 1862, and P. nicolisi Bittner, 1884. P. tuberculosus Milne Edwards, 1862, the generic type species, and P. dalpiazi Fabiani, 1911 were in the same situation, but new specimens of these two species are herein documented.
Scarcity of findings is probably caused by the low preservation potential for crustacean remains of the coral environment where these species lived. This paleoenvironment is confirmed by several studies (Lörenthey and Beurlen, 1929; Di Salvo, 1933; Miiller and Collins, 1991; Beschin et al, 2000). Reports of this genus are also scarce in Eocene outcrops from Véneto, Italy, most of them from the Lessini Mounts. After the studies of Bittner (1884) on P. nicolisi and of Fabiani (1911) on P. dalpiazi, in the last decades the genus has been found again in Middle Eocene deposits at "Main" quarry (Arzignano, Vicenza) (Busulini et al, 1982), at "Boschetto" quarry (Nogarole Vicentino, Vicenza) (Beschin et al., 1994), and many specimens referred to the taxon found at "contrada Gecchelina" (Monte di Malo, Vicenza) from Early Eocene limestones are being studied (Beschin et al, 2000).
Some species originally referred to Phlyctenodes must be placed within other genera: for P. hantkeni Lörenthey, 1898 found in Eocene levels of Hungary and Sicily, Italy, Pseudophlyctenodes gen. nov. is herein erected. Daira depressa (Milne Edwards, 1865), originally referred to the genus Phlyctenodes and commonly found in Oligocene outcrops in Véneto, Italy, is briefly discussed here.
SYSTEMATIC PALEONTOLOGY
Systematic arrangement proposed by Martin and Davis (2001) is here adopted. Measurements (width = W; length = L) are given in millimeters (mm). Specimens are deposited in the Museo civico "G. Zannato", Montecchio Maggiore, Vicenza (Italy) under acronym MCZ.
Superfamily Xanthoidea MacLeay, 1838
Family Xanthidae MacLeay, 1838
Subfamily Actaeinae Alcock, 1898
Genus Phlyctenodes Milne Edwards, 1862
Type species. Phlyctenodes tuberculosus Milne Edwards, 1862, Eocene, France.
Description. Carapace ovate, wider than long, vaulted longitudinally especially in the anterior part; dorsal surface with large tubercles, round and more or less isolated, never fused, more numerous and apparent on the anterior part of the carapace; regions not well defined. Front wide with four or more tubercle–like teeth (excluding inner–orbital spines); orbits large, subcircular, rimmed, without upper orbital fissures; frontal and anterolateral margins forming a wide, regular arch; anterolateral margins thick, with tubercles whose number and size change with the species; lateral angle underlined by a short little dorsal ridge usually tuberculate; posterolateral margins converging posteriorly, almost straight to concave, smooth and thick; posterior margin straight to a little convex, a little shorter than fronto–orbital margin. Pereiopods, sternum and abdomen unknown.
Frontal, epi– and protogastric regions fused, not distinguishable; the part of the carapace corresponding to these regions is herein indicated as "interorbital regions".
Discussion. The systematic position of the genus Phlyctenodes Milne Edwards, 1862 has always been problematic, also because its fossil remains consist only of the carapaces. Milne Edwards (1862) put it among the carpiliids and observed that no other representative of this group shows large and isolated tubercles on the dorsal part of the carapace, a feature typical of this genus. He asserted that it resembles the genera Actaea de Haan, 1833 and Actaeodes Dana, 1851, but also noted that the features of the rich ornamentation of the species of these two extant genera are different from those of Phlyctenodes: actually, the tubercles on the carapace of Actaea savignyi (H. Milne Edwards, 1834) are made up of little granules or tubercles and show a raspberry–like structure (Barnard, 1950).
Some authors have observed the resemblance between the genus Phlyctenodes Milne Edwards, 1862 and the genus Daira de Haan 1833 (Guinot, 1967; Via, 1969). Daira, initially placed within the Xanthidae MacLeay, 1838, is now referred to the family Dairidae Ng and Rodriguez, 1986, superfamily Parthenopoidea, MacLeay, 1838 (Martin and Davis, 2001). Formerly, Guinot (1978) had already placed Daira within the Parthenopoidea. Guinot (1967) thought that the genus Daira represents an advanced evolutionary stage among the parthenopoids as it has evolved to a "xanthiforme" direction and assumes an intermediate position between these two groups. She noted that it is difficult to say if Phlyctenodes has more characters of Daira than of Actaea. The genus would have differentiated from an ancestral form, the same from which Daira would descend and that would represent also the stock from which some species referred to Actaea could have arisen. From a more ancient form, one evolutionary branch originated the parthenopoids with the fossil genus Phrynolambrus Bittner, 1893 and the living one Dairoides Stebbing, 1920; a second branch, already differentiated during the Eocene, gave rise to Daira: this genus appears as a living fossil; a third branch, similar to the others and precociously differentiated (with forms like Phlyctenodes) would have produced more groups of xanthids: one with Glyptoxanthus Milne Edwards, 1879, Euxanthus Dana, 1851, Hypocolpus Rathbun, 1897, Carpoporus Stimpson, 1871 and Edwardsium Guinot, 1967 and another one with Actaea.
Even if Phlyctenodes and Daira appear very similar, some important differences can be noted; in Phlyctenodes the lateral angle is more apparent than in Daira: in this second genus the antero– and posterolateral margins are not clearly distinct and the posterolateral margins begin in the posterior quarter of the carapace; in Phlyctenodes the ornamentation is made up of tubercles that appear isolated, never fused and become less apparent on the axial and posterior regions, while in Daira there are larger and flat nodules developed also on the posterior regions; in Phlyctenodes the front is wide with four or more tubercle–like teeth whereas in Daira it is bilobed and deflected; in Phlyctenodes the dorsal regions are not well defined and a large, pentagonal mesogastric region can be observed; on the contrary, in Daira the same region is relatively narrower with an anterior process well developed and directed forward to the front.
In many studies, Phlyctenodes is placed within the Xanthidae (Glaessner, 1969; Via, 1969; Busulini et al., 1982; Miiller and Collins, 1991;Beschine et al. (1994) and, at last, it has been referred to the Carpiliidae Ortmann, 1893, again by Beschin et al. (2004).
The observation of the features of the carapace, when chelipeds and ventral parts lack, makes it difficult to establish with certainty the systematic position of this genus. Anyway, recent studies (see for example Schweitzer, 2003) have shown that some characters of the dorsal carapace can be considered as diagnostic at the family and generic level even if the ventral parts are not preserved.
Schweitzer (2003) studied in detail fossil records of the Carpiliidae and on the basis of their features clarified that they possess carapaces wider than long that may be ornamented with large and flat nodes, regions moderately to poorly defined, grooves usually not developed, front usually with bilobed median projection and blunt inner–orbital spines, orbits circular, entire, rimmed or beaded; anterolateral margins long and convex, posterolateral margins short, straight or slightly concave, posterior margin narrow, nearly straight. The analysis of all the species referred to Phlyctenodes shows that this genus cannot be placed within the carpiliids. In Phlyctenodes the front is almost straight, generally with a median notch, adorned with tubercles and the carapace shows a typical ornamentation made of tubercles above all on the anterior and lateral regions (among the carpiliids the carapace has sometimes only flat nodes as in O calina floridana Rathbun, 1929 characterized by broad, low swellings and large tubercles on the frontal and anterolateral margins).
The dorsal peculiarities of the carapace oí Phlyctenodes allow to place the genus within the heterogeneous superfamily Xanthoidea MacLeay, 1838, apparently with close relationships with the subfamily Actaeinae Alcock, 1898 (family Xanthidae MacLeay, 1838) whose representatives possess carapaces usually divided into distinct regions, granulate, tuberculate or spinous, withabilobed and spinous front (Serene, 1984). Certain affinities can be observed above all between the genus Phlyctenodes and some species referred to the genus Actaea de Haan, 1833, as some authors have pointed out (Milne Edwards, 1862; Guinot, 1967). This is true above all for those species characterized by an ornamentation constituted by isolated tubercles such as Actaea polyacantha (Heller, 1861) living in coral environments in the Red Sea, the Indian and Pacific Oceans (Serene, 1984).
The species now referred to the genus Phlyctenodes Milne Edwards, 1862 constitute a fairly homogeneous group. They are distinguishable because of the arrangement of the tubercles and the degree of definition of the regions on the dorsal carapace. In the past, several authors have thought they ought to be divided into two groups: the first one with the complete carapace covered with tubercles, the second one with the posterior and median part almost smooth. Airaghi (1905) and afterward Fabiani (1911) referred to the first group "P. depressus" Milne Edwards, 1865 and "P. hantkeni" Lörenthey, 1898 and, to the second one, P. dalpiazi Fabiani, 1911, P. krenneri Lörenthey, 1898, P. nicolisi Bittner, 1884, P. pustulosus Milne Edwards, 1862, P. steinmanni Lörenthey, 1902, and P. tuberculosus Milne Edwards, 1862. Lörenthey and Beurlen (1929) also made a similar consideration, but their subdivision didn't agree with the preceding one: in fact they put in the first group "P. depressus", "P. hantkeni", P. nicolisi andP. steinmanni and in the second one P. dalpiazi, P. krenneri and P. tuberculosus while P. pustulosus would be in an intermediate position. These attempts at subdivision are baseless: in all the species the tubercles become smaller and less apparent in the posterior part of the carapace but are always present as can be deduced from the diagnoses and the figures; moreover "P. depressus" and "P. hantkeni" are currently placed within other genera. The first one is referred to the genus Daira de Haan, 1833. A new genus, Pseudophlyctenodes gen. nov., is here erected for the second one. P. nicolisi Bittner, 1884 differs slightly from the other species; in fact, its small tubercles don't have regular arrangement: this character could bring it near to P. pustulosus. The holotype of this second species is very damaged so it is not possible a certain observation of the disposition of the tubercles; anyway, the ornamentation on the posterior regions of the carapace is almost absent and the furrows typical of P. nicolisi are not apparent.
Ristori (1896) described P. irregularis from Middle Miocene rocks of Piemonte, NW Italy, from a fragment of carapace; its assignment is very difficult but he observed that some of the irregularly arranged tubercles are fused; this would exclude the attribution of the fragment to the genus Phlyctenodes: this species, therefore, is not considered in the present analysis.
Phlyctenodes tuberculosus Milne Edwards, 1862
Figures 2.1–2.3
Phlyctenodes tuberculosus Milne Edwards, 1862, p. 193, pl. 7, fig. 2.
Description. Carapace longitudinally vaulted with hardly defined regions; frontal margin wide with four tubercle–like teeth (excluding inner–orbital spines); orbits rimmed without upper orbital fissures; anterolateral margins with five tubercles; numerous isolated tubercles on the anterior and lateral regions of the carapace, smaller on the axial ones; four longitudinally rows of four tubercles on intraorbital regions; one small tubercle on each anterolateral angle of the mesogastric region; two series of four tubercles concentric to anterolateral margins; a transverse row of little tubercles on the lateral angle constitutes a little dorsal ridge; posterior branchial and cardiac regions almost smooth.
Discussion. Phlyctenodes tuberculosus Milne Edwards, 1862 has been erected with a sole carapace specimen found nearHastingues (Landes, France) in "terrain nummulitique" (probably Middle Eocene). Comparing the figure published in 1862 and the photo of the holotype (Figure 2.1) some important differences are apparent: the posterolateral margins are less concave and there are some tubercles on the cardiac region that, according to the original description given by Milne Edwards, would be smooth. A second specimen is described here. It has been found in Late Eocene rocks at San Feliciano (Vicenza, Italy) and it is very similar to the holotype in the general shape of the carapace, even if it is smaller, and in the arrangement of the tubercles that are isolated. Clearly shown on the right anterolateral margin are the five tubercles described by Milne Edwards; because of the better preservation they project forward and are pointed.
P. tuberculosus has great affinities with P dalpiazi Fabiani, 1911 and P krenneri Lörenthey, 1898 because of the presence of four longitudinal rows of tubercles on the interorbital regions, but in the type species, the regions are not defined and the tubercles are clearly isolated and form rows that don't diverge from the axial line; moreover there is a little tubercle on each anterolateral angle of the mesogastric region.
Material examined. Holotype (cast and photographs) R03826, Museum National d'Histoire Naturelle, Paris; one specimen (MCZ 2456 W 13.6 L 9.2) from San Feliciano (Colli Berici, Vicenza).
Occurrence. Middle Eocene: SW France; Late Eocene: Véneto, NE Italy.
Phlyctenodes dalpiazi Fabiani, 1911
Figures 2.4–2.6
Phlyctenodes dalpiazi Fabiani, 1911, p. 4, fig. 1.
Description. Carapace vaulted in the anterior half, axial regions fairly defined. Frontal margin with four tubercles (excluding inner–orbital spines), orbits rimmed, with sparse and little granules; anterolateral margins tuberculate; posterolateral margins a little concave; posterior margin a little wider than frontal one, straight, rimmed. Tubercles present on anterior and lateral regions. Interorbital regions with four longitudinal rows, diverging posteriorly, each composed of four, contiguous, broad, blunt tubercles. Irregularly arranged tubercles on the lateral regions. Mesogastric region pentagonal, fairly defined; cardiac and posterior branchial regions almost smooth.
Discussion. Phlyctenodes dalpiazi Fabiani, 1911 has been erected with only one specimen collected in limestone levels at San Feliciano (Vicenza, Italy) that Fabiani attributed to the Early Oligocene. More recent studies show them to be Late Eocene (Ungaro and Bosellini, 1965; Ungaro, 1978; De Angeli and Garassino, 2002). Fabiani (1911) described also the propodus of a small claw preserved on the same rocky fragment and suggested that it could be a part of the same individual: the features of this propodus make his hypothesis unlikely.
Some specimens referred to the species are housed in the Museo civico "G. Zannato" (Montecchio Maggiore, Vicenza): two of them have been found in the type locality (San Feliciano) and one in coeval levels at Campolongo di San Germano (Vicenza). They preserve the frontal margin, lacking in the holotype: it has four small tubercles. It is possible to confirm the differences between P. dalpiazi and P. krenneri Lörenthey, 1898 regarding the lack of well defined tubercles on the mesogastric region (and usually on the cardiac one) and the features of the tubercles (Fabiani, 1911).
Material examined. Holotype, MGPD 23654, Museo Geológico e Paleontológico, Universitá di Padova; two specimens (MCZ 2458, L 7.0; MCZ 2459, W 9.1, L 7.0) from San Feliciano (Vicenza) and one (MCZ 2457, W 11.8, L 8.2) from Campolongo di San Germano (Vicenza).
Occurrence. Late Eocene: Véneto, NE Italy.
Phlyctenodes krenneri Lörenthey, 1898
Figures 2.7–2.10
Phlyctenodes krenneri Lörenthey, 1898, p. 46, pl. 2, fig. 9.
Phlyctenodes krenneri; Checchia–Rispoli, 1905, p. 312, pl. 1, fig. 10.
Phlyctenodes krenneri; Lörenthey and Beurlen, 1929, p. 201, pl. 12, fig. 9.
Phlyctenodes krenneri; Di Salvo, 1933, p. 20.
Phlyctenodes krenneri; Miiller and Collins, 1991, p. 76, pl. 5, fig. 9, pl. 6, fig. 1.
Description. Carapace longitudinally vaulted with poorly defined regions. Front wide with four small tubercles (excluding inner–orbital spines); anterolateral margins tuberculate; posterolateral margins a little curved; posterior margin straight, rimmed. Furrows between gastric and hepatic regions. Surface covered with pointed tubercles on anterior regions; four longitudinal rows of tubercles on interorbital regions; four small tubercles in transverse row on pentagonal mesogastric region; three small tubercles constituting a triangle on wide, transversely ovate cardiac region; series of large tubercles on the hepatic and anterior branchial regions concentric to anterolateral margins. Posterior regions almost smooth.
Discussion. Phlyctenodes krenneri differs from P. dalpiazi in the constant presence of four tubercles in a transverse row on the mesogastric region, with three others forming a triangle on the cardiac region and in the features of the tubercles on the dorsal surface that appear definitely pointed and isolated (Fabiani, 1911). The species has been found also in Sicily: Checchia–Rispoli (1905) reported a damaged specimen (Figure 2.8) and later Di Salvo (1933) reported six specimens: examination of the photographs of these specimens housed in the Museo geológico "G.G. Gemmellaro" (Palermo) allows to confirm their attribution and also to validate the persistence of the typical features of the species with a carapace varying in size; the length ranging from 2.5 to 10.0 and the width respectively from 4.0 to 13.2 (Di Salvo, 1933).
Material examined. Holotype (cast), Magyar Állami Fóldtani Intézet, Budapest; photographs of specimens BAGA006, BAGA015, BICH2, PILU002, PILU004, Museo Geológico "G.G. Gemmellaro", Palermo (Italy); one specimen (MCZ 2460, L 9.3) not well preserved from Campolongo di San Germano (Vicenza, Italy).
Occurrence. Middle Eocene: Sicily, Italy; Late Eocene: Hungary, Véneto (NE Italy).
Phlyctenodes nicolisi Bittner, 1884
Figures 3.1, 3.2
Phlyctenodes nicolisi Bittner, 1884, p. 5, pl. 1, fig. 5.
Description. Carapace ovate, vaulted with fairly evident axial regions; frontal margin wide, tuberculate, with a median notch; orbital margins inflated and beaded; anterolateral margins with seven tubercles (including outer–orbital angle); posterolateral margins concave, very thick, smooth; posterior margin almost straight; branchiocardiac furrows clear, reaching the posterior margin. Dorsal surface covered with many small, round, irregularly arranged tubercles; they are larger and forward directed near anterior margins; smaller on the posterior regions. Cardiac region with three tuberculate swellings forming a triangle.
Discussion. Only the holotype of this species is known: it was collected near Avesa (Verona). It preserves the orbito–antennal region, described by Bittner (1884); it is possible to observe a fairly wide, transverse basal segment of antennula and a narrow antennal fissure on the infraorbital margin.
One specimen reported in Beschin et al. (2000) from the Early Eocene of "contrada Gecchelina" (Monte di Malo, Vicenza) has been formerly attributed to the species but it is under study and its assignement is not certain yet.
Material examined. Holotype 1982, Museo Civico di Storia Naturale di Verona, Italy.
Occurrence. Middle Eocene: Véneto, NE Italy.
Phlyctenodes pustulosas Milne Edwards, 1862
Figures 3.3, 3.4
Phlyctenodes pustulosus Milne Edwards, 1862, p. 194, pl. 7, fig. 1.
Description. Carapace ovate, regions hardly distinguishable; axial furrow separating protogastric regions; branchiocardiac furrows visible. Frontal margin wide with four (or six) tubercles; six tubercles increasing in size toward lateral angle. Many isolated tubercles on anterior and lateral regions, smaller in posterior regions; posterior part of carapace nearly smooth; tubercles constituting a row concentric to anterolateral margins.
Discussion. The holotype, and sole specimen found at Nousse (Landes, France), is a carapace emerging from a marly limestone. The comparison of the specimen to the original figure suggests the specimen has been probably damaged after the publication and some of the characters described by Milne Edwards are not observed.
Material examined. Holotype (cast and photographs) A24551, Museum National d'Histoire Naturelle, Paris.
Occurence. Middle Eocene: SW France.
Phlyctenodes steinmanni Lörenthey, 1902
Figure 3.5 – 3.7
Phlyctenodes steinmanni Lörenthey, 1902, p. 111, pl. 1, fig. 4.
Phlyctenodes steinmanni; Lörenthey and Beurlen, 1929, p. 200, pl. 12, fig. 2.
Phlyctenodes steinmanni; Miiller and Collins, 1991, p. 76, pl. 5, fig. 9, pl. 6, fig. 1.
Phlyctenodes steinmanni; Beschin, Busulini, De Angeli and Tessier, 1994, p. 188, pl. 9, fig. 3.
Description. Carapace ovate with well defined regions; frontal margin with four spines; orbital and anterolateral margins spinous; posterolateral margins straight, smooth; posterior margin rimmed. Median furrows present on both front and protogastric regions; branchiocardiac furrows visible. Dorsal surface covered with pointed tubercles larger on the anterior regions. Three longitudinal rows of tubercles on each protogastric region; longitudinal rows of tubercles on branchial regions; mesogastric regions with four rows of small tubercles; cardiac region triangular with very small, irregularly arranged tubercles; metabranchial regions smooth, anteriorly delimited by a transverse row of small tubercles forming a dorsal ridge; intestinal region with smooth, transverse, slightly concave ridge.
Discussion. Specimens referred to this species have been found frequently, but the individuals mentioned herein allow a complete description of the posterior part of the carapace, damaged in the holotype; the presence of a transverse ridge on the intestinal region is significant.
Material examined. Holotype (cast) Magyar Állami Fóldtani Intézet, Budapest; two specimens (MCZ 2461, W 13.6, L 11.0; MCZ 2465, W 11.7, L 8.4) from Campolongo di San Germano (Vicenza, Italy) and two (MCZ 2462, W 11.5, L 8.0; MCZ 2466) from San Feliciano (Vicenza).
Occurrence. Middle Eocene: Véneto, Italy; Late Eocene: Hungary and Véneto (NE Italy).
Genus Pseudophlyctenodes gen. nov.
Type species. Phlyctenodes hantkeni Lörenthey, 1898, Eocene, Hungary and Italy.
Etymology. Pseudophlyctenodes (m.) from 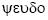 (gr.) = false, and the similar genus Phlyctenodes.
(gr.) = false, and the similar genus Phlyctenodes.
Diagnosis. As for type species.
Pseudophlyctenodes hantkeni (Lörenthey, 1898)
comb. nov.
Figures 3.9, 3.10
Phlyctenodes hantkeni Lörenthey, 1898, p. 44, pl. 2, fig. 10.
Phlyctenodes hantkeni; Lörenthey and Beurlen, 1929, p. 199, pl. 12, fig. 8.
Phlyctenodes hantkeni; Di Salvo, 1933, p. 21, pl. 2, fig. 1.
Description. Carapace ovate, wider than long, strongly longitudinally vaulted with fairly defined regions. Frontal and anterolateral margins forming a regular arch. Frontal margin wide. Orbits large, orbital margins poorly swelled, tuberculate. Anterolateral margins tuberculate; posterolateral margins strongly concave, very thick, almost smooth; posterior margin straight, rimmed. Furrows dividing gastric and cardiac regions from fused hepatic and branchial ones. Mesogastric region pentagonal. Protogastric regions large, separated by an axial furrow and reaching frontal margin. Dorsal surface covered with close, large, composite tubercles. Three longitudinal rows of composite tubercles on each protogastric region. Rows of tubercles on the hepatic and branchial regions diverging like rays from mesogastric region. Tubercles smaller, irregularly arranged on axial and metabranchial regions.
Discussion. This species, characterized by a very close, composite tuberculation, appears to be in an intermediate position between the representatives of the genus Phlyctenodes and those of Daira. Pseudophlyctenodes hantkeni (Lörenthey, 1898) is very similar to Daira, as Lörenthey and Beurlen (1929) have suggested, in the general carapace shape with antero– and posterolateral margins almost continuous. Definitely different, and near to Phlyctenodes, is the mesogastric region proportionally wider without a developed anterior process; the protogastric regions are separated only by an axial furrow. On the other hand, the species shows a rich ornamentation made up of assembled tubercles, very different from the individual tubercles typical of Phlyctenodes. The assignement to the new genus Pseudophlyctenodes gen. nov. allows to give this species a more adequate systematic position.
Material examined. Casts of type series, Magyar Állami Fóldtani Intézet, Budapest; photographs of specimens BAGA001, BAGA003, BAGA004, BAGA005, PILU003 Museo Geológico "G.G. Gemmellaro", Palermo (Italy).
Occurrence. Middle Eocene: Sicily, Italy; Late Eocene: Hungary.
Superfamily Parthenopoidea MacLeay, 1838
Family Dairidae Ng and Rodriguez, 1986
Genus Daira de Haan, 1833
Type species. Cancer perlatus Herbst, 1790, Recent, Indo–Pacific.
Daira depressa (Milne Edwards, 1865)
Figure 3.8
Phlyctenodes depressus Milne Edwards, 1865, p. 367, pl. 33, fig. 2.
Phlyctenodes depressus; Bittner, 1877, p. 446.
Phlyctenodes depressusv Bittner, 1883, p. 15.
Phlyctenodes depressus; Airaghi, 1905, p. 205, pl. 4, fig. 3.
Daira depressa; Glaessner, 1929, p. 135.
Daira depressa; Beschin et al, 2001, p. 20, pl. 2. figs. 2, 4.
Material examined. Two specimens (MCZ 2463, L 16.1, MCZ 2464) from Montecchio Maggiore (Vicenza, Italy).
Description. Carapace ovate, wider than long, almost flattened; orbits small; frontal margin tuberculate; frontal and anterolateral margins forming a wide arch; antero– and posterolateral margins not clearly distinct one another; posterolateral margins shorter than anterolateral ones and concave; regions moderately defined; dorsal surface covered with large tubercles very close one another, the largest on the anterior regions; tubercles longitudinally arranged on gastric regions: two longitudinal rows oneachprotogastric regions and an axial one on long, narrow anterior mesogastric process; mesogastric region pentagonal, relatively narrow; tubercles arranged in transverse rows on posterior regions of the carapace.
Discussion. The general shape of the carapace with anterolateral margins not clearly distinct from the posterolateral ones, the structure and the arrangement of the tubercles, and the mesogastric region relatively narrow with the anterior process very apparent, allow to attribute this species to the genus Daira as Bittner (1877, 1883) has already suggested. Particular affinities can be noted between Daira depressa and Daira speciosa (Reuss, 1871) of the Miocene of Europe. Daira depressa has been found frequently in Early Oligocene rocks of Véneto (see Beschin et al., 2001, for a detailed analysis).
Occurrence. Early Oligocene: Véneto, NE Italy.
ACKNOWLEDGEMENTS
We wish to thank R. Ghiotto (Museo Civico "G. Zannato", Montecchio Maggiore, Vicenza) for providing the study material housed in his museum; M. Fornasiero (Museo Geológico e Paleontológico, Universitá di Padova) for providing access to the type originally described by Fabiani at her institution, and S. Castelli for the photographs of the same specimen; A. Vaccari (Museo Civico di Storia Naturale, Verona) for providing access to the type originally described by Bittner at her institution and the photographs of that specimen; L. Galletti (Museo Geológico "G.G.Gemmellaro", Universitá di Palermo) for providing photographs of some specimens originally studied by Checchia–Rispoli and Di Salvo; J.M. Pacaud (Museum National d'Histoire Naturelle, Paris) for providing casts of the holotypes originally studied by Milne Edwards. We are very grateful to D. Guinot (Museum National d'Histoire Naturelle, Paris) for her kind advice and to P. Miiller (Magyar Állami Fóldtani Intézet, Budapest) for casts and information concerning Hungarian specimens studied by Lörenthey.
Many thanks to F. Vega Vera (Instituto de Geología, UNAM) for his kind and thoughtful review of our manuscript and to R.H.B. Fraaije (Oertijdmuseum, Boxtel), G.A. Bishop (Museum of Geology, South Dakota) and H. Karasawa (Mizunami Fossil Museum) for their critical reviews. Thanks also to A. Checchi (Associazione Amici del Museo "G. Zannato") for his field work and to A. De Angeli (Associazione Amici del Museo "G. Zannato") for providing photographs of the holotypes studied by A. Milne Edwards. For their patient "linguistic assistance" thanks to I. Bozza and M. Busulini and to F. Turio for his photographic help.
REFERENCES
Airaghi, C, 1905, Brachiuri nuovi o poco noti del Terziario véneto: Atti Societá Italiana di Scienze Naturali, 44, 202–209. [ Links ]
Alcock, A., 1898, Materials for a carcinological fauna of India, No. 3, The Brachyura Cyclometopa, Part I, The Family Xanthidae: Journal of the Asiatic Society of Bengal, 67, part 2(1), 67–233. [ Links ]
Barnard, H.K., 1950, Descriptive Catalogue of South African Decapod Crustacea (Crabs and Shrims): Annals of the South African Museum, 38, 864 p. [ Links ]
Beschin, C, Busulini, A., De Angeli, A., Tessier, G., 1994, I Crostacei eocenici della cava "Boschetto" di Nogarole Vicentino (Vicenza–Italia Settentrionale): Societá Veneziana di Scienze Naturali, Lavori, 19, 159–215. [ Links ]
Beschin, C, Busulini, A., De Angeli, A., Tessier, G., Ungaro, S., 2000, The fauna of the Gecchelina quarry at Monte di Malo (Vicenza–Northern Italy): a preliminary study, in 1st Workshop on Mesozoic and Tertiary decapod crustaceans: Montecchio Maggiore, Vicenza, Museo Civico "G. Zannato", Associazione Amici del Museo, Studi e Ricerche, 7–10. [ Links ]
Beschin, C, De Angeli, A., Checchi, A., 2001, Crostacei decapodi associati a coralli della <<Formazione di Castelgomberto>> (Oligocene) (Vicenza – Italia settentrionale): Montecchio Maggiore, Vicenza, Museo Civico "G. Zannato", Associazione Amici del Museo, Studi e Ricerche, 13–30. [ Links ]
Beschin, C, Busulini, A., De Angeli, A., Tessier, G., 2004, The Eocene Decapod Crustacean fauna of the "Main" quarry in Arzignano (Vicenza – NE Italy) with the description of a new species of Raninidae: Societá Veneziana di Scienze Naturali, Lavori, 29, 109–117. [ Links ]
Bittner, A., 1877, Über Phymatocarcinus speciosus Reuss: Sitzungsberichten der Kaiserlichen Akademie der Wissenschaften in Wien, 75, 435–447. [ Links ]
Bittner, A., 1883, Neue Beitrage zur Kenntniss der Brachyuren–Fauna des Alttertiars von Vicenzaund Verona: Denkschriften der Kaiserlichen Akademie der Wissenschaften in Wien, 46, 299–316. [ Links ]
Bittner, A., 1884, Beitrage zur Kenntniss tertiarer Brachyuren–Fauna: Denkschriften der Kaiserlichen Akademie der Wissenschaften in Wien, 48, 15–30. [ Links ]
Bittner, A., 1893, Decapoden des pannonischen Tertiars: Sitzungsberichten der Kaiserlichen Akademie der Wissenschaften in Wien, I Abth., 102, 10–37. [ Links ]
Busulini, A., Tessier, G., Visentin, M., 1982, Brachyura della Cava Main (Arzignano) – Lessini orientali (Vicenza) (Crustacea, Decapoda): Societá Veneziana di Scienze Naturali, Lavori, 7, 75–84. [ Links ]
Checchia–Rispoli, G., 1905, I Crostacei dell'Eocene dei dintorni di Monreale in Provincia di Palermo: Palermo, Giornale di Scienze Naturali ed Economiche, 25, 309–325. [ Links ]
Dana, J.D., 1851, On the classification of the Cancroidea: Silliman's American Journal of Science and Arts, series 2, 12, 121–131. [ Links ]
De Angeli, A., Garassino, A., 2002, Galatheid, chirostylid and porcellanid decapods (Crustacea, Decapoda, Anomura) from the Eocene and Oligocene of Vicenza (N Italy): Memorie della Societá Italiana di Scienze Naturali e del Museo Civico di Storia Naturale di Milano, 30(3), 40 p. [ Links ]
Di Salvo, G., 1933, I Crostacei del Terziario inferiore della provincia di Palermo: Palermo, Giornale di Scienze Naturali ed Economiche, 37, 44 p. [ Links ]
Fabiani, R., 1911, Di una nuova specie di Phyctenodes (Phi. Dalpiazi) dell'Oligocene dei Berici: Bollettino Museo Civico di Vicenza, 1(3) (1910), 3–6. [ Links ]
Glaessner, M.F., 1929, Crustacea Decapoda, in Pompeckji, F.J. (ed.), Fossilium Catalogus 1, Animalia 41: Berlin, W. Junk, 464 p. [ Links ]
Glaessner, M.F., 1969, Decapoda, in Moore, R.C. (ed.), Treatise on Invertebrate Paleontology, Part R, Arthropoda 4: Boulder and Lawrence, The Geological Society of America, University of Kansas Press, 2, 400–533, 626–628. [ Links ]
Guinot, D., 1967, Recherches préliminaires sur les groupements naturels chez les Crustacés Décapodes Brachyoures, III. A propos de affinités des genres Dairoides Stebbing et Daira de Haan: Bulletin du Museum National d'Histoire Naturelle, Paris, Serie 2, 39(3), 540–563. [ Links ]
Guinot, D., 1978, Principes d'une classification evolutive des Crustacés Décapodes Brachyoures: Bulletin Biologique de la France et de la Belgique, 112(3), 211–292. [ Links ]
Haan de, W., 1833–1850, Crustacea, in von Siebold, P.F. (ed.), Fauna japonica sive descriptio animalium, quae in itinere per Japoniam, jussu et auspiciis superiorum, qui summum in India Batava Imperium tenent, suscepto annis 1823–1830 collegit, notis, observationibus et adumbrationibus illustravit: Lugduni–Batavorum (Leiden), The Netherlands, 243 p. [ Links ]
Heller, C, 1861, Synopsis der im rothen Meere vorkommenden Crustaceen: Vehandlungen der Zoologisch–botanischen Gesellschaft in Wien, 11,3–32. [ Links ]
Herbst, J.F.W., 1782–1804, Versucheiner Naturgeschichte der Krabbenund Krebse, nebst einer systematischen Beschreibung ihrer verschie–denenArten: Berlin und Stralsund, G.A. Lange, 515 p. [ Links ]
Lörenthey, I.E., 1898, Beitrage zur Decapodenfauna des Ungarischen Tertiars: Természetrajzi Füzetek, 21, 133 p. [ Links ]
Lörenthey, I.E., 1902, Neuere Beitrage zur Tertiaren Decapodenfauna Ungarns: Mathematische und Naturwissenschaftliche Berichte aus Ungarn, 18,98–120. [ Links ]
Lörenthey, I.E., Beurlen, K., 1929, Die fossilen Decapoden der Lander der Ungarischen Krone: Geológica Hungarica, series Palaeontologica, 3, 420 p. [ Links ]
MacLeay, W.S., 1838, Illustrations of the Annulosa of South Africa; On the brachyurous decapod Crustacea brought from the Cape by Dr. Smith, in Smith, A. (ed.), Illustrations of the Zoology of South Africa, Invertebrate: London, Smith, Elder and Company, 53–71. [ Links ]
Martin, J.W., Davis, G.E., 2001, An updated classification of the Recent Crustacea: Natural History Museum of Los Angeles County, Science, series 39, 129 p. [ Links ]
Milne Edwards, A., 1861–1865, Histoire des Crustacés Podophthalmaires fossiles: Paris, Victor Masson et fils, 390 p. [ Links ]
Milne Edwards, A., 1862, Monographie des Crustacés fossiles de lafamille des Cancériens: Ármales des Sciences Naturelles, Zoologie, serie 4, 18,31–85. [ Links ]
Milne Edwards, H., 1834, Histoire naturelle des Crustacés comprenant l'anatomie, la physiologie et la classification de ces animaux; Atlas: Paris, Librairie Encyclopédique de Roret, 1, 468 p. [ Links ]
Müller, P., Collins, J.S.H.,1991, Late Eocene coral–associated decapods (Crustacea) from Hungary: Contributions to Tertiary and Quaternary Geology, 28 (2–3), 47–92. [ Links ]
Ng, P.K.L., Rodriguez, G., 1986, New records of Mimilambrus wileyi Williams, 1979 (Crustacea: Decapoda. Brachyura), with notes on the systematics of the Mimilambridae Williams, 1979, and Parthenopidae MacLeay, 1838, sensu Guinot, 1978: Proceedings of the Biological Society of Washington, 99(1), 88–99. [ Links ]
Ortmann, A., 1893, Die Dekapoden–Krebse des Strassburger Museums, 7. Theil, Abtheilung Brachyura (Brachuyra genuina Boas), 2. Unterabtheilung Cancroidea, 2. Section Cancrinea, 1. Gruppe, Cyclometopa: Zoologischen Jahrbiicher, Abtheilung fiir Systematik, Geographie und Biologie der Thiere, 7, 411–495. [ Links ]
Rathbun, M.J., 1897, A revision of the nomenclature of the Brachyura: Proceedings of the Biological Society of Washington, 11, 153–167. [ Links ]
Rathbun, M.J., 1929, Anew crab from the Eocene of Florida: Proceedings of the United States National Museum, Bulletin 75, Article 15, 4 p. [ Links ]
Reuss, A., 1871, Phymatocarcinus speciosus, eine neue fossile Krabbe aus dem Leithakalk des Wiener Beckens: Sitzungsberichten der Kaiserlichen Akademie der Wissenschaften in Wien, 63, 325–330. [ Links ]
Ristori, G., 1896, Crostaceineogenici di Sardegnae di alcune altre localitá italiane: Bollettino Societá Geológica Italiana, 15, 504–513. [ Links ]
Schweitzer, C.E., 2003, Utility of proxy characters for classification of fossils: an example from the fossil Xanthoidea (Crustacea: Decapoda: Brachyura): Journal of Paleontology, 77(6), 1107–1128. [ Links ]
Serene, R., 1984, Crustacés Décapodes Brachyoures de l'Ocean Indian Occidental et de la Mer Rouge; Xanthoidea: Xanthidae et Trapeziidae, Avec un addendum par Croisner, A.: Carpiliidae et Menippidae: Faune tropicale, 24, 400 p. [ Links ]
Stebbing, T.R.R., 1920, South African Crustacea (Part X of S. A. Crustacea, for the Marine Investigations in South Africa): Annals of the South African Museum, 17(4), 231–272. [ Links ]
Stimpson, W., 1871, Preliminary report on the Crustacea dredged in the Gulf Stream in the Straits of Florida by L.F. de Pourtales, assistant United States Coast Survey, Part I. Brachyura: Bulletin of the Museum of Comparative Zoology at Harvard College, 2, 109–160. [ Links ]
Ungaro, S., 1978, L'Oligocene dei Colli Berici: Rivista Italiana di Paleontología, 84(1), 199–278. [ Links ]
Ungaro, S., Bosellini, A., 1965, Studio micropaleontologico e stratigrafico sul limite Eocene–Oligocene nei Colli Berici occidentali: Annali Universita di Ferrara (nuova serie), sezione 9, Scienze Geologiche e Mineralogiche, 3(9), 157–183. [ Links ]
Via, L., 1969, Crustáceos Decápodos del Eoceno español: Pirineos, 91–94, 479 p. [ Links ]

