










Servicios Personalizados
Revista

Articulo
 Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
 Citado por SciELO
Citado por SciELO Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista mexicana de ciencias geológicas
versión On-line ISSN 2007-2902versión impresa ISSN 1026-8774
Rev. mex. cienc. geol vol.23 no.3 Ciudad de México ene. 2006
Sección especial
El Espinal, a new plattenkalk facies locality from the Lower Cretaceous Sierra Madre Formation, Chiapas, southeastern Mexico
Francisco J. Vega1, Pedro García–Barrera3, María del Carmen Perrilliat2, Marco A. Coutiño4, and Ricardo Marino–Pérez3
1 Instituto de Geología, Universidad Nacional Autónoma de México, Ciudad Universitaria, Delegación Coyoacán, 04510 México D.F., Mexico. vegver@servidor.unam.mx
2 Instituto de Geología, Universidad Nacional Autónoma de México, Ciudad Universitaria, Delegación Coyoacán, 04510 México D.F., Mexico.
3 Facultad de Ciencias, Universidad Nacional Autónoma de México, Ciudad Universitaria, Delegación Coyoacán, 04510 México D.F., Mexico.
4 Museo de Paleontología Eliseo Palacios Aguilera , Instituto de Historia Natural y Ecología de Chiapas, Calzada de los Hombres Ilustres s/n, Parque Madero, 29000 Tuxtla Gutiérrez, Chiapas, Mexico.
Manuscript received: February 8, 2006
Corrected manuscript received: April 14, 2006
Manuscript accepted: May 20, 2006
ABSTRACT
A new plattenkalk facies locality with plants, invertebrates and vertebrates in laminar dolomitic limestones from the Albian of the lower part of the Sierra Madre Formation in central Chiapas is reported. The gastropod ?Perissoptera sp., one odonate–zygopteran nymph, one hemipteran–?belostomatid adult, a new species of lobster, Palinurus palaciosi (first report for the genus as a fossil in America), and the paguroid Roemerus robustus Bishop, 1983 are described. Plant remains as leaves and a cone are also present. An Albian age is suggested on the basis of stratigraphic position and some fauna. On the basis of palynological analysis and lithofacies, thepaleoenvironment is interpreted as a brackishwater shallow lagoon with occasional freshwater influence.
Key words: plattenkalk, Cretaceous, Albian, Sierra Madre Formation, Chiapas, Mexico.
RESUMEN
Se reporta una nueva localidad de facies plattenkalk con plantas, invertebrados y vertebrados en dolomitas laminares de la parte inferior de la Formación Sierra Madre (Albiano), en la región central de Chiapas. Se describe el gasterópodo ?Perissoptera sp., una ninfa de odonato–zigóptero, un adulto hemíptero–?belostomátido, una nueva especie de langosta Palinurus palaciosi (primer registro fósil del género en América), y el paguroide Roemerus robustus Bishop, 1983. También están presentes restos de plantas como hojas y un cono. Se sugiere una edad albiana con base en la posición estratigráfica y el alcance bioestratigráfico de algunas especies. Con base en análisis palinológico y litofacies, el paleoambiente es interpretado como una laguna salobre, con influencia ocasional de agua dulce.
Palabras clave: plattenkalk, Cretácico, Albiano, Formación Sierra Madre, Chiapas, México.
INTRODUCTION
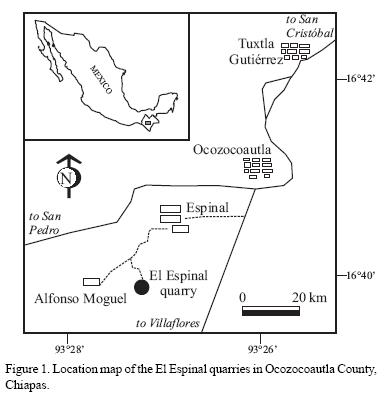
El Espinal quarry is located approximately 45 km west of Tuxtla Gutiérrez, Chiapas, at Long 93°26'97" W and Lat 16°41' 1" N (Figure 1). A series of shallow excavations exploited by locals have yielded plant remains, fishes, mollusks, crustaceans and coprolites as the most conspicuous fossils found on finely laminated clay layers interbedded with dolomitic limestone. Similar fossils have been found at the Tlayua Formation lithographic limestones of Albian age in Puebla, southern Mexico, as fishes, one odonate nymph and isopods (Applegate, 1988, 1992, 1995; Applegate et al., 2006; Feldmann et al., 1998). There are practically no formal reports on any fossil from El Espinal quarry, except for two studies on fishes by González et al. (2002) and Ovalles–Damián (2004). To the moment, the most abundant macrofossils are crustaceans, particularly tanaidaceans, which are found in discrete horizons of thin orange clay, interbedded with relatively thick layers of cream limestone that range from 5 to 10 cm. Numerous tanaidaceans and two species of Isopoda (Sphaeromatidae and Cirolanidae) are also present on the clay layers, as well as an odonate–zy–gopteran nymph, one hemipteran–?belostomatid, the lobster Palinuruspalaciosi new species, and the pagwoid Roemenis robustas Bishop, 1983. Other macrofossils include gastropods (?Perissoptera sp. and cerithids), bivalves, echinoid plates, ichnites, and diverse plant remains. No microfossils are present, except unidentified ostracods.
Stratigraphy
The studied rocks correspond to the lower part of the Sierra Madre Formation, formally described by Gutiérrez–Gil (1956), but previously reported by other authors (Bose, 1905; Ver Wiebe, 1925; Müllerried, 1936; Imlay, 1944). The formation was studied in more detail and subdivided in several members (Chubb, 1959; Sánchez–Montes de Oca, 1969; Zavala–Moreno, 1971; Castro–Mora et al., 1975; Michaud, 1987; Quezada–Muñetón, 1987).
A composed thickness of 2,590 m was suggested by Steele and Waite (1986) for the Sierra Madre Formation (Figure 2), subdivided into 21 lithofacies. The lowermost lithofacies corresponds to the stratigraphic level of El Espinal quarry, defined by Steele and Waite (1986) as dolomite and dolomitic breccia, located between 650 and 700 m from the base of the formation, which lies conformably over the Upper Jurassic–Lower Cretaceous San Ricardo Formation. The top is covered unconformably by the Campanian–Maastrichtian Ocozocoautla Formation.
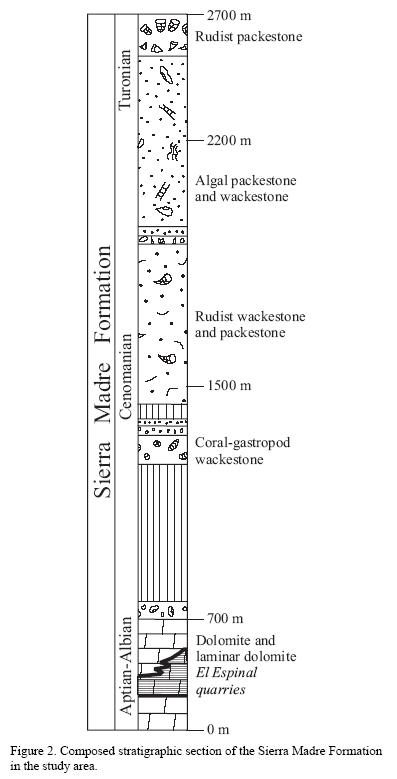
Based on the rudists Radiolites sp., Caprina sp., and Toucasia sp., Gutiérrez–Gil (1956) suggested a Lower to "middle" Cretaceous age for the entire Sierra Madre Formation. Sánchez–Montes de Oca (1969) assigned to this formation an age of upper Albian to Cenomanian, based on presence of the benthic foraminifera Dicyclina schlumbergeri and Nummoloculina heimi. Steele and Waite (1986) indicated an age of upper Aptian to Santonian for the entire Sierra Madre Formation, and defined an age of Neocomian?–lower Albian for their basal unit (dolomite), which corresponds to the El Espinal stratigraphic level. Lithologic unit two (stratigraphically above El Espinal beds) of Steele and Waite (1986, p. 141) contains Nummoloculina heimi (Albian–Cenomanian of Mexico, Texas and Florida), as well as Simplorbitolina sp. and Coskinolinoides sp., orbitolinid foraminifera of Aptian–Albian age. Michaud (1987) measured a section nearby El Espinal, from which he reported Praechrysalidina sp. cf. P. infracretacea from the upper Aptian–lower Albian. Presence of the hermit crab Roemerus robustas Bishop, 1983, first reported from the early Albian of Texas, reinforces an Albian age interpretation for the El Espinal beds.
Some beds at El Espinal quarry contain cerithid and aporrhaid gastropods, similar to species reported for the Aptian–Albianbeds of Europe. Anew genus of paraclupeid fish was described from El Espinal by Ovalles–Damián (2004), who discussed affinities of this new form with fishes described from Aptian–Albian localities in South America and China.
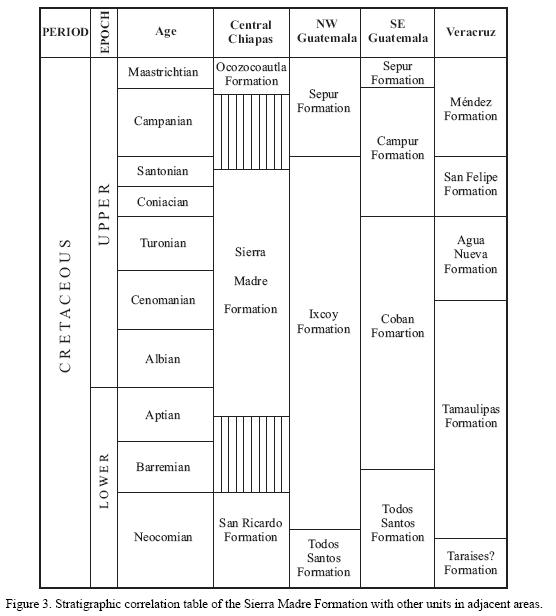
At a regional scale, the Sierra Madre Formation is thus correlated with the upper portion of the Ixcoy and Coban formations, and with the lower part of the Campur Formation, all from Guatemala (Figure 3).
Paleoenvironment
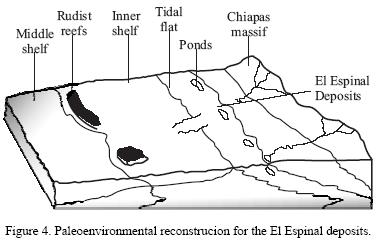
Palynological analysis of fossiliferous layers at El Espinal produced 90 – 97% of kerogen, which suggests an environment with high oxygen concentration and a high primary productivity. Spores of Sphagnum (peat moss), Selaginella and Osmunda (both ferns) are present, as well as palynomorphs such as Classopolis, Pseudoplicapollis, Arecipites, Retinomocolpites, Normapolles, Ephedra, and from the families Bombacaceae, Lauraceae, Osmundaceae, Ulmacaceae, Lilidae and Moraceae. Fungospores, acritarcs, dinoflagellates, plant cuticle and tracheids are also present. These results show a mixture of marine elements, such as acritarcs and dinoflagellates with continental forms. This mixture is also observed on the macroscopic assemblage, with typical marine forms such as pycnodont and macrose–mid fishes, tanaidaceans, isopods, lobster, pagurid, marine gastropods and echinoid remains, together with continental elements as the odonate nymph and diverse plant remains, including leaves and a gymnosperm cone.
Some layers show ripples, dessication marks, algal mats, and flat–pebble conglomerates where a few isopods and groups of ostracods can be found in between the rip–up clasts. This suggests sporadic influence of strong waves and/or currents. It is thus suggested that sediments from El Espinal were deposited in a brackish marginal environment (Figure 4). This is consistent with the interpretation given by Steele and Waite (1986), whose biofacies F was defined as a supratidal environment, where they reported ostracods, gastropod impressions and pellets in fine grain dolomite. The diversity and type of organisms found at the El Espinal Quarry (Figures 5.1–5.6) supports an interpretation of a shallow lagoon or estuary for these deposits.
Specimens are deposited in the Museo de Paleontología "Eliseo Palacios Aguilera", Instituto de Historia Natural y Ecología de Chiapas, Calzada de Los Hombres Ilustres s/n, Parque Madero, Tuxtla Gutiérrez 29000, Chiapas, and registered under numbers with acronym IHNFG.
SYSTEMATIC PALEONTOLOGY
Class Gastropoda Cuvier, 1797
Order Caenogastropoda Cox, 1959
Superfamily Stromboidea Rafinesque, 1815
Family Aporrhaidae da Costa, 1778
Genus Perissoptera Tate, 1865
Type species. Rostellariaparkinsoni Mantell, 1822, by subsequent designation of Cossmann (1904). Albian. Europe.
? Perissoptera sp.
Figures 5.7, 5.8
Description. Shell small to medium sized, medium high spired, fusiform; protoconch not preserved; teleoconch of six convex whorls. Ornamentation of axial ribs on whorls. Relatively long straight rostrum; with one short digitation connected to the spire, no posterior digitation connected to spire; ear–like extension is very small.
Material examined. Four specimens, hypotypes IHNFG–2835 to IHNFG–2838.
Occurrence. El Espinal quarry, Sierra Madre Formation, Chiapas, Albian.
Measurements. Hypotype IHNFG–2835, height: 24.9 mm, diametenll.O mm; hypotype IHNFG–2838, height: 22.8 mm, diameter: 12.8 mm.
Discussion. Owing to the relatively poor preservation of the specimens, they are assigned with doubt to Perissoptera. Perissoptera has been discussed and type species clarified by Saul (1998, p. 124), who literally stated "Perissoptera has been widely used for moderately high spired aporrhaids having the axial sculpture much stronger than the spiral, and an expanded outer lip that is drawn out at its posterior extremity into a blade set off from the lobate medial distal margin by a sinus. The type species has a relatively short and wide rostrum and is moderately carínate on the last whorl. Included within Perissoptera are species with a longer rostrum and stronger spiral sculpture than are in Arroghes, as well as a distinctive outer lip drawn out posteriorly and channeled interiorly.
Perissoptera differs from Latíala in usually having a strong spiral sculpture and in having the outer lip sinused on its distal margin, just anterior to the posterior extension, which is channeled interiorly.
Perissoptera occurs more commonly in Europe than in America, although the genus is present in the Cenomanian of the Western Interior [Perissoptera prolabiata (White, 1876)] and the Campanian of the Gulf Coast (Perissoptera prolabiata mississippiensis Dockery, 1993). The only species described from the Pacific Slope is Perissoptera hannai (Allison, 1955) from the Aptian–Albian, Alisitos Formation of Baja California, Mexico."
The Mexican specimens differ from Perissoptera prolabiata (White, 1876) in that the species of the Colorado Formation have a large, broad wing; its outer border is nearly straight or slightly convex, its anterior extremity is abruptly rounded to a broad, concave front margin; the posterior wing is divided by a deep rounded sinus into two portions and larger in size.
Perissoptera prolabiata mississippiensis Dockery, 1993, from Coffee Sand, Mississippi is also a medium sized, axially ribbed, turriculated aporrhaid and has abroad bilobate outer lip with a long, narrow posteriorly directed upper lobe, but the Mexican specimens differ in that they are smaller in size and a broad bilobed outer lip is absent.
Perissoptera hannai (Allison, 1955) from the Alisitos Formation (Aptian–Albian), Baja California, is a moderate small shell with a high spire, with nine convex whorls, 14 heavy and smoothly rounded axial ribs and spiral ornamentation of approximately 15 narrow ribs; body whorl keel extended outward an slightly posterior from outer lip as long slender dilatation, anterior canal long. The Chiapas specimens are not this species and maybe this species could be Drepanocheilus.
Perissoptera marginata (Sowerby, 1829) (Gardner, 1857, p. 198, pi. 6, figs. 1–3) from Gault of Folkestone, Paris Basin, Mediterranean Basin, and Switzerland is composed of eight convex angulated whorls, which are closely covered with spiral striae, the whorls ornamented by short elongated ribs, which are nodose and tubercular on the lower, but thin and linear on the upper whorls. On the last whorl there are two more or less distinct rows of tubercular nodes; the uppermost row being the more important, is continued in the form of a tuberculated ridge on the wing process. The wing is large, broad and quadrate. The Mexican specimens have no nodose and tubercular ribs and have more whorls and are smaller shells.
Perissoptera parkinsoni (Mantell, 1822) (Gardner, 1875, p. 200, pi. 6, figs. 4–7) from the lower Albian, abundant in Folkstone, Cambridge and Blacktown, and the Albian of Belgium, Austria and Madagascar, is composed of nine or 10 convex whorls, which are finely striated spirally. Each whorl is rather irregularly ornamented by 16 to 20 or more, slightly flexuous, slender ribs, which have, though rarely, a tendency to produce varices. The last whorl wholly destitute of carinae, and is prolonged in a broad rounded expansion obliquely truncated at the extremity and sinuous at its anterior margin, where it units with canal; there is at the posterior margin a deep sinus equal to half the length of the wing, and above the sinus is a long recurved canaliculated point. The aperture is narrow, and the anterior canal moderately long. The Mexican specimens also differ from this species in number of ribs, in the wing and being smaller in size.
Class Insecta Linnaeus, 1758
Order Odonata Fabricáis, 1793
Suborder Zygoptera Selys–Longchamps, 1854
Figures 6.1, 6.2
Description. Odonate nymph of medium size. Head circular, eyes rounded; its length about one–eighth total length, width 2/3 maximum width (thorax). Abdomen about half total length, apparently with 6–7 segments. First pair of legs directed forward, second and third ones backward. Femur of first leg 34 % total leg length; femur of second leg 39 % (+ 5 %) the total length; femur of third leg 44 % (+ 5 %) the total length. Posterior wing pads 1/5 the total body length.
Material examined. One odonate nymph. Hypotype IHNFG–2839.
Occurrence. El Espinal quarry, Sierra Madre Formation, Chiapas, Albian.
Measurements. Body length: 21.0 mm, width: 4.6 mm; head length: 2.8 mm, width: 3.9 mm; pronotum length: 0.6 mm, width: 2.7 mm; thorax length: 5.9 mm, width: 5.7 mm; abdomen length: 11.4 mm, width: 4.6 mm. First right leg basis length: 2.6 mm, width 1.3 mm; femur length: 3.6 mm, width: 0.5 mm; tibia length: 2.9 mm, width: 0.3 mm; tarsus length: 1.5 mm, width: 0.2 mm. Second right leg basis length: 1.2 mm, width: 0.8 mm; femur length: 3.4 mm, width: 0.7 mm; tibia length: 2.9 mm, width: 0.5 mm; tarsus length: 2.5 mm, width: 0.2 mm. Third right leg basis length: 1.7 mm, width: 0.8 mm; femur length: 6.0 mm, width: 0.5 mm; tibia length: 3.5 mm, width: 0.6 mm; tarsus length: 2.6 mm, width: 0.2 mm.
Discussion. Feldmann et al. (1998) indicated that acute shape of legs, short separation between each pair of legs and well developed eyes are features that distinguish nymphs of the order Odonata. Furthermore, the general shape of the nymph (slender) resembles those of suborder Zygoptera (maximum width one–fifth total length) (de la Fuente, 1994). However, lack of diagnostic features (caudal gills), makes it difficult to identify our specimen with more detail.
Subclass Pterygota Brauer, 1885 Superorder Cimicidea Laicharting, 1781
Order Hemiptera Linnaeus, 1758
Suborder Heteroptera Latreille, 1810
Superfamily Hydrocorisae Latreille, 1810
Family ?Belostomatidae Leach, 1815
Figures 6.3, 6.5
Description. Body ovoid, head triangular, 1/10 the total length with prominent rounded eyes. Pronotum trapeziform, 1/4 the total length, posterior margin slightly wider than anterior margin. Scutellum semisquare, 1/7 the total length. Wings length 2/3 the total length; clavus 1/4 the total length; clavus comissure nearly 1/4 the total length. Right anterior leg directed forward; part of femur and tibia evident. Median legs directed backward; right median leg femur 1/3 the total length of leg, tibia and tarsus each about 1/4 the total length of leg.
Material examined. One adult specimen. Hypotype IHNFG–2840.
Occurrence. El Espinal quarry, Sierra Madre Formation, Chiapas, Albian.
Measurements. Total body length: 6.1 mm, maximum width: 3.0 mm; head length: 0.4 mm, width: 1.4 mm; pronotum length: 1.0 mm, width: 2.0 mm; abdomen length: 4.0 mm, width: 3.0 mm.
Discussion. Carpenter (1992, p. 259) indicated the hemelytron "Forewing typically with the proximal part strongly coriaceus (clavus and corium) and the distal part membranous" as a feature that distinguishes members of the suborder Heteroptera. The clavus (= anterior inner portion of the hemelytron) is clearly observed in our specimen; moreover, the rest of wings usually held flat over abdomen, and the clavus borders the scutellum, the triangular dorsal portion of the mid – thorax, located between the anterior portions of the hemelytra (Schuh and Slater, 1995; Epler, 2006) are also clearly observed. Stratigraphic range: Permian to Recent (Shcherbakov and Popov, 2002).
The specimen is referred to the Hydrocorisae by presence of short antennae, probably hidden below eyes; big compound eyes occupy both sides of head in dorsal and lateral views, and oval hydrodynamic shape typical of aquatic hemipterans (Schuhand Slater, 1995). Stratigraphic range of hemipterans: middle Triassic to Recent (Shcherbakov and Popov, 2002). A possible affinity with the Belostomatidae (Jurassic to Recent) is suggested by presence of eggs (8) on wings (Carpenter, 1992). Length of recent representatives of this family range from 9 to 112 mm.
Order Decapoda Latreille, 1802
Infraorder Palinura Latreille, 1802
Superfamily Palinuroidea Latreille, 1802
Family Palinuridae Latreille, 1802
Genus Palinurus Weber, 1795
Type species. Astacus elephas Fabricius, 1787, by monotypy. Recent, Eastern Atlantic.
Palinurus palaciosi new species
Figures 6.4, 6.6
Diagnosis. Carapace subrectangular, covered by numerous small, forward directed scale–like spines. Frontal horns triangular. Cervical groove weak, placed at carapace midlength. Abdominal somites smooth; pleural spines of somites I–V triangular, directed backward, pleural spine of somite VI suboval. Endopod and exopod of telson with external margin spinose. Pereiopods I–V long, with numerous setal pits on dactylus. Ischiocerite, carpocerite and merocerite of right antennular peduncle subcilindrical.
Description. Carapace subrectangular, covered by small, forward–directed scale–like spines, more numerous and pronounced on lateral margins. Small forward directed spines on anterior portion of carapace. Spines scarce and weaker on posterior portion of carapace. A strong, foreward–directed spine behind cervical groove. Cervical groove weak, perpendicular to lower margin. Frontal horns subtriangular, flat, strong, smooth, projected beyond anterior margin. Lower margin of left horn slightly convex; upper margin nearly straight; length of horns 1/5 that of carapace. Frontal margin between horns concave, with a small spine. Postmarginal groove weak, slightly narrower than cervical groove.
Abdominal segments subrectangular in lateral view. Abdomen length 6/5 that of carapace. Abdominal segment II the largest. Anterior groove of abdominal somites I–III stronger than in somites IV–VI. Abdominal somites smooth, no grooves evident. Pleural spines of somites I–V subtriangular, inclined about 45° posteriorly. Pleural spine of somite VI rounded, lobe–like. Small, rounded notch directed backwards at junction of each somite. Lateral junction between somites sinuous, curved below notch and becomes straight, perpendicular to lower margin, and curves forward again to reach upper margin of pleural spine. Right uropod protopod of telson subtriangular, outer margin with five teeth; right exopod also subtriangular, with five, stronger teeth on outer margin.
Right antennular peduncle with smooth, subcilindrical ischiocerite, carpocerite and merocerite. The first two segments of about the same height and length, merocerite sligthly longer and higher.
Pereiopods long, first pairs nearly twice as long as carapace. Basis and ichium indistinct. Meras rectangular, elongated, surface covered by small, spiny tubercles; carpus short, semirectangular, smooth, 1/5 the length of meras, covered by spiny tubercles; propodus long, rectangular, nearly as long as meras, also covered by spiny tubercles; dactylus semitriangular, 1/2 the length of propodus, covered by strong and numerous setal pits.
Material examined. A molt of medium–sized specimen. Holotype IHNFG–2841.
Occurrence. El Espinal quarry, Sierra Madre Formation, Chiapas. Albian.
Measurements. Holotype IHNFG–2481, carapace length: 67.2 mm, height: 20.5 mm; abdomen length: 74.7 mm, height: 25.8 mm; pereiopod meras length: 25.3 mm, height: 7.5 mm; carpus length: 11.0 mm, height: 6.0 mm; propodus length: 28.8 mm, height: 5.0 mm; dactylus length: 13.4 mm, height: 3.7 mm.
Etymology. The specific name honors Eliseo Palacios Aguilera, precursor of paleontology in Chiapas state.
Discussion. The sole specimen represents a molt; the carapace and abdomen still articulated but separated behind the postmarginal groove. Loss of many appendages and parts of telson suggest the exuvia suffered certain but not significative transport. From the species here described, Palinurus palaciosi new species represents the only estenohaline element.
Several differences with members of the genus Panulirus White, 1847 can be noted. Frontal horns are stronger in Palinurus, the frontal margin between horns concave and with a central teeth, and the surface is covered by smaller and less numerous spines.
Fossil representatives oí Palinurus are known from the Cretaceous (Turanian, Senonian) of Europe, from the Upper Cretaceous (Cenomanian) of Lebanon, and from the Tertiary (Middle Eocene) of Europe. Palinurus palaciosi new species differs from P. woodwardi Fristch, 1887 (Turanian of Germany), P. baumbergicus Schlüter, 1872 (Senonian of Germany) and from Palinurus sp. (Cenomanian of Lebanon, Garassino, 1994), in size and shape of frontal horns, which are larger and stronger in the Mexican specimen. Ornamentation of the carapace is also different, as the small scale–like spines of P. palaciosi are not observed in any of the previously reported taxa. An unconfirmed report ofPalinurus on Tertiary deposits of Europe reads as follows: " ... one specimen in Anchille de Zignoí s private collection was recorded by de Zigno himself as Palinurus desmaresti, making reference to Desmarest's earlier paper. However, de Zigno provided neither a description of the morphology of this taxon, nor an illustration." (Garassino and De Angeli, 2003). Two more reports deal with the occurrence of Palinurus in Lower Cretaceous (upper Barremian–Aptian; Albian) beds of NE Italy. Palinurus sp. from the Barremian–Aptian of Udine, Italy (Garassino, 2000), differs from P. palaciosi in having a carapace slightly shorter, ornamentation seem to consist of uniformly distributed tubercles, but frontal horns seem to be similar. The three specimens of Palinurus reported from the Albian of Petina, Italy (Bravi and Garassino, 1998) are smaller than the Mexican species. More detailed morphological comparison is difficult, due to the poor preservation of the specimens from south Italy. The Italian records represent the oldest occurrence of Palinurus, and P. palaciosi is the third oldest record and the first report for the genus as a fossil in America.
Infraorder Anomura MacLeay, 1838
Superfamily Paguroidea Latreille, 1802
Family Diogenidae Ortmann, 1892
Genus Roemerus Bishop, 1983
Type species. Roemerus robustus Bishop, 1983, by mono–typy. Albian, Texas.
Roemerus robustus Bishop, 1983
Figure 6.7
Roemerus robustus Bishop, 1983, p. 42, pi. 3, figs. 10–31, fig. 8g, table 4.
Roemerus robustus Bishop, 1986, p. 133, fig. 6J, table 1.
Description. Chelipeds equal. Meras subtriangular, much narrow to its base, maximum height distally; inner surface flat, smooth; outer surface concave, ornamented with tubercles. Carpus triangular, narrower to its lower margin, maximum length (1/2 that of carpus) on upper margin; highest at junction with manus; inner surface flat and smooth; outer surface concave, ornamented with tubercles. Manus subrectangular, nearly as high as long, slightly narrower in junction with carpus; three times as long as carpus; maximum height to its mid–length; semicircular in transverse section; inner surface flat, smooth; outer surface concave and ornamented with tubercles. Upper margin slightly convex; lower margin concave, with tubercles. Fixed finger triangular, maximum length (2/3 that of manus) at straight, lower margin; occlusal surface straight and smooth; color pattern preserved from mid–length to tip of finger. Movable finger triangular, deflexed, maximum length on convex upper margin; occlusal surface smooth, straight; junction to manus concave, 2/3 the maximum height; color pattern preserved from mid–length to tip of finger.
Fourth pereiopods long, 1/3 longer than first cheliped length. Meras rectangular, smooth, narrows to half its height to base, wider on junction with carpus. Carpus smooth, rectangular, as wide as meras and propodus, 1/5 total cheliped length. Propodus rectangular, 1/3 total cheliped length, with setal pits. Dactylus triangular, nearly unciform, with setal pits, 1/8 total cheliped length, with setal pits.
Partial propodus and dactylus of possible third pereio–pod preserved. Propodus rectangular, covered with setal pits. Dactylus semirectangular, covered with setal pits.
Material examined. Pair of chelae, fourth and partial third pereiopods. Hypotype IHNFG–2842.
Occurrence. El Espinal quarry, Sierra Madre Formation, Chiapas. Albian.
Measurements. Hypotype IHNFG–2482, right meras length: 6.9 mm, height: 3.9 mm; right carpus length: 1.1 mm, height: 3.9 mm; right manus length: 7.0 mm, height: 4.2 mm; left meras length: 7.1 mm, height: 2.1 mm; left carpus length: 2.3 mm, height: 2.6 mm; left manus length: 7.5 mm, height: 4.0 mm. Left fourth pereiopod meras length: 4.1 mm, height: 1.2 mm; left fourth pereiopod carpus length: 2.1 mm, height: 0.7 mm; left fourth pereiopod propodus length: 3.3 mm, height: 1.0 mm; left fourth pereiopod dactylus length: 3.1 mm, height: 0.7 mm.
Discussion. Schweitzer and Feldmann (2001) pointed out differences between the paguroid families, and stated that those specimens showing equal chelipeds belong to the Diogenidae. The Mexican specimen is identical with Roemerus robustus Bishop 1986, described from the lower Albian of the Glen Rose Limestone of Texas, on the basis of five specimens. Bishop (1983) noticed that this genus was similar in the cheliped shape to Palaoepagurus Van Straelen, 1925 and Petrochirus Stimpson, 1859, but these genera have a more elongate palm. Eocalinus Via, 1959 is also similar, but the lower margin of manus is more convex and more elongate chela than Roemerus (Beschin et al., 2002). Paguristes Dana, 1853, has an outer surface of manus covered with strong tubercles, and a more curved lower margin more curved (Feldmann et al., 1993; Collins et al, 1995). Schweitzer and Feldmann (2002) illustrated an indeterminate paguroid from the Eocene of California, that resembles the Mexican specimen, but the manus is ovate and upper margin convex.
Stratigraphic occurrence of R. robustus in lower Albian beds of Texas provides support to interpret an Albian age for the Sierra Madre Formation exposed at El Espinal quarry.
ACKNOWLEDGMENTS
Our sincere gratitude to H. Karasawa, R. H. B. Fraaije, and G. A. Bishop for their review and useful comments to the original manuscript. A. Garassino kindly provided important literature.
REFERENCES
Allison, E.C., 1955, Middle Cretaceous Gastropoda from Punta China, Baja California, Mexico: Journal of Paleontology, 29, 400–423. [ Links ]
Applegate, S.P., 1988, Anew genus and species of teleostean belonging to the Family Ophiopsidae, Teoichthys kallistos, from the Cretaceous near Tepexi de Rodríguez, Puebla: Universidad Nacional Autónoma de México, Instituto de Geología, Revista, 7, 200–205. [ Links ]
Applegate, S.P., 1992, A new genus and species of pycnodont from the Cretaceous (Albian) of central Mexico: Universidad Nacional Autónoma de México, Instituto de Geología, Revista, 10, 164–178. [ Links ]
Applegate, S.P., 1995, An overview of the Cretaceous fishes of the quarries near Tepexi de Rodríguez, Puebla, Mexico, in Arratia, G., Viohl, G. (eds.), Mesozoic Fishes –Systematics and Paleoecology: Munich, Germany, Friedrich Pfeil Verlag, 529–538. [ Links ]
Applegate, S.P., Espinosa–Arrubarrena, L., Alvarado–Ortega, J., Benammi, M., 2006, Revision of Recent investigations in the Tlayua quarry, in Vega, F.J., Nyborg, T.G., Perrilliat, M.C., Montellano–Ballesteros, M., Cevallos–Ferriz, S.R.S., Quiróz–Barroso, S.A. (eds.), Studies on Mexican Paleontology: The Netherlands, Springer, Topics in Geobiology, 24, 265–291. [ Links ]
Beschin, C, Busulini, A., De Angeli, A., Tessier, G., 2002, Aggiornamento ai crostacei eocenici di cava "Main" di Arzignano (Vicenza – Italia Settentrionale) (Crustacea, Decapoda): Montecchio Maggiore, Vicenza, Museo Civico "G. Zannato", Associazione Amici del Museo, Studi e Ricerche, 2002, 7–28. [ Links ]
Bishop, G.A., 1983, Fossil decapod crustaceans from the Early Cretaceous Glen Rose Limestone of Central Texas: Transactions of the San Diego Society of Natural History, 20, 27–55. [ Links ]
Böse, E., 1905, Reseña acerca de la geología de Chiapas y Tabasco: México, Boletín del Instituto Geológico, 20, 5–100. [ Links ]
Brauer, F., 1885, Systematische–zoologische Studien: Sitzungsberichte der Keiserlichen Akademie der Wissenschaften in Wein, mathema–tische–naturwissenschaftlichen Klasse, 91, 237–431. [ Links ]
Bravi, S., Garassino, A., 1998, Plattenkalk of the Lower Cretaceous (Albian) of Petina, in the Alburni Mounts (Campania, S Italy) and its decapod crustacean assemblage: Atti della Societá italiana di Scienze naturali e del Museo civico di Storia naturale in Milano, 138,89–118. [ Links ]
Carpenter, F.M., 1992, Treatise on invertebrate paleontology. Part R Arthropoda 4, v. 3 Superclass Hexapoda: Lawrence, The University of Kansas and The Geological Society of America, 271 p. [ Links ]
Castro–Mora, J., Shleper, C, Martínez–Rodríguez, E., 1975, Estratigrafía y microfacies del Mesozoico de la Sierra Madre del Sur, Chiapas: Boletín de la Asociación Mexicana de Geólogos Petroleros, 27(1–3), 103 p. [ Links ]
Chubb, L.J., 1959, Upper Cretaceous of Central Chiapas, Mexico: American Association of Petroleum Geologists, Bulletin, 43, 725–756. [ Links ]
Collins, J.S.H., Fraaiye, R.H.B., Jagt, J.W.M., 1995, Late Cretaceous anomurans and brachyurans from the Maastrichtian type area: Acta Palaeontologica Polonica, 40(2), 165–210. [ Links ]
Cossmann, M., 1904, Essais de Paléoconchologie Comparée, v. 6: Paris, privately published, 151 p. [ Links ]
Costa, E.M. da, 1778, Historia Naturalis Testaceorum Britanniae, or the British Conchology; containing the descriptions and other particulars of natural history of the shells of Great Britain and Ireland: London, privately published, 254 p. [ Links ]
Cox, L.R., 1959, Thoughts on the classification of the Gastropoda: Proceedings of the Malacological Society of London, 33, 239–261. [ Links ]
Cuvier, C, 1797, Tableau élementaire de l'histoire naturelle des animaux: Paris, Baudouin, 710 p. [ Links ]
Dana, J.D., 1853, Crustacea, in United States Exploring Expedition during the Years 1838, 1839, 1840, 1841, 1842 under the command of Charles Wilkes, U.S.N. v. 13–14: Philadelphia, C. Sherman, 1618 p. [ Links ]
De la Fuente, J.A., 1994, Zoología de Artrópodos: España, McGraw–Hill, 805 p. [ Links ]
Dockery, D.T. III., 1993, The Streptoneuran gastropods, exclusive of the Stenoglossa, of the Coffee Sand (Campanian) of northeastern Mississippi: Jackson, MS, Mississippi Department of Environmental Quality, Office of Geology, Bulletin, 129, 191 p. [ Links ]
Epler, J.H., 2006, Aquatic and semiaquatic Heteroptera of Florida: Tallahassee, FL, Florida Department of Environmental Protection, Division of Water Resource Management, 195 p. [ Links ]
Fabricius, J.C., 1787, Mantissa insectorum sistens eorum species nuper detectas adjectis characteribus generis, differentiis specifis, emendationibus, observationibus: Hafniae, C.G. Proft, 348 p. [ Links ]
Fabricius, J.C., 1793, Entomológica Systematica, v. 2: Hafniae, C.G. Proft, 519 p. [ Links ]
Feldmann, R.M., Tshudy, D.M., Thomson, M.A., 1993, Late Cretaceous and Paleocene Decapod Crustaceans from James Ross Basin, Antarctic Peninsula: Lawrence, KS, The Paleontological Society, Memoir, 28, Supplement to Journal of Paleontology, 67(1), 41 p. [ Links ]
Feldmann, R.M., Vega, F.J., Applegate, S.P., Bishop, G.A., 1998, Early Cretaceous arthropods from the Tlayúa Formation at Tepexi de Rodríguez, Puebla, México: Journal of Paleontology, 72, 79–90. [ Links ]
Fristch, A., 1887, Malacostraca – Ordnung Decapoda, in Fritsch, A., Kafka, J. (eds.), Die Crustaceen der bomischen Kreideformation: Prague, Selbstverlag, 20–53. [ Links ]
Garassino, A., 1994, The macruran decapod crustaceans of the Upper Cretaceous of Lebanon: Paleontología Lombarda, Nouva serie, 3, 1–27. [ Links ]
Garassino, A., 2000, Palinurus sp. (Crustacea, Decapoda) del Cretácico inferiore (Barremiano superiore–Aptiano) della Valle del Torrente Cornappo (Udine, NE Italia), Gortania: Udiine, Atti del Museo Friulano di Storia Naturale, 22, 65–68. [ Links ]
Garassino, A., De Angeli, A., 2003, Justitia Holthuis, 1946, (Crustacea, Decapoda, Palinuridea) from the Middle Eocene of Verona and Vicenza (northern Italy): Contributions to Zoology, 72(2/3), 131–132. [ Links ]
Gardner, J.S., 1875, On the Gault Aporrhaidae: The Geological Magazine, New Series, Decade II, 2, 198–203. [ Links ]
González, K., Garcia, P., Avendaño, J., 2002, Un nuevo registro de la familia Macrosemiidae (Neopterygii–Halecostomi) en Chiapas, sureste de México (abstract), in XVI Convención Geológica Nacional y III Reunión Nacional de Ciencias de la Tierra, Puerto Vallarta, Jal., Libro de Resúmenes: México, D.F., Sociedad Geológica Mexicana, p. 303. [ Links ]
Gutiérrez–Gil, R., 1956, Geología del Mesozoico y Estratigrafía Pérmica del Estado de Chiapas, in 20th International Geological Congress, Guidebook, Excursion C–15: México, D.F., International Geological Congress, 15–23. [ Links ]
Imlay, R.W., 1944, Cretaceous formations of Central America and Mexico: American Association of Petroleum Geologists, Bulletin, 28, 1077–1195. [ Links ]
Laicharting, J.N. von., 1781, Verzeichniss und Beschriebung der Tyroler Insecten 1: Zurich, Fuessley, 248 p. [ Links ]
Latreille, PA., 1802, Histoire naturelle genérale et particuliére des Crustacés et des Insectes, v. 3: Paris, Dufart, 467 p. [ Links ]
Latreille, PA., 1810, Considerations genérales sur l'ordre naturel des animaux: Paris, F. Schoell, 123 p. [ Links ]
Leach, W.E., 1815, A tabular view of the external characters of four classes of animals, which Linné arranged under Inserta, with the distribution of the genera composing three of these classes into orders, and description of several new genera and species: Transactions of the Linnean Society of London, 11, 306–400. [ Links ]
Linnaeus, C. von., 1758, Systema Naturae per Regna Tria Naturae: secundum classes, ordines, genera, species, cum characteribus diferentiis synonymis, locis: Holmiae, Laurentius Salvius, 10th ed., revised, (Facsimle, British Museum (Natural History) 1956), 824 p. [ Links ]
MacLeay, C.L., 1838, On the brachyurous Crustacea brought from the Cape by Dr Smith, in Illustrations of the Zoology of South Africa, 5. Invertebrate: London, Smith Elder and Co., 53–71, [ Links ]
Mantell, G.A., 1822, Fossils of the South Downs, or illustrations of the geology of Sussex: London, Lupton Relfe, 372 p. [ Links ]
Michaud, F., 1987, Stratigraphie et Paleogeographie du Mesozoique du Chiapas (sud est du Mexique: Paris, Université Marie Curie, Memoires Sciences de la Terre, (Ph.D Thesis), 300 p. [ Links ]
Müllerried, F.K.G., 1936, Estratigrafía preterciaria preliminar del Estado de Chiapas: Boletín de la Sociedad Geológica Mexicana, 9, 31–41. [ Links ]
Ortmann, A., 1892, Die Dekapoden–Krebse des Strassburger Museums, IV Die Abtheilungen Galatheidea und Paguridea: Jena, Zoologische Jahrbücher, Abtheilung für Systematik, Geographie und Biologie derTiere, 6, 241–326. [ Links ]
Ovalles–Damián, E., 2004, Determinación taxonómica de un paraclupéido (Teleostei: Clupeomorpha) fósil de la Cantera El Espinal, Ocozocoautla, Chiapas: Tuxtla Gutiérrez, Mexico, Universidad de Ciencias y Artes de Chiapas, Tesis profesional, 73 p. [ Links ]
Quezada–Muñetón, F.M., 1987, El Cretácico medio–Superior y el límite Cretácio Superior–Terciario inferior en la Sierra de Chiapas: Boletín de la Sociedad Mexicana de Geólogos Petroleros, 39, 3–98. [ Links ]
Rafinesque, C.S., 1815, Analyse de la nature, ou Tableau de l'univers et des corps organises: Palermo, 224 p. [ Links ]
Sánchez–Montes de Oca, R., 1969, Estratigrafía y paleontogeografía del Mesozoico de Chiapas, in Seminario sobre Exploración Petrolera, Mesa Redonda 5, Problemas de exploración de la zona sur: Mexico, Instituto Mexicano del Petróleo, 31 p. [ Links ]
Saul, L.R., 1998, Eight Aporrhaid Gastropod species from the Cretaceous of the Pacific Slope of North America and Clarification of the Type Species of Perissoptera: The Nautilus, 111, 119–142. [ Links ]
Schuh, R.T., Slater, J.A., 1995, True bugs of the World (Hemiptera: Heteroptera). Classification and natural history: Ithaca, Cornell University Press, 350 p. [ Links ]
Schlüter, H.C. von, 1872, Die Macruren Decapoden der Senon– und Cenomanian–Bildungen Westphalens: Zeitschrift Deutsche Geologische Gesellschaft, 14, 702–749. [ Links ]
Schweitzer, CE., Feldmann, R.M., 2001, New Cretaceous and Tertiary decapod crustaceans from western North America: Bulletin of the Mizunami Fossil Museum, 28, 173–210. [ Links ]
Schweitzer, C.E., Feldmann R.M., 2002, New Eocene decapods (Thalassinidea and Brachyura) from Southern California: Journal of Crustacean Biology, 22(4), 938–967. [ Links ]
Selys–Longchamps, M.E. de., 1854, Monographie des Caloptérygines: Bruxelles, Mémoires de la Société Royale des Sciences de Liége, 9, 291 p. [ Links ]
Shcherbakov, D.E., Popov, Y.A., 2002, Superorder Cimicidea Laicharting, 1781 Order Hemiptera Linne, 1758. The bugs, cicads, scale insects, etc, in Rasnitsyn, A.P, Quicke D.L.J. (eds), History of Insects: Dordrecht, Kluwer Academic Publishers, 143–157. [ Links ]
Steele, D.R., Waite, L.E., 1986, Contributions to the stratigraphy of the Sierra Madre Limestone (Cretaceous) of Chiapas: Universidad Nacional Autónoma de México, Instituto de Geología, Boletín 102, 1–175. [ Links ]
Stimpson, W., 1859, Notes on North American Crustacea: New York, Natural History Annuary. 7–53. [ Links ]
Straelen, V van, 1925, Contribution a l'étude des crustacés décapodes de la période jurassique: Academie Royale de Belgique, Cl. Scientifiqué, Mémoire, 4, series 2, 7, 1–462. [ Links ]
Sowerby, J., 1829, The Mineral Conchology of Great Britain or coloured figures and descriptions of those remains of testaceous animals or shells, which have been preserved at various times and depths in the Earth, v. 6: London, 250 p. [ Links ]
Tate, R., 1865, On the so called Rostellariae of the Cretaceous rocks, with a descriptive catalogue of the British species: Geological and Natural History Repertory 1 (for 1865): 93–102. [ Links ]
Ver Wiebe, W.A., 1925, Geology of southern Mexico oil fields: Pan American Geology, 94, 121–138. [ Links ]
Via, L., 1959, Decápodos fósiles del Eoceno español: Instituto Geológico y Minero de España, Boletín, 70, 333–395. [ Links ]
Weber, F, 1795, Nomenclátor entomologicus secundum Entomologiam Systematicum ill. Fabricii adjectis speciebus recens detectis et varietatibus: Chilonii and Hamburgi, Germany, Carolum Ernestum Bohn, 171 p. [ Links ]
White, A., 1847, List of species of Crustacea in the collection of the British Museum: London, 1–143. [ Links ]
White, C.A., 1876, Invertebrate paleontology of the Plateau Province, in Powell, J. W. (ed.), Report on the Geology of the Eastern Portion of the Uinta Mountains and a region of country adjacent thereto. With Atlas, Chapter 3: Washington, United States Geological and Geographical Survey of the Territories, Government Printing Office, 74–135. [ Links ]
Zavala–Moreno, J.M., 1971, Estudio geológico del proyecto hidroeléctrico Cañón del Sumidero, Río Grijalva, Estado de Chiapas: Boletín de la Asociación Mexicana de Geólogos Petroleros, 23, 1–92. [ Links ]

