










Servicios Personalizados
Revista
Articulo
 Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkAgricultura técnica en México
versión impresa ISSN 0568-2517
Agric. Téc. Méx vol.35 no.4 México oct./dic. 2009
Artículos
El potencial de agua del sustrato en la germinación de maíces con tolerancia y sensibilidad a la sequía*
The substrate water potential in the drought tolerant and sensitive maize germination
Nikolaos Tsougkrianis1, Cecilia Beatriz Peña–Valdivia1 §, Carlos Trejo López1 y José Domingo Molina Galán1
1 Posgrado en Recursos Genéticos y Productividad, Fisiología Vegetal. Colegio de Postgraduados. Km. 36.5 carretera México–Texcoco. C. P. 56230. Montecillo, Estado de México. Tel. 01 595 95 2 05 00. Ext. 1313. E–mails: catre@colpos.mx, jmolina@colpos.mx.
§Autora para correspondencia:
cecilia@colpos.mx.
* Recibido: Noviembre de 2007
Aceptado: Diciembre de 2009
RESUMEN
El objetivo de este estudio fue caracterizar el efecto del potencial de agua (ΨA) del sustrato en la germinación de las semillas de maíces tolerantes y sensibles a la sequía en condiciones de laboratorio. Se utilizaron los cultivares Cafime O, QPM H519, Tux. Seq. C0 y Zac. 58 O sensibles a la sequía, y los cultivares Cafime SM16, Tux. Seq. C8 y Zac. 58 SM19, tolerantes a la sequía. Las semillas se germinaron en vermiculita con (ΨA) de –0.03 (bien hidratada), –1.28, –1.41, –1.58 y –2.05 MPa (con 6% de humedad), se pesaron cada 12 h y se calculó el porcentaje de agua embebida y la germinación. La imbibición máxima (hasta el momento de germinar) en el ΨA de –0.03 MPa de los siete cultivares fue significativamente mayor respecto a los demás ΨA evaluados y alcanzó entre 40 y 70%, en tiempos variables entre los cultivares, desde 48 h (cv. Caf. Seq.) hasta 170 h (cv. Tux. Seq. C8). Los cultivares Cafime SM16 y Zac. 58 SM19 alcanzaron germinación máxima cuatro veces mayores con el ΨA de –2.05 MPa respecto a sus respectivos cultivares sensibles a la sequía. La tolerancia a la sequía de algunos cultivares, como Cafime SM16, se expresa desde la germinación; aunque esta respuesta contrasta entre los cultivares. Algunos cultivares tolerantes ala sequía reaccionan negativamente al ΨA alto del sustrato (humedad saturante) y alcanzan porcentajes de germinación significativamente mayores cuando el ΨA del sustrato es significativamente menor, entre de –1.28 y –1.58 MPa.
Palabras clave: déficit de humedad, vermiculita, Zea mays L.
ABSTRACT
The objective of this study was to characterize seed germination of drought sensitive and tolerant maize in laboratory conditions. Cafime O, QPM (quality protein maize) H519, Tuxpeño Sequia C0 and Zacatecas 58 O, drought sensitive, and Cafime SM16 Sequia, Tuxpeño Sequia C8 and Zacatecas 58 SM19 Sequia, drought tolerant cultivars, were evaluated. The seeds were germinated in vermiculite with Ψw of–0.03 MPa (well watered), –1.28, –1.41, –1.58 y –2.05 MPa (with 6% of humidity), were weighted every 12 h and the percentage of water absorbed and germination was calculated. The maximum imbibition (up to germination) in the Ψw of –0.03 MPa of all cultivars was significantly major compared with the other Ψw evaluated and it reached between 40% and 70%, in variable times between the cultivars, from 48 h (cv. Caf. Seq.) untill 170 h (cv Tux. Seq. C8). The Cafime SM16 and Zac. 58 SM19 cultivars reached maximum germination four times higher with the Ψw of –2.05 MPa compared with their respective drought sensitive cultivars. The drought tolerance of some drought tolerant cultivars, like Cafime SM16, is expressed since the germination stage; even though this respond contrasts between cultivars. Some drought tolerant cultivars react negatively at the high substrate Ψw and reach germination percentages significantly higher when the substrate Ψw is significantly lower, between –1.28 and –1.58 MPa.
Key words: water deficit, vermiculite, Zea mays L.
INTRODUCCIÓN
Existe una serie de factores que regulan tanto la absorción de agua como la germinación de las semillas. Se han identificado genes que contribuyen al debilitamiento de la pared celular, otros involucrados en el metabolismo energético (Bradford et al. , 2000) y los que pueden jugar un papel importante en la protección contra patógenos de la radícula que está emergiendo durante la germinación (Wu et al. , 2001). Los ácidos giberélico (GA) y abscísico (ABA) son de fundamental importancia en el proceso de germinación (Ritchie y Gilroy, 1998); Koornneef et al. (2002) destacaron la acción antagónica del GA y ABA. Además, el papel del agua es de fundamental importancia en la germinación. Según Vertucci (1989), la cinética de la imbibición de las semillas es un proceso muy complej o ya que involucra dos procesos, adsorción (humedecimiento) y flujo hidráulico, los cuales presentan distinta dinámica y las variables que controlan estos fenómenos cambian durante cada una de estas dos fases. Vertucci y Leopold (1987) demostraron que existe una relación exponencial entre el ΨA y el contenido de agua en semillas de soya (Glycine max L.). Así, la tasa de absorción de agua no se afecta de una manera predecible al manipular el contenido de humedad inicial de la semilla. Vertucci y Leopold (1983) y Hsu (1983) puntualizaron que disminuyendo el contenido de humedad de la semilla, por un lado se logra aumentar el ΨA de la semilla pero, por otro lado, se cambia su permeabilidad.
Se desconoce la reacción de la germinación de las semillas de maíz al ΨA del sustrato, el ΨA del sustrato mínimo requerido para que la germinación se lleve acabo, la relación y contraste de estos factores en las semillas de los cultivares tolerantes y sensibles a la sequía. Esta información puede ayudar a hacer más eficiente la selección y mejoramiento de cultivares tolerantes a la sequía, por la reducción significativa del tiempo requerido y la cantidad de cultivares que podrían ser evaluados en poco tiempo (Sánchez–Urdañeta et al., 2005).
La presente investigación tuvo como objetivo caracterizar la reacción de la germinación de maíces, sensibles y tolerantes a la sequía, al ΨA del sustrato y determinar si en esta etapa inicial del desarrollo se expresan diferencias fisiológicas dependientes de la tolerancia a la sequía.
MATERIALES Y MÉTODOS
Material vegetal
Se utilizaron los cultivares Cafime O, QPM H519, Tux. Seq. C0 y Zac. 58 O sensibles a la sequía, y los cultivares Cafime SM16, Tux. Seq. C8 y Zac. 58 SM19, tolerantes a la sequía provenientes del Colegio de Postgraduados en Ciencias Agrícolas, Centro de Internacional de Mejoramiento de Maíz y Trigo (CIMMYT) e Instituto de Investigaciones Forestales Agrícolas y Pecuarias (INIFAP).
Los cultivares Cafime SM16 y Zac. 58 SM19 fueron generados a partir de los cultivares originales respectivos, Cafime O y Zac. 58 O, por el método de selección masal visual estratificada en suelo con limitada humedad (Molina, 1983). El cv QPM H–519 es un maíz de alta calidad proteínica, resultado de un programa CIMMYT–INIFAP, iniciado en 1996, en el que se generaron híbridos con hasta 100% más lisina y triptofano respecto a los maíces comunes que contienen 39% (Sierra et al., 2001). El cv Tux. Seq. C8 fue obtenido después de ocho ciclos de selección recurrente a partir del cultivar denominado Tux. Seq. C0 (Johnson et al., 1986).
Desarrollo del experimento
Las semillas se pesaron individualmente y colocaron en contenedores de cloruro de polivinilo (PVC, con 40 y 100 mm de diámetro interno y altura, respectivamente) con vermiculita con ΨA de –0.03, –1.28, –1.41, –1.58 y –2.05 MPa (equivalentes a saturación, 10, 9, 8 y 6% de agua). Cada recipiente se cubrió con un fragmento de polietileno negro, fijado con una banda elástica, para mantener constante el ΨA de la vermiculita y permanecieron en oscuridad a 25 ± 1 °C. Cada 12 h se registró el peso individual de las semillas en una balanza analítica (Scientech), con precisión de 0.0001 g. Se calculó el porcentaje de agua embebida, en el tiempo, por las semillas respecto a su propio peso hasta la germinación y se contabilizaron las semillas que iban germinando. Las semillas fueron consideradas germinadas cuando la raíz había alcanzado un centímetro de longitud.
La vermiculita con los ΨA de –0.03, –1.28, –1.41, –1.58 y –2.05 MPa se obtuvo al mezclar vermiculita seca con 100, 10, 8, 9 y 6 partes de agua destilada (p:v), respectivamente. Las mezclas se prepararon 48 h antes de usarse y se mantuvieron en bolsas de polietileno selladas, para que la vermiculita contenida en la bolsa alcanzara un ΨA homogéneo. Al término de este período se tomaron muestras de la vermiculita y se incubaron en cámaras psicrométricas (Wescor C–52, Inc., Utha, USA) por un período de 4 h; luego se determinó el ΨA, con un microvoltímetro (Wescor HR–33T, Inc, Utha, USA) operado en el modo de punto de rocío y conectado a las cámaras. Se evaluaron 12 repeticiones de cada ΨA en muestras de vermiculita utilizadas a lo largo del experimento.
Diseño experimental y análisis estadístico
Se utilizó un diseño experimental completamente al azar con arreglo factorial de tratamientos y tres repeticiones. Cada repetición estuvo constituida por 25 semillas. Los factores (y niveles) fueron: siete cultivares de maíz (Cafime O, QPM H519, Tux. Seq. C0 y Zac. 58 O sensibles a la sequía, y los cultivares Cafime SM16, Tux. Seq. C8 y Zac. 58 SM19) y cinco niveles de humedad (–0.03, –1.28, –1.41, –1.58 y –2.05 MPa).
Los datos obtenidos fueron sometidos a un análisis de varianza y comparación múltiple de medias con la prueba de Tukey, para lo cual se utilizó el paquete estadístico SAS para computadora personal (SAS, 1999–2000, versión 8.1). La representación gráfica de los datos se realizó con el programa SigmaPlot de Jandel Scientific (2004, versión 9), para computadora personal.
RESULTADOS Y DISCUSIÓN
Germinación
Imbibición
Las semillas de los siete cultivares iniciaron la imbibición en las primeras 12 h (imbibición inicial) después de ser transferidas a la vermiculita, con los cinco ΨA evaluados. La imbibición continuó incrementando a tasas dependientes de los ΨA y en general, decayó en los ΨA menores. Así, la imbibición de los siete cultivares de maíz mostró una tendencia con el mejor ajuste de una hipérbola rectangular simple (y= ax/b + x), en la que el incremento decae en el tiempo (Figuras 1 A, B a 4 A, B). Figuras 1, 2, 3 y 4). La imbibición inicial de las semillas fue significativamente mayor en la vermiculita bien hidratada, con ΨA de –0.03 MPa, respecto a la alcanzada en los otros cuatro ΨA. La excepción a esta respuesta la presentó el cv. Cafime O, cuyas semillas embebieron (13%) cantidades de agua estadísticamente equivalentes en la vermiculita con el ΨA mayor y los dos menores (–1.58 y –2.05 MPa). La imbibición inicial de las semillas en la vermiculita bien hidratada varió entre 6.81% para el cv. Tux. Seq. C8 y 31.6% para ambos cultivares Zac.,mientras que en la vermiculita con el ΨA menor alcanzó valores entre 3.0% para el cv. Tux. Seq. C8 y 17.1% en el Tux. Seq. C0 (Figuras 1 A, B a 4 A, B).
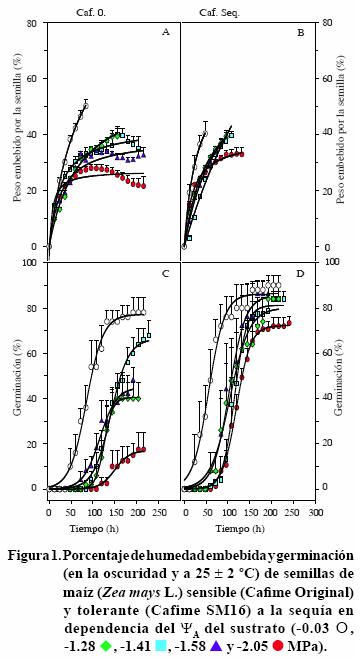

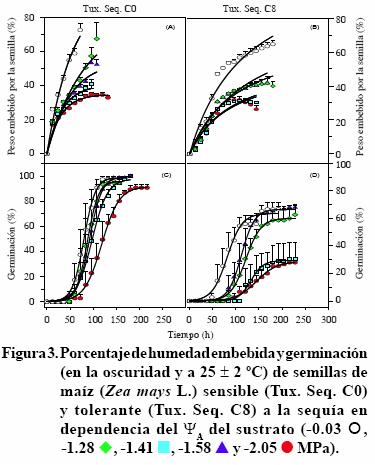
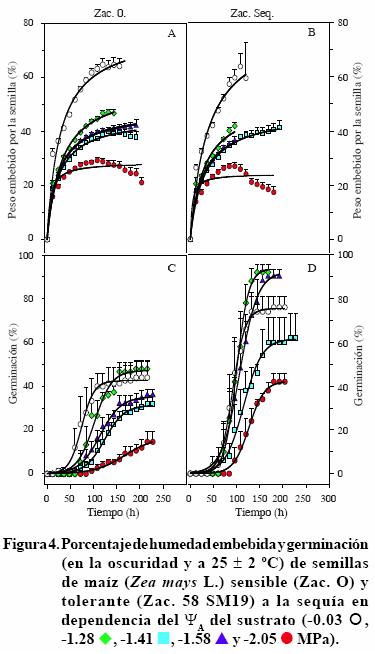
Además de los mayores porcentajes de agua embebida inicialmente, en la vermiculita con ΨA de –0.03 MPa la imbibición máxima (hasta el momento de germinar) de los siete cultivares fue significativamente mayor respecto a los demás ΨA evaluados y alcanzó entre 40% y 70%, en tiempos variables entre los cultivares, desde 48 h (cv Caf. Seq.) hasta 170 h (cv. Tux. Seq. C8). En contraste, en la vermiculita con los cuatro ΨA menores la imbibición máxima fue significativamente menos acelerada que la correspondiente al ΨA de –0.03 MPa respectivo a cada cultivar. Así, la imbibición máxima con el ΨA de –2.05 MPa fue la menor (< 30%) en los siete cultivares (Figuras 1 A, B a 4 A, B). Estos resultados indican que el porcentaje de agua embebida inicialmente por las semillas de maíz es proporcional a la humedad del sustrato y tanto la proporción inicial embebida como la máxima, varían notablemente entre los cultivares de maíz. Al respecto, Vite et al. (2005) evaluaron el efecto del ΨA del sustrato (entre –0.03 y –1.28 MPa) en la germinación de las semillas de frijol silvestre de tres tamaños (0.093, 0.116 y 0.147 g) y observaron que, en promedio los tres grupos de semilla duplicaron su peso en las primeras 24 h; luego, continuaron embebiendo agua a menor velocidad. Pero, la máxima cantidad de agua embebida por las semillas de los tres tamaños fue significativamente mayor en la vermiculita con ΨA de –0.03 y –0.18 MPa (bien hidratada y con 25% de agua, respectivamente). Los mismos autores concluyeron que el tamaño de semilla de frijol silvestre es un carácter que determina el requerimiento de humedad para la germinación y las semillas adaptan su metabolismo para usar eficientemente el agua disponible, cuando hay restricción de ella, durante la germinación.
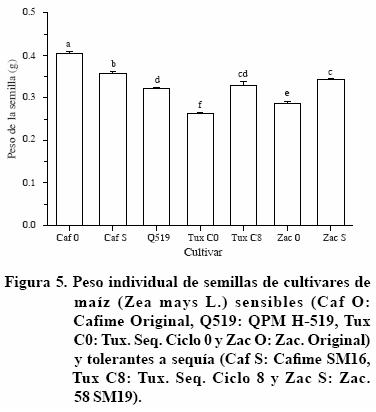
En el caso de los cultivares de maíz utilizados en el estudio presente, se observó que ninguno embebió cantidades de agua equivalentes a las embebidas por el frijol. Además, el peso de las semillas (Figura 5) no mostró alguna relación directa con la capacidad de imbibición; ya que el patrón de imbibición tanto de las semillas más pesadas, como Cafime O y Cafime Seq., como de las más ligeras, como Tux. Seq. C0 y Zac. O, mostró similitudes y contrastes equivalentes a las semillas de los otros tres cultivares. Tampoco se detectó algún patrón de respuesta entre los cultivares originales y los tolerantes a la sequía correspondientes.
Emergencia de la raíz
La exposición de la radícula de los siete cultivares de maíz, en los cinco ΨA, mostró una tendencia con el mejor ajuste aun modelo logístico y= k / 1 + ((k–n/n) e –rx), donde n es el valor de germinación esperado al tiempo x= 0, k e s la asíntota horizontal máxima, o máxima germinación en el tiempo total y r puede ser interpretada como la velocidad media de germinación (SAS, 1991).
La fase inicial, tiempo que tomó la exposición de la radícula, fue variable entre los cultivares y parcialmente dependiente del ΨA de la vermiculita, el incremento acelerado de la germinación fue dependiente tanto del cultivar como del ΨA, y la fase estacionaria final, mostró la germinación máxima de cada cultivar, dependiente al menos parcialmente del ΨA (Figuras 1 C y D a 4 C y D).
La fase inicial más breve correspondió a aquellos tratamientos en los que el ΨA de la vermiculita era de –0.03 MPa, desde 12 h para Cafime SM16 y Zac. O, hasta 36 h para Cafime O, Tux. Seq. C0 y Zac. 58 SM19; en contraste, en general esta fase inicial de exposición de la raíz alcanzó tiempos notablemente prolongados cuando las semillas estuvieron en la vermiculita con los ΨA menores. En estos ΨA la fase inicial de exposición de la raíz de la mayoría de los cultivares tomó alrededor de 100 h (106 hen el cv. Cafime O) en la vermiculita con ΨA de –2.05 MPa (Figuras 1 C y D a 4 C y D). Los cultivares QPM H–519 y Cafime SM16 fueron la excepción de este retraso de la germinación en la vermiculita con el ΨA –2.05 MPa, pues emplearon 48 y 75 h, respectivamente, para iniciar la geminación en este ΨA (Figuras 1 C y 2 B).
Con excepción del cv QPM H–519, la germinación máxima en la vermiculita con ΨA de –2.05 MPa fue significativamente menor que la alcanzada con el ΨA de –0.03 MPa, independientemente de la tolerancia a la sequía del cultivar. La diferencia en la máxima germinación entre los dos ΨA extremos fue variable entre los cultivares; los cultivares que mostraron las diferencias mayores fueron Cafime O (18 y 80% en los ΨA de –2.05 y –0.03 MPa, respectivamente) y Zac. S (42 y 76%).
Los cultivares Cafime SM16 y Zac. 58 SM 19 alcanzaron germinación máxima cuatro veces mayores con el ΨA de –2.05 MPa respecto a sus respectivos cultivares sensibles a la sequía (Figuras 1 C y D, y 2 C y D).
Debe señalarse que algunos de los cultivares no alcanzaron 100% de germinación cuando el ΨA de la vermiculita era –0.03 MPa. Entre esos cultivares sobresalió el Zac. 58 SM 19 debido a que con los ΨA de –1.28 y –1.58 MPa se alcanzaron los porcentajes de germinación significativamente mayores, cercanos a 100%; mientras que con el ΨA de –0.03 MPa la germinación fue menor a 75% (Figura 4D). Este resultado indica que, al menos uno del cultivares tolerantes a la sequía incluidos en el estudio, el Zac. 58 SM 19, es afectado en la germinación por la saturación de humedad del sustrato.
Los resultados del presente estudio muestran que las semillas de maíz, en general, son capaces de germinar con ΨAdel sustrato tan bajo como el generado con sólo 6% de humead (–2.05 MPa). Aunque, en algunos cultivares el porcentaje de germinación fue significativamente bajo. En este sentido, el cv QPM H–5 19 sobresalió, pues aunque con los ΨA menores a –0.03 MPa mostró una fase inicial retardada de hasta 25 h y en la fase de acelerada de la germinación también hubo retrazo, el porcentaje de germinación máxima (= 100%) fue estadísticamente igual en todos los ΨA probados (Figura 2 B). Este resultado puede tener diversas explicaciones. Una de ellas es que en el proceso de mejoramiento que originó al cultivar, indirectamente se seleccionaron genes para tolerancia a la sequía por lo que se alcanza la máxima germinación independientemente del ΨA del sustrato y, además, que este cultivar es uno de los que no son afectados en la germinación por el potencial alto (–0.03 MPa).
En general, los resultados mostraron una cierta relación directa entre la imbibición máxima alcanzada en los diferentes ΨA del sustrato y la germinación máxima alcanzada por cada cultivar. Sin embargo el cv QPM H–519, alcanzó un porcentaje máximo de germinación (100%) independientemente de las diferencias estadísticamente significativas en la imbibición, entre 35 y 70 % (Figura 2 A y B). La germinación máxima de algunos de los cultivares sensibles a la sequía, como Cafime O y Zac. O, mostró un gradiente dependiente directamente del ΨA del sustrato. Así, la germinación máxima alcanzó el porcentaje menor en la vermiculita con ΨA –2.05 MPa.
CONCLUSIONES
La tolerancia a la sequía de algunos cultivares, como Cafime SM16, se expresa desde la germinación; aunque esta respuesta contrasta entre los cultivares. Algunos cultivares tolerantes a la sequía reaccionan negativamente al ΨA alto del sustrato (humedad saturante) y alcanzan porcentajes de germinación significativamente mayores cuando el ΨA del sustrato es significativamente menor, entre de –1.28 y –1.58 MPa.
LITERATURA CITADA
Bradford, K. J.; Chen, F.; Cooley, M. B.; Dahal, P.; Downie, B.; Fukunaga, K. K.; Gee, O. H.; Gurusinghe, S.; Mella, R. A.; Nonagaki, N.; Wu, C.–T.; Yang, H. and Yim, K.–O. 2000. Gene expression prior to radicle emergence in imbibed tomato seeds. In: Black, M.; Bradford, K. J. and Vásquez–Ramos, J. (eds.) Seed biology: advances and applications. Wallingford, UK, CABI International. p. 231– 251. [ Links ]
Hsu, K. H. 1983. A diffusion model with a concentration dependent diffusion coefficient for describing water movement in legumes during soaking. Journal of Food Science 48:618–622. [ Links ]
Johnson, E. C.; Fischer, K. S.; Edmeades, G. O. and Palmer, A. F. E. 1986. Recurrent selection for reduced plant height in lowland tropical maize. Crop Sci. 26:253–260. [ Links ]
Koornneef, M.; Bentsink, L. and Hilhorst, H. 2002. Seed dormancy and germination. Current opinion in Plant Biology 5:33–36. [ Links ]
Molina, G. J. D. 1983. Selección masal visual estratificada. Colegio de Postgraduados, Montecillos, México. 35 p. [ Links ]
Ritchie, S. and Gilroy, S. 1998. Gibberellins: regulating genes and germination. New phytologist 140: 363–383. [ Links ]
Sierra, M. M.; Palafox, A.; Cano, O.; Rodríguez, F. A.; Espinosa, A.; Turrent, A.; Gómez, N.; Córdoba, H.; Vergara, N.; Aveldaño, R.; Sandoval, J. A.; Barrón, S.; Romero, J.; Caballero, F.; González, M. y Betanzos, E. 2001. Descripción varietal de H–519C, H–553C y V–537C. Maíces de alta calidad de proteína para el trópico húmedo de México. INIFAP–CIRGOC. Campo experimental Cotaxtla. Veracruz, México. 21 p. (Folleto Técnico Núm . 30). [ Links ]
Sánchez–Urdaneta, A. B.; Peña–Valdivia, C. B.; Trejo, C.; Aguirre, J. R. and Cárdenas, E. 2005. Root growth and proline content in drought sensitive and tolerant maize (Zea mays L.) seedlings under different water potentials. Cereal Research Communications. 33:697–704. [ Links ]
Vertucci, C. W. and Leopold, A. C. 1983. Dymanics of imbibition in soybean embryos. Plant physiology 72:190–193. [ Links ]
Vertucci, C. W. and Leopold, A. C. 1987. Water binding in legume seeds. Plant physiology 85:224–231. [ Links ]
Vertucci, C. W. 1989. The kinetics of seed imbibition: controlling factors and relevance to seedling vigor. In: Seed moisture. Crop science society of America. CSSA special publication. p. 93–115. [ Links ]
Vite, M. A. C.; Peña–Valdivia, C. B. y Sánchez–Urdaneta, A. B. 2005. Potencial de agua del substrato y tamaño de semilla en la germinación de frijol silvestre (Phaseolus vulgaris L.). Memorias del XVI Congreso Venezolano de Botánica, Maturín, Venezuela. p. 277–279. [ Links ]
Wu, C. T; Leubner–Metzger, G.; Meins, F. and Bradford, K. J. 2001. Class I β–1,3–glucanase and chitinase are expressed in the micropylar endosperm of tomato seeds prior to radicle emergence. Plant Physiology 126:1299–1313. [ Links ]

