










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkRevista mexicana de ciencias geológicas
On-line version ISSN 2007-2902Print version ISSN 1026-8774
Rev. mex. cienc. geol vol.23 n.3 Ciudad de México Jan. 2006
Revista mexicana de ciencias geológicas
Taxonomy, stratigraphic distribution and palaeobiogeography of the Early Cretaceous coral genus Holocystis
Hannes Löser
1 Estación Regional del Noroeste, Instituto de Geología, Universidad Nacional Autónoma de México, Apartado Postal 1039, 83000 Hermosillo, Sonora, Mexico.
E–mail: loeser@paleotax.de
Manuscript received: April 25, 2006
Corrected manuscript received: August 8, 2006
Manuscript accepted: August 28, 2006
ABSTRACT
The Early Cretaceous Scleractinian coral genus Holocystis Lonsdale, 1849 –first reported for southern England and for a long time believed to be restricted to this region– is easy to recognise but it is also rare. Abundant material from the Late Barremian to Early Albian found in Sonora (Mexico) as –well as the available type material and additional material from Europe and East Africa allowed a systematic revision. Five species are distinguished. One of them –Holocystis nomikosi– is described as a new species. Three species previously described for other genera were assigned to the genus Holocystis and two species formerly assigned to Holocystis were found to belong to other genera. Two genera (Tetracoenia and Nowakocoenia) are considered junior synonyms of Holocystis. The stratigraphic extent of the genus was limited to the range from the Barremian to the Early Albian. Holocystis is nota common coral genus but occurred in a large geographic area comprising the central Tethys, the Caribbean, and even the eastern to south–eastern Tethys. The genus is restricted to sediments with a certain terrigenous input.
Key words: Scleractinia, Cretaceous, taxonomy, corals, palaeobiogeography.
RESUMEN
El género de corales Holocystis Lonsdale. 1849 del Cretácico Temprano fue reportado por primera vez en el sur de Inglaterra y por mucho tiempo se consideró como restringido a esta región. El género es fácil de reconocer pero es un género raro. Abundantes muestras del Barremiano tardío hasta el Albiano temprano encontradas en Sonora (México), así como el material tipoy muestras adicionales de Europa y Africa oriental permitieron una revisión sistemática. Se identifican cinco especies. Una especie –Holocystis nomikosi– se describe como especie nueva. Tres especies originalmente asignadas a otros géneros se reasignan a Holocystis. y dos especies originalmente asignadas a este género pertenecen a otros géneros. Dos géneros (Tetracoenia y Nowakocoenia) son considerados como sinónimos de Holocystis. El rango estratigráfico del género es limitado del Barremiano hasta el Albiano temprano. Holocystis no es un género común de coral, pero se encuentra en un área grande que comprende la región central del mar de Tethys, el mar Caribe y también en le este y sureste del mar de Tethys. El género está casi restringido a sedimentos con una cierta influencia terrigena.
Palabras clave: Scleractinia, Cretácico, taxonomía, corales, paleobiogeografia.
INTRODUCTION
The colonial coral genus Holocystis Lonsdale, 1849 is an easily recognisable Early Cretaceous genus owing to its tetrameral septal arrangement. It was first described from the Early Aptian of Southern England, with Cyathophora elegans Lonsdale, 1849 as type species, and is a common coral in the Lower Greensand at the south coast of the Isle of Wight. Five Holocystis species were described subsequently from Lower Cretaceous outcrops in Europe, two of which belong to other genera. Three other species were described under genera different from Holocystis but belong to this genus. Only two species are reported commonly in the literature. Except for the study by Ryder (1927), which is restricted to the type species of the genus, the genus has never been considered in detail. New, previously unreported material mainly from the Late Barremian to Early Albian from Mexico, but also from Spain and Greece and the discovery of a new species demand a taxonomical revision of the genus, and also will allow a better limitation of its stratigraphical extent.
MATERIAL
The material comes from various localities. Most of them are listed, commented and provided with additional references in Löser et al. (2005); the locality code is provided in parenthesis after the locality name. Herein, details are only provided if not already reported in that publication.
Azerbaijan, Kubatlinskij rajon, Alikuliushagi (AZ.146); Barremian.
France, Dépt. Aube, Les Croûtes (F.417). According to d'Orbigny (1851, p. 165), the locality has an Aptian age, which was later specified by Corroy (1925, p. 456) as Early Aptian. An Aptian age is improbable.
France, Dépt. Yonne, Seignelay. Without stratigraphy mentioned by d'Orbigny (1850, p. 121). Corroy (1925, p. 456) claimed an Early Aptian age, which is improbable according to the geological situation. In this area, which concerns also Les Croûtes, sediments of the Calcaire à Spatangus (Early Hauterivian, zone of Acanthodiscus radiatus) crop out (F.418).
Germany, Bavaria, Allgäuer Helvetikum, Tiefenbach (D.256); Schrattenkalk, Early Aptian.
Greece, Viotía, Chiarmena SE Aliartos (GR.537); Evangelistria–Schichten, Late Aptian–Early Albian.
Italy, L'Aquila, Abruzzi, Monti d'Ocre, Fossa Cerasetti (1.1734); Late Aptian.
Mexico, Sonora, Municipio Ures, Cerro de Oro; Cerro de Oro Formation (Late Barremian–Early Aptian) and Cerro La Espina Member, Mural Limestone (Early Albian). Corals are found in two different levels in the area of the small village Cerro de Oro: in the Late Barremian–Early Aptian (zone of Palorbitolina lenticularis) of the Cerro de Oro Formation and in the Early Albian of the Cerro La Espina Member, Mural Limestone (both Bisbee Group; González–León and Lucas, 1995). Two sample locations are distinguished in the Cerro de Oro Formation (CO4 and BZl) and one sample location in the Cerro La Espina Member (CO5). Lawton et al. (2004) claimed a Middle Albian age in comparable sections for the Cerro La Espina Member, which is not well constrained by biostratigraphic data according to the opinion of this author, and therefore was not applied in this paper.
Poland, Malopolskie, Lanckorona, Jastrzebia (PL. 1170); Grodziszcze Sandstones, Early Aptian.
Poland, Slaskie, Bielsko–Biala, Rudzica (PL.3627); Late Barremian–Early Aptian.
Romania, Suceava, Pojorîta area, Cîmpulung–Moldovenesc, Valea Seaca and Valea Izvorul Alb (RO.280, RO.827); Wildflysch, Early Aptian.
Serbia, East Serbia, Zljebine (SM.126); Early Aptian.
Spain, Cataluña, Lérida, Com. Alt Urgell, Mun. Cabo, Senyús section (E.2273); Senyús Formation, early Late Aptian.
Spain, Cataluña, Lérida, Com. Alt Urgell, Mun. Coll de Nargó, Set Comelles, El Caso section (E.2040); Font Bordonera Formation, latest Aptian.
Spain, Cataluña, Lérida, Com. La Noguera, Montsec de Rubies, sectionNWLa Cabrua quarry; early Late Aptian. The coral fauna from this locality is still undescribed, but the stratigraphy of the locality was discussed in Löser and Decrouez (2000).
Spain, Murcia, Jumilla, Sierra deSopalmo; Early Aptian The locality is described in Morycowa et al. (2001).
Spain, Tarragona, Cataluña, Com. BaixPenedés, Mun. Sant Martí Sarroca, Can Grau (E.834); Late Aptian.
Tanzania, Tanganyika, Mtwara, Nambawala plateau, Pilepile (EAT. 16); Kiturika member, Late Aptian.
Turkmenistan, Krasnovodskaya oblast, Small Balean (TUR. 137); Early Barremian.
United Kingdom, Isle of Wight, Atherfield (GB.312); Perna Beds, Early Aptian. The locality at the coast has been well known for many years. The localities Atherfield point, coast guard station, Sandown (bay) and Shanklin are close to the type area and of Early Aptian age as well.
SYSTEMATIC PALAEONTOLOGY
The following institutional abbreviations are used: BSP: Bayerische Staatssammlung für Paläontologie und Geologie München, Germany; ERNO: Universidad Nacional Autónoma de México, Instituto de Geología, Estación Regional del Noroeste, Hermosillo, Mexico; GPS: Geologische und Paläontologische Sammlung der Universität Leipzig, Germany; JCSV: Coll. Saldaña–Villodre, San Vicente del Raspeig, Spain; MB: Naturkundemuseum der Humboldt–Universitat Berlin, Germany; MGSB: Museo Geológico del Seminario de Barcelona, Spain; MNHN: Muséum National d'Histoire Naturelle Paris, France; NHM: The Natural History Museum London, United Kingdom; NHMW: Naturhistorisches Museum Wien, Austria; PU: Museo di Geologia e Paleontología dell' Università di Torino, Italy; TUM: The Tohoku University Museum, Sendai, Japan; UJ: Jagiellonian University, Instytut Nauk Geologicznych, Kraków, Poland.
The following abbreviations are used: c: calicular diameter; ccd: distance of calicular centers; s: number of septa; sc: number of costae; ac: acetate peel; ths: thin section; v = the present author has seen the material belonging to this citation; * = first publication. The term 'cycle' or 'septal cycle' refers to radial elements of equal width and length. The first three cycles are assumed to have formed at the same time (Milne–Edwards 1857, p. 45).
Order Scleractinia Bourne, 1900
Suborder Stylinina Alloiteau, 1952
Family Cyathophoridae Vaughan and Wells, 1943
Genus Holocystis Lonsdale, 1849
Type species. Cyathophora elegans Lonsdale, 1849, by monotypy, from the Lower Greensand of Atherfield (Isle of Wight, United Kingdom).
Diagnosis. Massive colonial coral with sub–cerioid or plocoid calicular arrangement. Calices round, well distinguished, separated by a mostly narrow coenosteum. Septa short, regular in four systems and up to three septal cycles. A fourth septal cycle may appear, but is invariably incomplete. Septa are thin, compact, invariably free, and broad in their peripherical part. They do not bear auriculae (small winged ornamentations at the inner septal margin; Gill, 1977). A columella is absent. The endotheca consists of regular tabulae and/or dissepiments. The coenosteum consists of dissepiments.
Synonym. Tetracoenia (type species Tetracoenia dupiniana d'Orbigny, 1850, by monotypy). The types of the type species could not be found recently in the collections of the MNHN. D'Orbigny indicated the species from two localities: Early Aptian of Les Croûtes (Dépt. Aube, France) and from Seignelay (Dépt. Yonne, France; without stratigraphy). Both villages are in areas where only Early Hauterivian sediments typically yield colonial corals. However, Holocystis could not be found in Museum collections or in the widely exposed sediments of the Early Hauterivian of the Paris Basin. According to the description and illustration given by d'Orbigny (1851), the genus is most likely synonymous with Holocystis. Wery (1954, p. 41) distinguished Holocystis and Tetracoenia by the extent of their coenosteum. This character may vary within one colony and cannot be used to distinguish these genera. Wery (1954) did not note whether or not he had the type material of Tetracoenia dupiniana in hand.
Possible synonym. Nowakocoenia Kolodziej and Gedl, 2000 (type species Nowakocoenia cieszynica Kolodziej and Gedl, 2000, by original designation). The authors of the genus distinguished it from Holocystis on the basis of morphological characteristics of wall formation. The character of the wall was described as "ambiguous, however at least in some places of the colony", and they claimed that it had developed prior to the septa. They did not compare their distinctions with Holocystis. It was also claimed that the septal pattern oí Holocystis was more regular and "different" from that in Nowakocoenia, without going into detail. The septal insertion in Nowakocoenia is indeed not very regular, but neither is it in Holocystis. The main distinguishing feature proposed by the authors of Nowakocoenia –the development of the wall before the septa– is based on thin sections showing strongly reduced septa and the presence of the wall (Kolodziej and Gedl, 2000, figs. 9C, D). The extreme reduction of the septa to small ridges is no exception in genera of the Cyathophoridae and Stylinidae. But it cannot be claimed in such cases that the wall developed first and the septa later. Serial sections beginning from the top of the colony would be needed to support these arguments, but no such sections were provided by the authors of Nowakocoenia. Until such is proven to be the case in the type material of Nowakocoenia cieszynica, it is considered synonymous with Holocystis dupini, and therefore the genus is synonymous with Holocystis.
Not regarded as synonyms anymore. Cyathocoenia de Fromentel, 1884 non Duncan, 1867 nee Volz, 1896 (type species Cryptocoenia neocomiensis d'Orbigny, 1850, subsequently designated by Wells, 1936) was originally created by de Fromentel (1884, p. 541) as a subgenus oí Cyathophora. It was included in the synonymy of Holocystis by Wells (1956, p. 375). The lectotype oí Cryptocoenia neocomiensis (designated by Wells, 1936; MNHN, Coll. d'Orbigny, no. 5274) belongs to Cryptocoenia d'Orbigny, 1849. Therefore Cyathocoenia de Fromentel, 1884 cannot be a junior synonym of Holocystis, but probably of Cryptocoenia or Adelocoenia (which are probably synonymous genera).
Occurrence. The genus occurs in the Boreal and Tethys realms (from Europe to middle Asia and east Africa) and in the Caribbean province (Mexico). It ranges from the Barremian (Azerbaijan and Turkmenistan) to the Early Albian (Greece and Mexico). Holocystis predominantly occurs in siliciclastic deposits or carbonate deposits with an influence of siliciclastic sediments (as lagoonal environments or carbonate ramps). In pure platform carbonates the genus is rare.
Description. Massive sub–cerioid to plocoid corallum with more or less densely arranged calices. Each calice has its own wall. Calicular outline round or slightly quadratic. Septa free, confluent to subconfluent, compact, straight. The septa of the first cycle reach up to 40 percent of the calicular radius but may be longer. They are commonly wedged–shaped. Those of the second cycle are shorter and reach up to 20 percent of the radius. Those of the third cycle are very short, commonly only represented by ridges. The septa do not have auriculae at their inner margins. The septal flanks and the septal upper border are smooth.
The first and second septal cycles each encompass four septa, the third cycle eight septa, and the fourth up to 16 septa. The last septal cycle may be incomplete. The fourth cycle is in all examined samples incomplete and the maximum number of septa found is 24 (fourth cycle with eight septa). The septa of the different cycles can be easily distinguished by their length and thickness.
The coenosteum consists of the septal costae and dissepiments. It can be strongly reduced. The endotheca is made of tabulae and dissepiments. In some species (such as H. elegans), tabulae predominate, in others (such as H. nomikosi) dissepiments dominate. The wall is parathecal. Columella, pali and synapticulae are absent. The budding of new calices is extracalicular.
Remarks. Lonsdale (1849, p. 83) established the new genus only provisionally and cited the type species still under the genus Cyathophora. Milne–Edwards and Haime (1851, p. lxiv) confirmed the genus and Lonsdale as author. Ryder (1927) considered the ontogenetic stages and the nature of the septal insertation. Observations on the type and additional topotypic material show that the septal insertion is different from the description of Ryder (1927): new calices successively develop the four septa of the first cycle. Four or six septa do not appear at the same time. After the first cycle is completed, the calice gains size, and then the second cycle appears. The third cycle normally appears before the second cycle is completed. Therefore, calices can be found that have reached the average ' adult' calicular diameter with six, eight, ten, 12 or 14 septa.
Ryder (1927) also mentioned the rare existence of a columella. None of the specimens of Holocystis elegans from the type locality, which were observed directly, show any trace of a central calicular structure.
Relationship. Holocystis can be considered as a successor of the Late Jurassic and Early Cretaceous Pseudocoenia d'Orbigny, 1850 (sensu d'Orbigny, 1850; Löser, 1998b) which shows an octameral symmetry. Holocystis compares also with the genera A delocoenia and Cryptocoenia, which show hexameral symmetry, and with the pentameral Pentacoenia d'Orbigny, 1850.
Scleractinian corals with tetrameral septal symmetry, as in Holocystis, are rare. Besides Holocystis, there is one species of Heterocoenia (Heterocoenia tetrasepta Eguchi, 1951, Aptian of Japan), one species of Stylina (Stylina sucrensis Wells, 1944, Aptian of Venezuela), two –probably synonymous– species of Paronastraea (Paronastraea bassanii (Prever, 1909), and Paronastraea serrata (Prever, 1909), both Late Aptian of Italy), another species of Paronastraea (P. occulta Löser, 2006), and finally one still undescribed species of Enallhelia (labelled as Enallhelia rathieri d'Orbigny, 1850; unpublished specimen TUM without number, depository number D21L–L25, Albian of Japan) with a tetrameral symmetry.
Species. So far, six coral species have been assigned to the genus Holocystis (bukowinensis, calzadai, elegans, octolobata, polyspathes, similis; Löser et al., 2002; Kuzmicheva, 2002). The species are distinguished on the basis of their calicular diameter and the number of septal cycles. Calicular diameters vary between 1.3 and 4.1 mm. Within one colony, there is a maximum variation of 38 per cent between the smallest and the largest calicular diameter; the average of percentual variation is 20 per cent. The distance of the calices shows a higher variation of up to 53 per cent, the average is 22 per cent. The number of septa varies between 4 and 24. The variation of the number of septa within one colony is low: only half of the material included in the calculation shows any variation. In most cases, the number of septa varies in an interval between eight and 16, rarely between four and eight.
In the following synonymy lists, pure quotations of taxa in the literature are mainly suppressed. First, it cannot be proven whether the concerned quotation belongs to the species or not, and second, the data have been already published elsewhere (Löser et al., 2002). Localities are given only for material that was observed directly, or the description/depiction in the literature allows a sure comparison, and the locality data are precise and reliable.
Holocystis ?bukowinensis Volz, 1903
Figures 1a–c
* 1903 Holocystis bukowinensis; Volz, p. 27, pi. 4, figs 14–17.
v 1909 Polytremacis magnicanaliculata; Prever, p. 68, pi. 1, fig. 20, fig. 22 a.
v non 1964 Holocystis bukowinensis Volz; Morycowa, p. 25, pi. 3, fig. 3, pi. 5, fig. 1 [? =Holocystis elegans].
? 1971 Holocystis bukowinensis Volz; Morycowa, p. 44, pi. 6, fig. 4.
. 1981 Holocystis bukowinensis Volz; Turnsek and Mihajlovic, p. 19, pi. 13, figs 5–6.
non 1987 Holocystis bucowinensis Volz; Kuzmicheva, p. 222, pi. 1, fig. 3 [=Holocystis dupini].
non 1988 Holocystis bukowinensis Volz; Kuzmicheva and Aliev, p. 157, pi. 1, fig. 4 [=Holocystis dupini].
v non 1989 Holocystis bukowinensis Volz; Morycowa, p. 61, pi. 19, fig. 2 [? =Holocystis elegans].
non 2002 Holocystis bukowinensis Volz; Kuzmicheva, p. 165, pi. 23, fig. 3 [? =Pentacoenia sp.].
Type. The collection of Wilhelm Volz, held at the former Natural History Museum in Breslau (now: Muzeum Geologiczne im. Henryka Teisseyre, Instytut Nauk Geologicznych Uniwersytetu Wroclawskiego, Wroclaw) was almost completely destroyed during the World War II. The type(s) oí Holocystis bukowinensis must be considered lost, according to A. Pacholska (personal communication, 2002). A neotype has not yet been selected. Topotypical material could not be made available.
Dimensions. Volz (1903, p. 27) c: 1.3–1.5 mm, ccd: 3.0 mm, s: 8, sc: 16; Morycowa (1971, p. 44) c: 1.3–1.7 mm, ccd: 1.5–2.5 mm, s: 16, sc: 16; present material c: 1.2–1.7 mm, ccd: 1.4–2.4 mm, s: (4) 8–10, sc 16.
Description. Small platy colonies with a cerioid appearance, but sub–cerioid structure. The calices are small and have irregular outlines or are quadratic. There are two cycles of septa. The first cycle remains short and reaches only 15 percent of the calicular diameter. Septa of the second cycle are much shorter and reduced to short ridges. The second cycle is in some cases not complete. Endotheca and coenosteum of the material are known only from sample PU 18118. The coenosteum is formed by sub–confluent to confluent costae, up to 16 per calice. The endotheca is made of strong tabulae and thin dissepiments.
Remarks. The characteristics of the species are presently not clearly known due to the circumstance that the type is lost, and material from the type area contradicts with the original description. Unfortunately, the material described by Morycowa (1971, p. 44) cannot be currently found in the UJ collection. Morycowa confirmed (personal communication, 2005) only that the single specimen described by her was small and poorly preserved. The descriptions given by Volz (1903) and Morycowa (1971) differ in the number of septal cycles. If the number of septa reaches 16, H. bukowinensis is close to, perhaps even synonymous with H. dupini, and to the material with eight septa the species name magnicanaliculata must be applied. If the number of septa is eight, the material previously described by Morycowa (1971) might be assigned to H. dupini.
Occurrence. Roadcut near Perachorion, NW Levadia (Viotia, Greece), Early Aptian, BSP 2003 XX 5754; section NW La Cabrua quarry (Montsec de Rubies, Com. La Noguera, Lérida Cataluña, Spain), early Late Aptian, BSP 2003 XX 4200; Fossa Cerasetti, Monti d'Ocre (Abruzzi, L'Aquila, Italy), Late Aptian, PU18118 (holotype of Polytremacis magnicanaliculata).
Occurrences according to the literature. Valea Seaca and Valea Izvorul Alb (Cîmpulung–Moldovenesc, Pojorîta area, Suceava, Romania), Early Aptian; Zljebine (East Serbia), Early Aptian.
Holocystis dupini (d'Orbigny, 1850)
emend, de Fromentel, 1861
Figure 1d
* 1850 Tetracoenia Dupiniana d'Orbigny, p. 121.
. 1851 Tetracoenia Dupiniana; d'Orbigny, p. 166, fig. 303.
. 1861 Holocystis Dupini; de Fromentel, p. 308.
. 1987 Holocystis bucowinensis Volz; Kuzmicheva, p. 222, pi. 1, fig. 3.
. 1988 Holocystis bukowinensis Volz; Kuzmicheva and Aliev, p. 157, pi. 1, fig. 4.
v. 1995 Holocystis calzadai n. sp.; Reig Oriol, p. 8, pi. 1, fig. 1.
2000 Nowakocoenia cieszynica gen. et sp. nov.; Kolodziej and Gedl, p. 187, figs 3, 8–9.
Type. The type material could not be found in the collections of the MNHN.
Dimensions. (d'Orbigny, 1851, p. 166) c: (1.5) 1.7–2.2 mm, ccd: (1.7) 2.0–2.5 (4.0) mm, s: 8–16, sc: 16; other material c: (1.3) 1.5–2.0 (2.2) mm, ccd: 1.7–3.5 mm, s: 16, sc: 16.
Description. Small flat colonies with a sub–cerioid to plocoid appearance. The distance between calices may vary giving a different appearances to the calicular arrangement. Calicular outline circular, in places irregular. Septa straight, free and compact. Three septal cycles are developed, the first reaching ca. 40 percent of the calicular diameter, the second reaching up to 20 percent, and the third remaining very short, only marked by vertical ridges in calices. In small calices the third cycle may be absent or incomplete. The length of septa may vary. They are much shorter in specimens described as Nowakocoenia cieszynica. Sixteen costae can be counted. They are non–confluent to sub–confluent, depending on the calicular distance. The endotheca consists of thin tabulae; the exotheca of dissepiments.
Remarks. D'Orbigny (1850) gave only a very short description. A good illustration is provided in d'Orbigny (1851, p. 166). Since the work of Felix (1914, p. 25), H. dupini is considered a junior synonym of H. elegans. This cannot be confirmed because H dupini has smaller calicular dimensions.
D'Orbigny (1850, p. 121) indicated two localities for the new species: Les Croûtes (Dépt. Aube, France), and Seignelay (Dépt. Yonne, France), both without stratigraphical data. An Early Aptian age was claimed subsequently (Corroy, 1925, p. 456). Early Aptian coral–bearing sediments were never confirmed in later literature (e.g., Chevalier and Masse, 1980) from the Paris Basin. No coral specimen belonging to Holocystis could be found in the collections or during extensive excavations by the author in the Dépt. Yonne (Löser, 2001).
The material described by Kolodziej and Gedl (2000, p. 187) differs from the other material by having a very reduced coenosteum. However, the width of the coenosteum may differ in one and the same colony as it can be recognised for instance in H. nomikosi and might not be a specific character. Additionally, the number of septa is greater; the beginning of a fourth cycle can be observed in Nowakocoenia cieszynica.
Occurrence. Can Grau (Mun. Sant Martí Sarroca, Com. Baix Penedés, Tarragona, Cataluña, Spain), Late Aptian, MGSB 55820 (holotype of Holocystis calzadai); Cerro de Oro (Municipio Ures, Sonora, Mexico), Late Barremian–Early Aptian, ERNO 3032,3023 (from BZl), ERNOL–4378 (from CO4).
Occurrences according to the literature. Small Balean (Krasnovodskaya obi., Turkmenistan), Early Barremian; Alikuliushagi (Kubatlinskij r, Azerbaijan), Barremian; Rudzica (Bielsko–Biala, Slaskie, Poland), Late Barremian–Early Aptian.
Holocystis elegans (Lonsdale, 1849)
Figures 1e–g
1847 Astreal elegans Fitton, p. 296 [nomen nudum, non Astrea elegans Goldfuss, 1826].
* v 1849 Cyathophoral elegans; Lonsdale, p. 83, pi. 4, figs 12–15.
. 1851 Holocystis elegans; Milne–Edwards et al., p. 70, pi. 10, fig. 5.
. 1851 Holocystis elegans; Bronn, p. 143, pi. 29.4, fig.13.
1860 Holocystis elegans; Milne–Edwards, (3), p. 326.
? 1861 Holocystis elegans E. H.; Giimbel, p. 566.
. 1879 Holocystis elegans; Nicholson, (1), p. 211, fig. 102
. 1879 Holocystis elegans; Quenstedt, p. 454, pi. 158, fig. 20.
non 1883 Tetracoenia elegans; de Fromentel, p. 520, pi. 139, fig. 1 [=Actinastrea sp.].
. 1927 Holocystis elegans (Fitton); Ryder, p. 120, figs 1–9
v ? 1964 Holocystis bukowinensis Volz; Morycowa, p. 25, pl. 3, fig. 3, pi. 5, fig. 1.
v ? 1989 Holocystis bukowinensis Volz; Morycowa, p. 61, pl. 19, fig. 2.
v non 1998 Holocystis elegans (Lonsdale ,1849); Schollhorn, p. 76, figs 36, pi. 18, figs 4–5, pi. 21, figs 1–3, pi. 27, fig. 1 [=Holocystis octolobata].
v. 2003 Holocystis elegans (Lonsdale, 1849); Baron–Szabo and González–León, p. 204, figs 6A–B.
Holotype by monotypy. NHM R. 50224 [46800], Lower Greensand, Early Aptian, Atherfield (Isle of Wight, United Kingdom).
Dimensions. Holotype c: 2.2–2.8 mm, ccd: 3.0–3.5 mm, s: 8–16, sc: 16; other material c: (2.0) 2.5–3.0 (3.2) mm, ccd: (2.8) 3.0–3.5 (5.0) mm, s: 8–16, sc 16.
Description. Small rounded colonies. In the type locality they are commonly attached to bivalve shells. The colony appears cerioid, but there is a narrow coenosteum. The calicular outline is round, in small calices quadratic. Septa compact, straight and free. They occur in three cycles. The four septa of the first cycle reach up to 40 percent of the calicular radius. Those of the second cycle are much shorter, reaching only up to ten percent of the radius or only being marked by ridges in the calice. Eight septa of the third cycle are only visible as ridges. Sixteen costae are developed. The peritheca is made of dissepiments and the confluent to subconfluent costae. The endotheca consists of very regular tabulae and some dissepiments. Approximately four tabulae occur within two millimetres of vertical section.
Remarks. Lonsdale used Fitton as author of the type species. Although Fitton (1847, p. 296) was the first author to use the name Astrea? elegans, he mentioned only the name. The first description and illustration was given by Lonsdale (1849, p. 83).
Corroy (1925) mentioned the species from various outcrops of the Early Hauterivian and Early Aptian of the Paris Basin, but his collection could not be found, and no material belonging to Holocystis was found from the indicated localities.
The species does not occur at the locality Cerro de la Espina (Sonora, Mexico) as indicated by Baron–Szabo and González–León (2003, p. 204). The original sampling list and the material were both in hand and indicate that all Holocystis samples described by Baron–Szabo and González–León (2003) are from the locality Cerro de Oro (Sonora, Mexico; sample locality BZ1).
The material mentioned by Giimbel (1861, p. 566) could not be found in the collection of the BSP A recent revision of the corals from the Allgau area (Baron–Szabo, 1997) did not report the occurrence of Holocystis, and also the author's own excavations in the outcrop area did not yield any Holocystis.
The material depicted by Morycowa (1964) as Holocystis bukowinensis shows a slightly larger calicular diameter and the beginning of a fourth septal cycle. The largest measured calicular diameter is 4.1 mm, the highest number of septa 24. These values make its assignment even to the species with the largest dimensions (Holocystis elegans) unsure.
Occurrence. Atherfield (Isle of Wight, United Kingdom), Early Aptian, NHM R. 50224 [46800] (holotype); Atherfield point (Isle of Wight, United Kingdom), Early Aptian, BSP 2003 XX 5431, 6044–6047, GPS FLX 2092; Cerro de Oro (Municipio Ures, Sonora, Mexico), Late Barremian–Early Aptian, ERNO 3018 (from BZ1), ERNO L–4313, L–4317, L–4324 (from CO4); Cerro de Oro (Municipio Ures, Sonora, Mexico), Early Albian, ERNO L–4339, L–4344, L–4345, L–4349, L–4380 (from CO5); Sierra de Sopalmo, Early Aptian (JCSV C3).
Occurrence unsure. Jastrzebia (Lanckorona, Malopolskie, Poland), Early Aptian, UJ 4P nn.
Occurrence according to the literature. Rye, Peasemarsh (Sussex, United Kingdom), Early Aptian; Coast–Guard Station; Atherfield, Redhill cutting; Sandownbay, Shanklin (all Isle of Wight, United Kingdom) Early Aptian.
Holocystis octolobata Kuzmicheva, 2002
Figures 1h–i
v. 1998 Holocystis elegans (Lonsdale 1849); Schollhorn, p. 76, fig. 36, pi. 18, figs 4–5, pi. 21, figs 1–3, pi. 27, fig. 1.
* 2002 Holocystis octolobata Kuzmicheva, p. 166, pi. 23, figs 4–5.
Type. Holotype PIN 4768/127 (by original designation), Barremian, Akkyr (Turkmenistan).
Dimensions, c: (1.8) 2.2–3.0 (3.2) mm, ccd: 2.5–.0 mm, s: 4–8.
Description. Small rounded colonies with calices on all sides. Calicular arrangement sub–cerioid to plocoid. The calices have a quadratic form. The compact, straight and free septa appear in two cycles. The septa of the first cycle are long (half of the radius of the calice), those of the second cyle are very short and in some cases hardly recognisable. Eight costae canbe found. The endotheca is well developed and consists of thin tabulae, which are slightly convex. Approximately six tabulae can be counted at a distance of two millimetres in a vertical section. The exotheca consists of dissepiments.
Remarks. The description and illustration differ from each other in Kuzmicheva (2002, p. 166). In the text, the calicular diameter is indicated as 3.0–3.5 mm, the illustrations show a diameter of 2.2–3.0 mm (fig. 4) and 2.8–3.2 mm (fig. 5). It is not clear whether H. octolobata represents an independent species or is synonymous with H. elegans. In its dimensions, it does not differ much from the latter; only the lower number of septa is a clear difference.
Occurrence. Senyus section (Mun. Cabo, Com. AltUrgell, Lérida, Cataluña, Spain), early Late Aptian, BSP 2003 XX 4052; El Caso section (Set Comelles, Mun. Coll de Nargó, Com. Alt Urgell, Lérida, Cataluña, Spain), latest Aptian, BSP 2003 XX 4023.
Occurrence according to the literature. Akkyr (Turkmenistan), Barremian.
Holocystis nomikosi sp. nov.
Figures 1j–1
v. 1926 Cyathophora sp.; Dietrich, p. 66, pi. 6, fig. 3.
Derivation of name. After D. Nomikos, the owner of the tomato puree company where the type locality is located.
Holotype. BSP 2003 XX 6200.
Paratypes. BSP 2003 XX 6172, 6173, 6199, 6207, 6215.
Type locality. North of the Chiarmena SE Aliartos, on the grounds of the tomato puree company of D.Nomikos, located at the eastern end of Aliartos on the road to Thiva, (Viotia, Greece); WGS84 position Lat 38°22'50" N, Long 23°8'25" E. Evangelistria–conglomerates of Late Aptian–Early Albian age based on the occurrence of the microfossils Pseudocyclammina vasconica, Trocholina aptiensis, Mesorbitolina texana and Simplorbitolina conulus (see Löser and Raeder, 1995).
Material. Holotype with two thin sections, and five paratypes from the type locality. One specimen from Tanzania.
Diagnosis. Holocystis with a calicular diameter of 1.5–1.9 mm and a number of septa of 10–14. The third cycle is always present, but never completed.
Dimensions, c: 1.5–1.9 (2.0) mm, ccd: 2.0–2.5 (3.2) mm, s: 10–14, sc: 16.
Description. Small rounded colonies (20–30 mm diameter). The calicular arrangement appears cerioid at the outer surface. The thin section shows that each calice has its own wall, and a narrow coenosteum occurs. The outlines of calices are round or slightly quadratic. They are regular in size, but larger calices occur. Septa are thin, straight, free and compact. They appear in three cycles. The first cycle consists of four septa, reaching almost to the centre of the calices (ca. 80 per cent of the radius of the calice). The septa have a slightly broader peripherical part, therefore appearing wedge–shaped. Four septa of the second cycle are short (ca. 20 per cent of the radius of the calice). The third cycle with up to six septa is only developed as ridges on the inner calicular wall. Normally, only two septa of the third cycle are visible in the calices. There are invariably 16 costae. The coenosteum is made of confluent or subconfluent costae and dissepiments, which extend from the calice. The endotheca consists of tabulae and disspiments. Both are thin, and dissepiments dominate. About six to eight endothecal elements occur in two millimetres of vertical section.
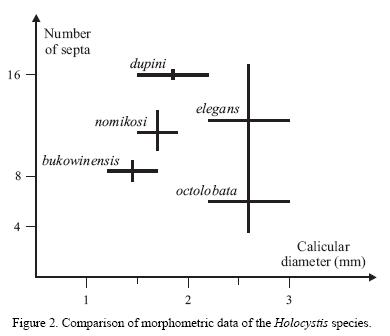
Comparison and remarks. H elegans has larger dimensions. H. dupini shows a higher calicular diameter and a completed third cycle, at least in mature calices, whereas the third cycle is in no case complete in the new species. H. bukowinensis is the most similar species, but shows a slightly smaller calicular diameter and a smaller number of septa (Figure 2).
The material described by Dietrich (1926, p. 66) is only well preserved on the surface; the inside is completely recrystallized and does not show any details.
Occurrence. Pilepile, Nambawala plateau (Mtwara, Tanganyika, Tanzania), Late Aptian, MB K1398; Chiarmena SE Aliartos, Aliartos (Viotia, Greece), Late Aptian–Early Albian, BSP 2003 XX 6172, 6173, 6199, 6200, 6207, 6215.
Stratigraphical range. Late Aptian–Early Albian.
Questionable species
Holocystis polyspathes Giimbel, 1861 was only once listed (Giimbel, 1861, p. 566) and not described or depicted afterwards. The type material could not be found at the B SP. The species must be considered nomen nudum. In a recent revision (Baron–Szabo, 1997) of the corals from the Allgau area, Holocystis was not indicated.
The following species were previously or currently assigned to Holocystis, but belong to other genera.
The holotype (by monotypy, NHMW, without number) oí Holocystis similis Toula, 1877 from Kama (Sveti–Nikola–Balkan, East Serbia) with Early Aptian belongs to the genus Hydnophoraraea.
The holotype (by monotypy, NHMW 1998z0054/0001) oí Holocystis tenuis Toula, 1878 from Vratsa (Vrachanska oblast, Bulgaria) with Late Barremian to Early Aptian was originally assigned to Holocystis, but correctly reassigned to the genus Felixigyra by Masse and Morycowa (1994).
DISCUSSION
Stratigraphical distribution
The genus Holocystis is limited to the Barremian to Early Albian (Figure 3). It appeared with three species in the Barremian, had its maximum of four species in the Late Aptian, and disappeared in the Early Albian, with only two species. The temporal distribution reflects the general abundance of corals in the Cretaceous (Löser, 1998a, fig. 1): increasing values until the Early Aptian and a slight drop in the Early Albian. The sudden disappearance of the genus is probably linked to global anoxic events during the Albian. Almost no hermatypic coral faunas are known from the Middle Albian, and numerous genera disappeared together with Holocystis before the Cenomanian (Löser, 2005). Middle and Late Albian faunas with corals are less numerous in species and are dominated by solitary, most likely ahermatypic corals. Such faunas are mainly known from Texas (USA; Wells, 1933). Colonial coral faunas from the Middle to Late Albian are known from southern England (Jukes–Browne, 1900), southern India (Stoliczka, 1873), northern Spain (Reitner, 1987) and New Mexico (USA; Turnsek et al., 2003). They do not contain Holocystis.

Palaeobiogeography
The genus is known from only three large regions: the central Tethys, the Caribbean and the southern Tethys (Figure 4). It is unknown from the Tibetan or Japanese faunas. There is no regular pattern in the palaeobiogeographic distribution of the genus. In the majority of the localities, two species co–occur, but there is no repeating combination pattern that could indicate that those two species are synonymous.
The occurrence oí Holocystis in the New World seems problematic: it is restricted to two different levels (Late Barremian–Early Aptian Cerro de Oro Formation, Early Albian Cerro La Espina Member) within a geographic distance of less than one kilometre within one locality (Cerro de Oro) in Sonora, Mexico. There are numerous outcrops within the Cerro La Espina Member (Mural Limestone) with various coral faunas, but Holocystis could not be found in any of these faunas. A confusion of samples can be excluded, since samples were also collected recently by the author. Both formations are well distinguished in a tectonically relatively calm region. Between both levels is a succession of clastic sediments with a time span of about five Ma. A local persistence –the only explanation of this phenomenon– of coral species over a relatively long time span is rarely observed due to the rarity of sections with a comparable (coral bearing) facies over such a long time period. Comparable situations where species persisted, although rare elsewhere, are known from the Early and Late Cenomanian (Dixoni zone, Geslinianum zone; Löser, 1989) of Germany (Colonicyathusgeinitzi,Mesomorpha minima, Stylohelia granúlala), or from the Early and late Late Aptian of Spain (Schollhorn, 1998; cf. Comalia sp., Holocystis elegans). In the latter region, again a Holocystis species persisted over a longer time span in the same region.
Palaeoecology
The genus occurred preferably in a siliclastic or at least siliciclastic–influenced facies type. The type locality of Holocystis elegans, the Perna Beds of the Isle of Wright, is marked by sandstones and sandy clays. Localities in Greece are all in the Evangelistria conglomerates, a clastic sediment with coarse ophiolite pebbles as components. The material from Spain and Mexico occurs in argillaceous limestones. The material from Poland comes from conglomeratic sand stones. The isolated colony from Tanzania shows a pale white colour but no attached sediment, and could therefore come from a carbonate facies. Samples from the same locality, for which an Urgonian facies was claimed by Dietrich (1926), show different matrix, such as limestones and sandstones. The unique specimen from Italy was found in carbonatic sediments. Holocystis can be found in localities poor in species (such as Can Grau in Spain, Atherfield in the United Kingdom) and in localities rich in species, such as the outcrops in Greece, Mexico, Poland, Romania, Serbia or the localities in Lérida (Spain). Where they occur, coral colonies of the genus are not rare. At least the outcrops in Aliartos (Greece), near Atherfield (United Kinddom), close to Montsec de Rubies (Spain), and in Sonora (Mexico) have yielded numerous specimens, whereas other species are less abundant.
Evolution
Traditionally, the suborder Stylininais subdivided into three families: Stylinidae d'Orbigny, 1850, Cyathophoridae Vaughan and Wells, 1943, and Agathehidae Beauvais and Beauvais, 1975. Other families based on genera currently assigned to the suborder Stylinina (such as Euheliidae, Pseudocoeniidae, Cladophylliidae, Stylosmiliidae) comprise only one genus, which would not be very helpful for classification. Whereas the Agathehidae differ by their irregular septal symmetry and the formation of the coenos–teum, the Stylinidae and Cyathophoridae are distinguished by the presence or absence of a columella. Auriculae are present in the Stylinidae, but in the Cyathophoridae they are generally (not always) absent. This feature can also be linked to the presence of the columella. Members of the two families also differ in the size of their coenosteum (small in Cyathophoridae, large in Stylinidae), the number of costae (in Stylinidae they are often more numerous) and the wall formation (parathecal in Cyathophoridae, para–septothecal in Stylinidae).
Another concept for distinguishing the two families is put forward in Pandey et al. (2002) but not confirmed by appropriate type material and therefore doubtful. Both families comprise genera that form plocoid corals with a regular septal symmetry, using different systems (Cryptocoenia, Holocystis, Pentacoenia, Pseudocoenia in the Cyathophoridae; A canthocoenia, Enallhelia, Heliocoenia, Stylangia, Stylina in the Stylinidae). In both families, corals may show a tetrameral, pentameral, hexameral, octomeral ordecameral symmetry. Genera within the Cyathophoridae are traditionally distinguished by their symmetry, while in the Stylinidae all plocoid forms are referred to as Enallhelia, Heliocoenia, Stylangia, Stylina, depending on the growth form and the form of the columella. This does not apply, however, to Acanthocoenia, the genus with a columella and a pentameral symmetry.
Cyathophoridae and Stylinidae both originated in the Middle Jurassic (Lathuiliére, 1989); their plocoid colonies show hexameral, octomeral and decameral symmetries (e.g., Beauvais, 1964). Species older than Early Cretaceous with a tetrameral or pentameral symmetry are unknown in both families. Species with a pentameral symmetry suddenly appeared in both families in the Hauterivian, and those with a tetrameral symmetry in the Barremian (Figure 5). However, this experiment did not last long. In the Albian, the tetrameral and pentameral members disappeared in both families. The octomeral species became moreover extinct in both families at the end of the Aptian. With the exception of the decameral forms, genera showing the same symmetry also died out in both families at the same time. In both families, species with a hexameral symmetry are much more common than species with a septal symmetry not based on six, and pentameral and tetrameral species are few and far between.

The formation of the columella is, at least in the Stylinidae, a constant feature. Though its form may vary within one colony between lamellar and styliform (making the distinction between Heliocoenia and Stylinia a rather vague one), its presence or absence is a stable characteristic. Species of Cyathophoridae and Stylinidae may occur together, but none of them have the same calicular diameter and the same number of septa, differing only in the presence or absence of the columella. The two families are considered two different phylogenetic lines.
The surprising parallelism in the sudden variation of the basic number of septal symmetry in the early Cretaceous Cyathophoridae and Stylinidae is not considered just an experiment but is thought to relate to environmental conditions. The growth form of the colony, the size of the calices, the septal perforation and the formation of pennulae have been thoroughly discussed as being indicators of environmental changes (e.g., Dupraz and Strasser, 2002; Sanders and Baron–Szabo, 2005), but changes in the septal symmetry as a possible reaction have been unknown up to now.
ACKNOWLEDGEMENTS
I am indebted to the curators of the above listed museums, and university institutes who allowed me to study type material, as well as to José Carlos Saldaña–Villodre (San Vicente del Raspeig) for showing me material from his private collection. Fieldwork in England, France, Greece, and Spain, as well as sample preparation, was supported by DFG projects FL 42/73 and LO 769/2. Fieldwork in Mexico was supported by project PAPIIT DGAPA IN 107803. Carlos González León (Hermosillo, Mexico), Elmar Schollhorn (Mascefa, Spain), and Thomas Steuber (Bochum, Germany) introduced me to outcrop areas in Mexico, Spain, and Greece. Many thanks to Elzbieta Morycowa and Bogdan Kolodziej (both Kraków) for providing photographs of material from Poland for comparison. Valuable comments by Jacob Leloux (Leiden), Elzbieta Morycowa (Kraków) and Dragica Turnsek (Ljubljana) helped to improve the manuscript. For grammatical correction I would like to thank Gregory Webb (Brisbane) very much.
REFERENCES
Alloiteau, J., 1952, Embranchement des coelentérés, in Piveteau, J. (ed), Traite de Paléontologie: Paris, Masson, 1, 376–684. [ Links ]
Baron–Szabo, R.C., 1997, Die Korallenfazies der ostalpinen Kreide (Helvetikum: Allgauer Schrattenkalk; Nordliche Kalkalpen: Brandenberger Gosau) Taxonomie, Palokologie: Zitteliana, 21, 3–97. [ Links ]
Baron–Szabo, R.C., González–León, CM., 2003, Late Aptian–Early Albian corals from the Mural Limestone of the Bisbee Group (Tuape and Cerro de Oro areas), Sonora, Mexico, in Scott, R.W. (ed.) Cretaceous Stratigraphy and Paleoecology, Texas and Mexico: Society for Sedimentary Geology (SEPM)–Gulf Coast Section Foundation, Special Publications in Geology, 1, 187–225. [ Links ]
Beauvais, L., 1964, Etude stratigraphique et paléontologique des formations a madréporaires du Jurassique supérieur du Jura et de l'Est du Bassin de Paris: Mémoires de la Société géologique de France, 43(100), 1–287. [ Links ]
Beauvais, L., Beauvais, M., 1975, Une nouvelle famille dans le sous–ordre des Stylinida Alloiteau: les Agatheliidae nov. fam. (Madréporaires mésozoiques): Bulletin de la Société géologique de France, serie 7, 17(4), 576–581. [ Links ]
Bourne, G.C., 1900, The Anthozoa, in Lankester, R. (ed.), Treatise on Zoology. Part 2: Porifera and Coelenterata: London, A. & C. Black, 59–79. [ Links ]
Bronn, H.G., 1851, Lethaea geognostica (2, 3:) Meso–Lethaea (5, 4:) Kreide–Gebirge: Stuttgart, Schweizerbart, 412 pp. [ Links ]
Chevalier, J.–P, Masse, J.P, 1980, Apercu sur les formations récifales en France: Geobios. Mémoire Special, 4, 7–16. [ Links ]
Corroy, G., 1925, Le Néocomien de la bordure oriéntale du Bassin de Paris: Bulletin de la Société des Sciences naturelles de Nancy, 2(4), 171–506. [ Links ]
Dietrich, W.O., 1926, Steinkorallen des Malms und der Unterkreide im südlichen Deutsch–Ostafrika: Palaeontographica, suppl.7(l), 43–62. [ Links ]
Duncan, P.M., 1867, A monograph of the British fossil corals, ser. 2, 4. Corals from the zones of Ammonites planorbis, angulatus, Bucklandi, obtusus, and raricostatus of the Lower Lias; from the zones of Jamesoni and Henleyi of the middle lias; and from the aviculacontorta zone and the white lias: Palaeontographical Society Monographs, 20, 1–43. [ Links ]
Dupraz, C, Strasser, A., 2002, Nutritional Modes in Coral –Microbialite Reefs (Jurassic, Oxfordian, Switzerland): Evolution of trophic structure as a response to environmental change: Palaios, 17(5), 449–471. [ Links ]
Eguchi, M., 1951, Mesozoic hexacorals from Japan: Science Reports of the Tohoku Imperial University, ser. 2, Geology, 24, 1–96. [ Links ]
Felix, J., 1914, Anthozoa Cretácea: Fossilium Catalogus Animalia, 5–7, 1–273. [ Links ]
Fitton, W.H., 1847, Astratigraphical account ofthe section from Atherfield to Rocken End, on the south–west coast of the Isle of Wight: Quarterly Journal ofthe Geological Society of London, 3, 289–327. [ Links ]
Fromentel, E. de, 1861, Introduction a l'étude des polypiers fossiles: Mémoires de la Société d'émulation du Doubs, 5, 1–357. [ Links ]
Fromentel, E. de, 1883, Zoophytes, terrain crétacé (12), in d'Orbigny, A. (ed.), Paléontologie francaise: Paris, Victor Masson, t. 8, 513–528. [ Links ]
Fromentel, E. de, 1884, Zoophytes, terrain crétacé (13), in d'Orbigny, A. (ed.), Paléontologie francaise: Paris, Victor Masson, t. 8, 529–560. [ Links ]
Gill, G.A., 1977, Essai de regroupement des Stylinides (hexacoralliaires) d'aprés la morphologie des bords internes de leurs septes: Mémoires du Bureau de Recherches Géologiques et Miniéres, 89, 283–295. [ Links ]
González–León, C. M., Lucas, S.G., 1995, Stratigraphy and paleontology ofthe early Cretaceous Cerro de Oro Formation, central Sonora: Geological Society of America, Special Paper, 301, 41–47. [ Links ]
Gümbel, C.W., 1861, Geognostische Beschreibung des Bayerischen Alpengebirges und seines Vorlandes, in Gümbel, C.W. (ed.), Geognostische Beschreibung der Konigreichs Bayern, Abtheilung 1: Gotha, J. Perthes, 950 pp. [ Links ]
Jukes–Browne, A.J., 1900, The Cretaceous rocks of Britain, 1. The Gault and the Upper Greensand of England: Memoirs of the Geological Survey of the United Kingdom, 1–499. [ Links ]
Kolodziej, B., Gedl, E., 2000, Nowakocoenia cieszynica gen. et sp. nov. (Scleractinia) and its Barremian–Aptian age based on Dinocysts (Polish Outer Carpathians): Annales Societatis Geologorum Poloniae, 70, 181–192. [ Links ]
Kuzmicheva, E.I., 1987, Corals from Lower Barremian organogenous buildups in the Malyy Balkhan and Tuarkyr (in Russian), in Amanniyazov, K.N. (ed.), Geologicheskoe stroenie Turkmenistana: Aschabad, Ylum, 217–262. [ Links ]
Kuzmicheva, E.I., 2002, Skeletal morphology, systematics and evolution of the Scleractinia (in Russian): Trudy Paleontologicheskogo Instituta, 286, 1–211. [ Links ]
Kuzmicheva, E.I., Aliev, O.B., 1988, Corals (in Russian), in Ali–zade, A.A., Aliev, G. A, Aliev, M.M., Aliiulla, K., Khalilov, A.G.(eds.), Cretaceous fauna of Azerbaijan: Baku, Azerbaijan, Akademiia Nauk Azerbaidzhanskoi SSR, Institut Geologii, 153–184. [ Links ]
Lathuiliére, B., 1989, Repertoire objectif des coraux jurassiques: Nancy, Presses universitaires, 76 pp. [ Links ]
Lawton, T.F., González–León, CM., Lucas, S.G., Scott, R.W., 2004, Stratigraphy and sedimentology of the upper Aptian–upper Albian Mural Limestone (Bisbee Group) in northern Sonora, Mexico: Cretaceous Research, 25, 43–60. [ Links ]
Lonsdale, W., 1849, Notes on fossil Zoophytes found in the deposits described by Dr.Fitton in his Memoir entitled ,,A stratigraphical account of the section from Atherfield to Rocken End": Quarterly Journal of the Geological Society of London, 5, 55–103. [ Links ]
Löser, H., 1989, Die Korallen der sachsischen Oberkreide, 1. Hexacorallia aus dem Cenoman: Abhandlungen des Staatlichen Museums für Mineralogie und Geologie zu Dresden, 36, 88–154. [ Links ]
Löser, H., 1998a, Cretaceous corals –state of knowledge and current research: Zentralblatt für Geologie und Palaontologie, 11/12 (1996), 1475–1485. [ Links ]
Löser, H., 1998b, Adelocoenia versus Pseudocoenia–some rectifications: Fossil Cnidaria & Porifera newsletter, 27(1), 29–32. [ Links ]
Löser, H., 2001, Le site de Valliéres (département de l'Aube, France); résultats préliminaires sur des coraux de l'Hauterivien inférieur (Crétacé): Bulletin annuel de 1'Association géologique de l'Aube, 22, 39–53. [ Links ]
Löser, H., 2005, Stratigraphy of Cretaceous coral genera: Neues Jahrbuch für Geologie und Palaontologie, Abhandlungen, 238, 231–277. [ Links ]
Löser, H., 2006, Morphology, taxonomy and distribution of the Cretaceous coral genus Paronastraea (Barremian–Cenomanian; Scleractinia): Rivista Italiana di Paleontología e Stratigrafia, 112(1), 131–121. [ Links ]
Löser, H., Decrouez, D., 2000, Stratigraphy of selected Cretaceous coral localities in Northern Spain: Abhandlungen und Berichte für Naturkunde und Vorgeschichte, 21, 63–71. [ Links ]
Löser, H., Raeder, M., 1995, Aptian/Albian coral assemblages of the Helicon Mountains (Boeotia, Greece); palaeontological, palaeoecological and palaeogeographical aspects: Coral Research Bulletin, 4, 37–63. [ Links ]
Löser, H. and 25 co–authors, 2002, List of Citations, in The Catalogue of Cretaceous Corals: Dresden, CPress, 2, 784 p. [ Links ]
Löser, H. and 28 co–authors, 2005, List of Localities, in The Catalogue of Cretaceous Corals: Dresden, CPress, 3, 366. [ Links ]
Masse, J.P., Morycowa, E., 1994, Les Scléractiniaires hydnophoroides du Crétacé inférieur (Barrémien–Aptien inférieur) de Provence (S.E. de la France). Systématique, stratigraphie etpaléobiogéographie: Geobios, 27(4), 433–448. [ Links ]
Milne–Edwards, H., 1857, Histoire naturelle des coralliaires ou polypes proprement dits, v. 1, 2: Paris, Librairie Encyclopédique de Roret, VIII, v. 1, 326 p., v. 2, 633 p. [ Links ]
Milne–Edwards, H., 1860, Histoire naturelle des coralliaires ou polypes proprement dits, v.3: Paris, Librairie Encyclopédique de Roret, 560 p. [ Links ]
Milne–Edwards, H., Haime, J., 1851, Amonograph of the British fossil corals, 1. Introduction; Tertiary and Cretaceous: Palaeontographical Society Monographs, 3, i–lxxxv, 1–71. [ Links ]
Morycowa, E., 1964, Hexacoralla des couches de Grodziszcze (Néocomien Carpathes): Acta Palaeontologica Polonica, 9(1), 1–114. [ Links ]
Morycowa, E., 1971, Hexacorallia et Octocorallia du Crétacé inférieur de Rarau (Carpathes orientales roumaines): Acta Palaeontologica Polonica, 16(1/2), 1–149. [ Links ]
Morycowa, E., 1989, Class Anthozoa Ehrenberg, 1834, in Malinowski, L. (ed.), Geology of Poland, 3. Atlas of Guide and Characteristic Fossils, 2c. Mesozoic, Cretaceous: Warszawa, Wydawnictwa Geologiczne, 58–67. [ Links ]
Morycowa, E., Masse, J.P., Vilas, L., Arias, C, 2001, Montlivaltia multiformis Toula (Scleractinia) from the Aptian of the Prebetic domain (SE Spain): Revista Española de Paleontología, 16(1), 131–144. [ Links ]
Nicholson, H.A., 1879, Manual of Palaeontology: Edinburgh, W. Blackwood, 2nd ed., 2 vols. [ Links ]
Orbigny, A. d', 1849, Note sur les polypiers fossiles: Paris, Masson, 12 p. [ Links ]
Orbigny, A. d', 1850, Prodrome de Paléontologie stratigraphique universelle des animaux mollusques et rayonnés v. 1, 2: Paris, Masson, v. 1, 394 p., v. 2, 428 p. [ Links ]
Orbigny, A. d', 1851, Cours élémentaire de Paléontologie, 3. Polypiers ou Zoophytes: Paris, Masson, 189 p. [ Links ]
Pandey, D.K., Lathuiliére, B., Fürsich, ET, Kuldeep, S., 2002, The oldest Jurassic cyathophorid coral (Scleractinia) from siliciclastic environments of the Kachchh Basin, western India: Paläontologische Zeitschrift, 76(2), 347–356. [ Links ]
Prever, P.L., 1909, Anthozoa, in Parona, CE, Crema, C, Prever, P.L. (eds.), La Fauna Coralligena del Cretáceo dei Monti d'Ocre neir Abruzzo Aquilano: Rome, R. Comitato geológico del Regno, Memorie per serviré alia descrizione della carta geológica d'Italia, 5(1), 51–147. [ Links ]
Quenstedt, F.A., 1879, Petrefactenkunde Deutschlands, 6. Korallen (Rohren– und Sternkorallen), v. 1: Leipzig, Fues Verlag, 624 p. [ Links ]
Reig Oriol, J., 1995, Madreporarios cretácicos: Barcelona, 62 p. [ Links ]
Reitner, J., 1987, Mikrofazielle, palokologische und palaogeographische Analyse ausgewahlter Vorkommen flachmariner Karbonate im basko–kantabrischen Strike Slip Fault–Becken–System: Documenta naturae, 40, 1–239. [ Links ]
Ryder, T.A., 1927, The structure and development of Holocystis Ed. and Haime: Geological Magazine, 64, 119–122. [ Links ]
Sanders, D., Baron–Szabo, R.C., 2005, Scleractinian assemblages under sediment input: their characteristics and relation to the nutrient input concept: Palaeogeography, Palaeoclimatology, Palaeoecology, 216(1/2), 139–181. [ Links ]
Schollhorn, E., 1998, Geologie und Paläontologie des Oberapt im Becken von Organyà (Nordspanien): Coral Research Bulletin, 6, 1–139. [ Links ]
Stoliczka, F., 1873, The corals or Anthozoa from the Cretaceous rocks of South India: Palaeontologia Indica, Memoirs of the Geological Survey of India, 8(4/5), 130–202. [ Links ]
Turnsek, D., Mihajlovic, M., 1981, Lower Cretaceous Cnidarians from eastern Serbia: Razprave Slovenska akademija znanosti in umetnosti, 23(1), 1–54. [ Links ]
Turnsek, D., LeMone, D.V., Scott, R.W., 2003, Tethyan Albian corals, Cerro de Cristo Rey uplift, Chihuahua and New Mexico, in Scott, R.W. (ed.), Cretaceous Stratigraphy and Paleoecology, Texas and Mexico: Society for Sedimentary Geology (SEPM)–Gulf Coast Section Foundation, Special Publications in Geology, 1, p. 147–185. [ Links ]
Toula, F., 1877, Geologische Untersuchungen im westlichen Theile des Balkans und der angrenzenden Gebiete, 4. Ein geologisches Profil von Osmanich am Arcer, über den Sveti–Nicola–Balkan, nach Ak–Palanka an der Nisava: Sitzungsberichte der Mathematisch–Naturwissenschaftliche Classe der Kaiserlichen Akademie der Wissenschaften, (1) 75, 465–549. [ Links ]
Toula, F., 1878, Geologische Untersuchungen im westlichen Theile des Balkans und in den angrenzenden Gebiete, 5. Ein geologisches Profil von Sofia über den Berkovica Balkan nach Bercovar: Sitzungsberichte der Mathematisch–Naturwissenschaftliche Classe der Kaiserlichen Akademie der Wissenschaften, (1) 77, 247–317. [ Links ]
Vaughan, T.W., Wells, J.W., 1943, Revision ofthe suborders, families and genera of scleractinia: Geological Society of America, Special Paper, 44, 1–363. [ Links ]
Volz, W., 1896, Die Korallen der Trias monographisch bearbeitet: Palaeontographica, 43, 1–127. [ Links ]
Volz, W., 1903, Über eine Korallenfauna aus dem Neokom der Bukowina: Beitrage zur Palaontologie und Geologie Österreich–Ungarns und des Orients, 15(1), 9–30. [ Links ]
Wells, J.W., 1933, Corals of the Cretaceous of the Atlantic and Gulf Costal Plains and W–Interior of the United States: Bulletins of American Paleontology, 18(67), 83–292. [ Links ]
Wells, J.W., 1944, Cretaceous, Tertiary and Recent corals, a sponge and a alga from Venezuela: Journal of Paleontology, 18, 429–447. [ Links ]
Wells, J.W., 1936, The nomenclature and type species of some genera of recent and fossil corals: American Journal of Science, 31, 97–134. [ Links ]
Wells, J.W., 1956, Scleractinia, in Moore, R.C. (ed.), Treatise on Invertebrate Paleontology, F. Coelenterata: Lawrence and Kansas, The University of Kansas and The Geological Society of America, 328–444. [ Links ]
Wery, G., 1954, Contribution a la revision des Madréporaires du Néocomien de la Haute–Marne: Dijon, Université de Dijon, Diplome, 97 p. [ Links ]

