










Serviços Personalizados
Journal
Artigo
 Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkAgricultura técnica en México
versão impressa ISSN 0568-2517
Agric. Téc. Méx vol.35 no.3 México Jul./Set. 2009
Artículos
Genetic differences between Anastrepha ludens (Loew) populations stemming from a native and an exotic host in NE Mexico*
Diferencias genéticas entre poblaciones de Anastrepha ludens (Loew) de hospederos nativos y exóticos en el NE México
Víctor Pecina Quintero1§, José Isabel López Arroyo1, Jesús Loera Gallardo1, Juan Rull2, Enrique Rosales Robles1, Edgardo Cortez Mondaca1, Sanjuana Hernández Delgado3, Netzahualcóyotl Mayek Perez3 and Martín Aluja Schuneman2
1 Campo Experimental Río Bravo, INIFAP. A. P. 172, C. P. 88900, Río Bravo, Tamaulipas, México. Tel. 01 899 944 10 46. E– mails: lopez.jose@inifap.gob.mx, loera.jesus@inifap.gob.mx, rosales.enrique@inifap.gob.mx, cortez.edgardo@inifap.gob.mx.
2 Instituto de Ecología, A. C., A. P. 63 C. P. 91000 Xalapa, Veracruz, México. Tel. 01 228 842 18 41. E–mails: juan.rull@inecol.edu.mx, martin.aluja@inecol.edu.mx.
3 Centro de Biotecnología Genómica, Instituto Politécnico Nacional. A. P. 152, C. P. 88730. Ciudad Reynosa, Tamaulipas, México. E–mails: shernandez@ipn.mx, nmayek@ipn.mx.
* Recibido: Febrero, 2009
Aceptado: Septiembre, 2009
§Autor para correspondencia:
pecina.victor@inifap.gob.mx.
ABSTRACT
The objective of this study was to determine the genetic structure of populations of the mexican fruit fly Anastrepha ludens (Loew) in NE citrus growing regions of Mexico. The work was conducted during 2005 at the Center of Genomic Biotechnology in Reynosa, Tamaulipas, Mexico. AFLP markers using four different pairs of initiators were used on specimens collected from the native, ancestral host, yellow chapote (Casimiroa greggii S. Wats) and the exotic host, citrus (Citrus sinensis (L.) Osbeck cv Valencia). The four combinations of initiators amplified an average of 95 AFLP fragments. A total of 382 products were obtained and 259 (67%) of them were polymorphous. The overall index of genetic diversity was 28%, with 3% difference between the genotypes collected from yellow chapote as compared to the genotypes collected from sweet orange. Cluster analysis and principal components showed a close genetic relation among A. ludens specimens regardless of the origin. Although, the dendrogram showed two main groups, the genetic distance varied from 0.0 to 10%, suggesting that despite the reproductive isolation, host groups are not evolving. These results are discussed in the context of evolution and conclude on practical implications of the ongoing eradication program through sterile insect technique and the geographical origin of this species.
Key words: AFLP markers, genetic diversity, host fidelity.
RESUMEN
El objetivo de este trabajo fue determinar la estructura genética de poblaciones de mosca mexicana de la fruta Anastrepha ludens (Loew) en las regiones citrícolas del noreste de México. La caracterización molecular se realizó durante 2005 en Centro de Biotecnología Genómica del Instituto Politécnico Nacional en Reynosa, Tamaulipas. Se utilizaron marcadores AFLP con cuatro diferentes pares de iniciadores para analizar individuos de A. ludens colectados del hospedero nativo chapote amarillo (Casimiroa greggii S. Wats) y del hospedero exótico naranja dulce (Citrus sinensis L. Osbeck cv Valencia). Las cuatro combinaciones de iniciadores amplificaron en promedio 95 fragmentos de AFLP. Se obtuvieron 382 productos de los cuales, 259 (67%) fueron polimórficos. El índice general de diversidad genética fue de 28% con 3% de diferencia entre los genotipos colectados de chapote amarillo y los de naranja dulce. El análisis de conglomerados y de componentes principales mostró estrecha relación genética entre individuos de A. ludens independientemente de su origen. No obstante de que el dendrograma divide las poblaciones de mosca mexicana de la fruta en dos grandes grupos, la distancia genética varió de 0.0 a 10%, lo que sugiere que no obstante el aislamiento reproductivo entre los grupos, estos no están en evolución. Estos resultados se discuten en el contexto de evolución (historia de vida) y se concluye sobre las implicaciones que tienen sobre la campaña en curso para la erradicación mediante la técnica de insectos estériles y para determinar el origen geográfico de estas especies.
Palabras clave: diversidad genética, fidelidad al hospedero, marcadores AFLP.
INTRODUCTION
The genetic structure of A. ludens populations can be an important component for a successful region–wide pest management (Via, 1990; Krafsur, 2005) and it is fundamental to establish the origin and identity of the invasive species into agricultural regions (Roderick, 1996; Armstrong and Ball, 2005). The mexican fruit fly complex is different enough from others that renaming it has been long overdue (Baker et al., 1944; Aluja et al., 2003; Hernández–Ortiz et al., 2004). In the A. fraterculus sibling complex, some sibling species appear to have different geographic range, but others do not. Sibling species exhibit differences in biology, host diversity (Aluja et al., 2000a, 2003) and consistent morphological characteristics (Hernandez–Ortíz et al., 2004). Mating compatibility tests among different populations of South America A. fraterculus have provided evidence of prezygotic reproductive isolation due in part to asynchronies in the daily patterns of sexual activity (Vera et al., 2006). The latter finding has important management implications, particularly on the sterile insect technique (SIT). A. ludens is the most devastating pest of citrus in Mexico and Central America (Aluja, 1993; Thomas, 2003); however, its genetic structure is almost unknown. Up to date the only published report indicate that it is difficult to genetically differentiate A. ludens populations, because it's high intra–population variability (Mangan and Moreno, 2003). Currently, this species are subject to an area–wide management program based on the sterile insect technique that has resulted in the partial/temporary declaration of several fruit fly free areas in NW Mexico (Reyes et al., 2000). Introduced A. ludens populations have been eradicated through SIT from the San Diego area in the US. (Dowell et al., 2000) and have been a recurrent problem in the Rio Grande valley of Texas (Thomas, 2003).
In NE Mexico, the widespread abundance of naturally occurring ancestral hosts of A. ludens such as the yellow chapote (Casimiroa greggi S. Wats), hinders the official declaration of low pest prevalence area; therefore, a stiff regulation on citrus fruit transport has been impose. It is assumed that there is a permanent massive flow of mexican fruit flies from their native hosts (yellow chapote and white zapote [Casimiroa edulis Llave et Lex]) into citrus groves (Enkerlin, 1987), that complicates an effective pest control. However, there is no solid evidence based on the genetic structure of the populations that demonstrates that there is a flow of flies from the native habitats to crop areas. Studies on mobility and longevity of sterile mexican fruit flies revealed that adults can fly up to 9 km; however, 98% were recaptured within 100 m from the release site (Enkerlin 1987, Thomas & Loera–Gallardo, 1998). The latter study determined a mean mobility distance of 240 m (using a regression model), which suggest that mexican fruit flies in citrus, reproduce within the orchard or at nearby neglected plantations. Nevertheless, studies on the reproductive phenology of A. ludens in NE Mexico (Enkerlin, 1987; Thomas, 2003) suggest that the fly is essentially bivoltine in the area and may exploit yellow chapote fruits during spring and citrus at fall. A recent study compared the reaction of wild and lab–reared A. ludens strains to host fruits, the results showed that wild flies were incapable of recognize introduced grapefruit as host, while laboratory flies did. Such reaction is the effect of selective pressures to which the mexican fruit flies have been submitted at the laboratory (Robacker and Fraser, 2004) and a manifestation of the inherent plasticity of this species. Also, when wild and lab–reared flies had previous contact with yellow chapote, the attraction towards this host increased.
The objective of this study was to determine the genetic structure of A. ludens populations from two of the most important citrus producing states of Mexico: Tamaulipas and Nuevo Leon. Molecular characterization of A. ludens populations is important for the successful application of the SIT in pest management programs. If A. ludens, as is the case with A. fraterculus, is found to enclose a complex of cryptic species, then the strain currently being release on SIT programs in Mexico would be useless at certain areas due to reproductive isolation. Furthermore, molecular characterization of A. ludens populations will be an aid to confirm and identify the origin of recurrent putative introductions into citrus growing areas of California, Texas and Florida.
MATERIALS AND METHODS
DNA collection and extraction
A total of 25 specimens (adult and larvae) of the mexican fruit fly, A. ludens were directly collected from infested C. greggii (yellow chapote) and Citrus sinensis cv. Valencia (citrus), in different geographical regions within NE Mexico during 2005. (Table 1). The specimens were preserved in ethanol (90%), then washed with sterilized water (to remove alcohol) and stored at– 80 °C. The specimens were ground to a fine powder with a mortar and pestle. The protocol for DNA extraction was according to Corsini et al., 1999, with extraction solution (50 mM Tris HCl, pH 8.0, 50 mM, EDTA, 3% w/v SDS and 0.1 M 2–mercaptoetanol) and 25 µl proteinase K (10 mg ml).
AFLP analysis
The AFLP protocol utilized was that of Vos et al. (1995). It consists in the genomic DNA digestion and ligation of specific adaptors, followed by two PCR selective amplifications. For DNA digestion a reaction mixture of 30 µL containing 3 µL of DNA (150 ng µL-1), 1.5 µL of EcoRI (Roche®) of 10 U µL-1, 1.5 µL of Tru9I (Roche®) of 10 U µL-1 and 3 µL of 10X RL was used. The samples were incubated for 4 h at 37°C, followed by heating the reaction mixture to 70°C for 15 min to inactivate the restriction enzymes. The specific adapters were united to the sites of restriction. The mixture consisted of 1 µL of EcoRI adapter (5 pmol), 1 µL of MseI adapter (50 pmol), 1.2 µL of 10 mM ATP, 1 µL of 10X RL buffer and 1 µL of T4 DNA ligase (Roche®) of 1 U µL-1 in a total volume of 10 µL. The preamp included 5 µL of the ligation mixture plus the addition of a mixture of oligonucleotides specific AFLP (50 ng µL-1), 2 µL of 10 mM dNTPs in a total volume of 25 µL. The reaction mixture consisted of 0.2 µL addition of Taq DNA polymerase (Roche®) (5 U µL-1) and 5 µL of 10X PCR buffer in a final volume of 20 µL. The total volume of the reaction mixture was 50 µL. The mixture was subjected to PCR amplification in a GeneAmp 9 700 thermocycler (Applied Biosystems®). The following PCR program was used: 94 °C for 30 s, 56 °C for 60 s and 72 °C for 60 s for 20 cycles. Subsequently, 2.5 µL of pre–amplified DNA was added with 0.5 µL of the oligonucleotide EcoRI + 3 (EcoRI + AAA), 0.6 µL of oligonucleotide MseI + 3 (AAA, AAC, ATG and AGG), both with a concentration of 50 ng µL-1, and 0.4 µL of 10 mM dNTPs mix, for a total volume of 5 µL, plus 0.2 µL of Taq DNA polymerase 5 U µL-1 and 1.4 µL of 10 X PCR buffer, for a total volume of 11 µL.
The reaction mixture was subjected to the following PCR program: 94 °C for 30 s, 65 °C for 30 s and 72 °C for 60 s; this for 11 cycles where the temperature was reduced by alignment of 0.7 °C per cycle and then continued with the following PCR program: 94 °C for 30 s, 56 °C for 30 s and 72 °C for 60 s for 23 cycles. Amplified products were separated in acrylamide gel at 6%. Electrophoresis was performed at 2 000 V for 3 h. For detection of amplified products using the Silver Sequence Staining Reagents kit Promega®. The AFLP analysis was conducted at the laboratory of plant biotechnology of the Genomic Biotechnology Center of the National Polytechnic Institute in Reynosa, Tamaulipas, Mexico.
Data analysis
All bands that showed molecular weight over 250 bp were included in the AFLP analysis. It was assumed that bands of the same molecular weight in different individuals were identical. All gels were scored visually for both polymorphic and monomorphic bands. Band presence was indicated by a (1) and absence by a (0). Analysis of molecular variance was used to estimate variance components among and within groups, following Excoffier et al. (1992). Wright's (1951) F statistics were used to estimate the amount of genetic differentiation among populations and subdivisions of populations. Genetic relationship was calculated by the method of Nei and Li (1979), using the S–Plus Version 4.0 (S–Plus, 1997). Principal components (CPs) and cluster analysis were performed. Cluster analysis was performed on the relationship matrices using the unweighted pair group method arithmetic average (UPGMA), (Avise, 1994) and the relationships were graphically presented as dendrograms. Finally, the diversity index was calculated as DI= 1 – p2i, where pi is the frequency of the ith allele, each allele is considered as a unique locus and at the same time, an amplification fragment (Powell et al., 1996).
RESULTS AND DISCUSSION
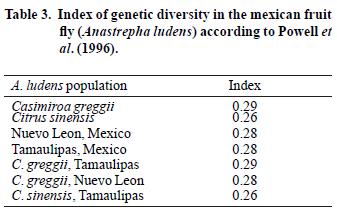
The four–primer combinations amplified among 91 and 99 bands, with a mean of 95 for each pair of primers. A total of 382 products were obtained and 259 of them (67%) were polymorphic (Table 2). A slight difference among hosts was noticed, specimens from yellow chapote showed 3% difference with respect to specimens from citrus (Table 3).
The analysis of molecular variance (AMOVA) showed significant differences (p<0.001) within A. ludens populations (among specimens), while no population differences in regard to host or region were observed. The fixation index (FST) for the three analyzed levels (population, host and region), showed a close relationship among A. ludens specimens in regard to host and region as evidenced by negligible genetic differences among subpopulations. Similarly, cluster analysis and principal components analysis confirmed that there is a strong genetic relationship among specimens of A. ludens (Table 4).
The dendrogram based on the cluster analysis of AFLP data (Figure 1) showed one main group that includes 88% of the specimens, with differences varying from 0.0 to 7.6%. In this main group, a defined grouping pattern was observed: first, genotypes of flies collected from yellow chapote, followed by genotypes from citrus, including the control CITSLP; however, two specimens from yellow chapote (CHANL1 and CHANL2) were mixed with specimens collected from citrus, and two genotypes from yellow chapote (CHATAM6 and CHANL4) clearly differ from the rest.
Due to the observed trend of grouping by host, which suggests the possibility of a marker linked to a fruit fly–host relation, a second cluster analysis was performed, using specimens collected from Tamaulipas and San Luis Potosi (citrus) as control. One thousand bootstrap replicates were obtained from the original data and a distance matrix was calculated for each replicate. In this new analysis, two main genotype groups were observed; the first one, included yellow chapote and citrus specimens from Tamaulipas and the other the citrus specimen from San Luis Potosi (control). The dendrogram showed that all specimens from yellow chapote are closely related, followed by citrus specimens collectedin Tamaulipas (Figure 2).
This suggests the existence of a DNA marker linked to the fruit fly–host relation. Moreover, in the matrices of AFLP, it was observed that the Eco/Mse AAA/AAA pair of primers amplify a DNA band which was present in the whole set of specimens collected in citrus; meanwhile, in the yellow chapote specimens this DNA band was absent, except for one individual.
The AFLP analysis showed a high level of polymorphism (67%), with a mean index of diversity of 28%. The technique was sensitive enough to detect differences among individuals. A slight trend toward greater genetic diversity of A. ludens populations stemming from native yellow chapote was observed, which is possibly due to the fact that these populations have been less perturbed (Robacker and Fraser, 2004) and subjected to a lesser selection than citrus populations subjected to frecuent insecticide sprays.
A weak grouping trend related to host origin was observed; particularly in the specimens from Tamaulipas (Figure 2). The analyses showed a close relation among specimens from the same host with genotypes collected from yellow chapote separated from those collected from citrus.
Phenotypic plasticity has been defined as the ability of a single genotype to produce more than one alternative form in response to environmental conditions (West–Eberhard, 1989). It has been recognized that plasticity itself is a trait subject to natural selection and evolutionary change (Williams, 1966). The results of this research indicate that A. ludens exploiting different hosts have a slightly different genetic composition, probably owing to the fact that citrus colonization by yellow chapote flies implies going through a series of bottlenecks as described by Alberti et al. (1999) for A. fraterculus in Argentina. Nevertheless, the detected differences were not significant, suggesting that reproductive isolation is not evolving among the studied populations. It is important to note that A. ludens is polyphagous and multivoltine (Aluja, 1994; Thomas, 2003). Multivoltine fruit flies have to cope with long periods when host fruit is not available without undergoing diapuase. There are a few possible life strategies to overcome this hurdle. The first one would be through evolution of adult longevity, which has occurred in some Anastrepha species (Aluja et al., 2000a) but not in the case of A. ludens; the second would be by exploiting hosts with long fruiting periods, as it is the papaya fruit fly, Toxotrypana curvicauda Gerstaecker. Multivoltine fruit flies can bridge the fruiting gaps of their main host by exploiting alternate hosts (Aluja and Mangan, 2008). This survival strategy might select for genetic plasticity but also restricts genetic differentiation of host plant races. A likely scenario would explain the observed results, where an A. ludens population switching from chapote to citrus undergoes a genetic bottleneck that can change allele frequency, followed by a return to the ancestral host that may result in homogenizing the gene pool. This life strategy and the phenotypic plasticity observed may help to explain the findings of Robacker and Fraser (2004) showing that mexican fruit flies with previous contact with yellow chapote will prefer this host over other alternatives regardless of their own origin. It also provides support to the idea that chapote populations move back and forth from chapote to citrus as documented by Enkerlin (1987) and more recently by Thomas (2003).
Practical implications of these findings suggest that at a regional scale, the sterile insect technique can be applied for A. ludens without the need to colonize different strains stemming from different hosts. AFLP markers proved to be a useful tool to differentiate among A. ludens specimens allowing to detect a high level of polymorphism, measure genetic diversity and to detect genetic differences in host relation. It is important to mention that this is the first step of several more to follow attempting to characterize A. ludens populations within their geographical range. Molecular scrutiny of these populations at a wider scale is essential for a thorough understanding of the evolution of A. ludens life strategies. Wide range sampling is also critical to rule out geographical effects (Atlantic–Pacific, altiplano–coastal plains) on A. ludens population's genetic composition and to get evidence that these factors may be leading to the evolution of inter population reproductive isolation. That attempt will also contribute to determine the origin of recurrent introductions of A. ludens into citrus growing regions of the U. S., whether occur by natural dispersal (the case of Texas and Nuevo Leon), or human transport. It is also necessary to widen the scope of sampling of host–associated populations considering that both chapote and citrus belong to Rutaceae and A. ludens has been reported infesting plants in very different families: Solanaceae (Thomas, 2004), Anacardaceae, and Rosaceae (Aluja et al., 2000b; Norrbom, 2003). Based on the above, the next step to genetically characterize the A. ludens populations in Mexico, will be to extend the sampling to other regions and other host plants and to incorporate other molecular tools as microsatellites.
CONCLUSIONS
Despite there is a high level of polymorphism (67%) in A. ludens in NE Mexico, the genetic diversity of this species is low (ID 28%).
The AFLP technique was sensitive enough to detect differences among individuals, a silght 3% difference was observed among specimens collected from yellow chapote as compared to those collected from sweet orange.
Cluster analysis and principal components showed a tight genetic relation among A. ludens individual regardless of origin.
Although the dendrogram divided divide the mexican fruit fly populations into two main groups, the genetic distance (0.0 to 10%) suggest that despite the reproductive isolation, host groups are not evolving.
ACKNOWLEDGEMENTS
We thank Victor Maya Hernandez for the collection of flies in Tamaulipas, Mexico and Alberto Anzures for the review of this manuscript. This study was sponsored by INIFAP, Fondo Mixto CONACyT–Gobierno de Nuevo Leon and Fundacion Produce Tamaulipas A. C. MA and JR acknowledge the support from the Campaña Nacional Contra Moscas de la Fruta.
LITERATURE CITED
Alberti, A. C.; Calcagno, G.; Saidman, B. O and Vilardi, J. C. 1999. Analysis of the genetic structure of a natural population of Anastrepha fraterculus (Diptera: Tephrititdae). Ann. Entomol. Soc. Am. 92:731–736. [ Links ]
Aluja, M. 1993. Manejo integrado de las moscas de la fruta. Ed. Trillas, Mexico D. F. México. [ Links ]
Aluja, M. 1994. Bionomics and management of Anastrepha. Ann. Rev. Entomol. 39:155–178. [ Links ]
Aluja, M. and Mangan, R. 2008. Fruit fly (Diptera: Tephritidae) host status determination: critical conceptual, methodological and regulatory considerations. Annu. Rev. Entomol. 53: In press. [ Links ]
Aluja, M.; Piñero, J.; Jácome, I.; Díaz–Fleischer, F. and Sivinski, J. 2000a. Behavior of flies in the genus Anastrepha (Trypetinae: Toxotrypanini), p. 375–408. In: Aluja, M. and Norrbom, A. L. (eds.), Fruit flies (Tephritidae): phylogeny and evolution of behaviour. CRC Press, Boca Raton, FL. [ Links ]
Aluja, M.; Piñero, J.; López, M.; Ruíz, C.; Zúñiga, A.; Piedra, E.; Díaz–Fleischer, F. and Sivinski. F. 2000b. New host plant and distribution records in Mexico for Anastrepha spp., Toxotrypana curvicauda Gerstacker, Rhagoletis zoqui Bush, Rhagoletis sp., and Hexachaeta sp. (Diptera: Tephritidae). Proc. Entomol. Soc. Wash. 102:802–815. [ Links ]
Aluja, M.; Pérez–Staples, D.; Macías–Ordóñez, R.; Piñero, J.; McPheron, B. and Hernández–Ortiz, V. 2003. Nonhost status of citrus sinensis cultivar Valencia and C. paradisi cultivar ruby red to mexican Anastrepha fraterculus (Diptera: Tephritidae). J. Econ. Entomol. 96:1693–1703. [ Links ]
Armstrong, K. F. and Ball, S. L. 2005. DNA barcodes for biosecurity: invasive species identification. Philos. Trans. R. Soc. Lond. Biol. Sci. 360:1813–1823. [ Links ]
Avise, J. C. 1994. Molecular markers, natural history and evolution. Chapman & Hall, New York, NY. [ Links ]
Baker, A. C.; Stone, W. E.; Plummer, C. C. and McPhail, M. 1944. A review of studies on the mexican fruit fly and related mexican species. USDA, Miscellaneous publication No. 531. [ Links ]
Corsini, G.; Manubens, A.; Lladser, A.; Lobos, S.; Seelenfreund, D. and Lobos, C. 1999. AFLP analisis of fruit fly Ceratitis capitata. Focus 21(3):72–73. [ Links ]
Dowell, R. V.; Sidiqui, I. A.; Meyer, F. and Spaugy, L. 2000. Mediterranean fruit fly preventive release program in Southern California, p. 369–375. In: Tan, K. H. (ed), Area–wide control of fruit flies and other insect pests. Universiti Sains Malaysia, Penang, Malaysia. [ Links ]
Enkerlin, W. 1987. Orientación y dispersión de poblaciones de la mosca mexicana de la fruta (Anastrepha ludens, Loew), estériles y silvestres, en el municipio de Allende, N. L., en el período de septiembre de 1985 a agosto de 1986. M. S. Thesis, Instituto Tecnológico y de Estudios Superiores de Monterrey (ITESM), Campus Monterrey, Monterrey, Nuevo León, México. [ Links ]
Excoffier, L.; Simouse, P. and Quattro, J. 1992. Analysis of molecular variance inferred for metric distances among DNA haplotypes: application to human mitochondrial DNA restriction data. Genetics 131:479–491. [ Links ]
Hernandez–Ortiz, V.; Gomez–Amaya, J. A.; Sanchez, A.; McPheron, B. A. and Aluja, M. 2004. Morphometric analysis of mexican and South American populations of the Anastrepha fraterculus complex (Diptera: tephritidae) and recognition of a distinct mexican morphotype. Bull. Entomol. Res. 94:487–499. [ Links ]
Krafsur, E. S. 2005. Role of population genetics in the sterile insect technique, pp. 389– 406. In: Dyck, V. A., Hendrichs, J. and Robinson, A. S. (eds). Sterile insect technique: principles and practice in area–wide integrated pest management. Springer, Dordrech, The Netherlands. [ Links ]
Mangan, R. L. and Moreno, D. S. 2003. Molecular analysis of Anastrepha ludens genetic variation. http://www.tard.state.tx.us/listing.php?id=3d6d01c202fe8.(05–08–2003). [ Links ]
Nei, M. and Li, W. H. 1979. Mathematical model for studying genetic variation in terms of restriction endonucleases. Proc. Natl. Acad. Sci. USA. 76:5269–5273. [ Links ]
Norrbom, A. L. 2003. Host plant data base for Anastrepha and Toxotrypana (Diptera: Tephritidae: Toxotrypanini). Diptera data dissemination disk. CD–ROM, North Amer. Dipterist's Soc. Washington, D. C. [ Links ]
Powell, W.; Morgante, M.; Andre, C.; Hanafey, M.; Vogel, J.; Tingey, S. and Rafalski, A. 1996. The comparasion of RFLP, RAPD, AFLP and SSR (microsatellite) markers for germplasm analysis. Mol. Breeding 2:225–238. [ Links ]
Reyes, J.; Santiago, G. and Hernández, P. 2000. The mexican fruit fly eradication programme, p. 377–380. In: Tan, K. H. (ed.). Area–wide control of fruit flies and other insect pests. University Sains Malaysia, Penang, Malaysia. [ Links ]
Robacker, D. C. and Fraser, I. 2004. Do mexican fruit flies (Diptera: Tephritidae) prefer grapefruit to yellow chapote, a native host? Fla. Entomol. 85:481–487. [ Links ]
Roderick, G. K. 1996. Geographic structure of insect populations: gene flow, phylogeography, and their uses. Ann. Rev. Entomol. 41:325–352. [ Links ]
S–Plus (S–Plus) User`s Guide Version 4.0. 1997. Seattle: Math Soft. 525 p. [ Links ]
Thomas, D. B. and Loera–Gallardo, J. 1998. Dispersal and longevity of mass–released mexican fruit files (Diptera: Tephritidae). Environ. Entomol. 27:1045–1052. [ Links ]
Thomas, D. B. 2003. Reproductive phenology of the mexican fruit fly Anastrepha ludens (Loew) (Diptera: Tephritidae) in the Sierra Madre Oriental, Northern Mexico. Neotrop. Entomol. 32:385–397. [ Links ]
Thomas, D. B. 2004. Hot peppers as a host for the mexican fruit fly Anastrepha ludens (Diptera: Tephritidae). Fla. Entomol. 87:603–608. [ Links ]
Vera, M. T.; Cáceres, C.; Wornoayporn, V.; Islam, A.; Robinson, A. S.; De La Vega, M. H.; Hendrichs, J. and Cayol, J. P. 2006. Mating in compatibility among populations of the South American fruit fly Anastrepha fraterculus (Diptera: Tephritidae). Ann. Entomol. Soc. Am. 99:387–397. [ Links ]
Via, S. 1990. Ecological genetics and host adaptation in herbivorous insects: The experimental study of evolution in natural and agricultural systems. Ann. Rev. Entomol. 35:421– 446. [ Links ]
Vos, P.; Hogers, R.; Bleeker, M.; Reijans, M.; Van de Lee, T.; Hornes, M.; Frijters, A.; Pot, J.; Peleman, J.; Kuiper, M. & Zabeau, M. 1995. AFLP: a new technique for DNA fingerprinting. Nucleic Acids Res. 23:4407–4414. [ Links ]
West–Eberhard, M. J. 1989. Phenotypic plasticity and the origins of diversity. Annu. Rev. Ecol. Syst. 20:249–278. [ Links ]
Williams, G. C. 1966. Adaptation and natural selection. Princeton University Press, Princeton, USA. [ Links ]
Wright, S. 1951. The genetical structure of populations. Ann. Eugen. 15:323–354. [ Links ]

