










Servicios Personalizados
Revista
Articulo
 Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkAgricultura técnica en México
versión impresa ISSN 0568-2517
Agric. Téc. Méx vol.35 no.3 México jul./sep. 2009
Artículos
Aptitud combinatoria y efectos recíprocos en líneas endogámicas de maíz de Valles Altos del centro de México*
Combinig ability and reciprocal effects in maize inbred lines from the highlands of central Mexico
Miguel Angel Avila Perches1§, Sergio Alfredo Rodríguez Herrera2, Mario Ernesto Vázquez Badillo2, Fernando Borrego Escalante2, Alejandro Javier Lozano del Río2 y Alfonso López Benítez2
1 Campo Experimental Valle de Toluca, INIFAP. Km. 4.5 carretera Toluca–Zitácuaro, vialidad Adolfo López Mateos. 51350. Zinacantepec, México. Tel. y Fax 01 722 27 8 00 31.
2 Departamento de Fitomejoramiento. Universidad Autónoma Agraria Antonio Narro. Buenavista, Saltillo, Coahuila. E–mails: sergio.rodriguez@uaaan.mx, mario.vazquez@uaaan.mx, fernando.borrego@uaaan.mx, alejandro.lozano@uaaan.mx, alfonso.lopez@uaaan.mx.
* Recibido: Junio, 2008
Aceptado: Julio, 2009
§Autor para correspondencia:
avila.miguel@inifap.gob.mx.
RESUMEN
El objetivo de este estudio fue determinar el tipo de acción génica involucrada y la influencia de los efectos recíprocos en el rendimiento y características agronómicas de cruzas dialélicas, directas y reciprocas, de ocho líneas endogámicas de maíz de Valles Altos del centro de México. Se evaluaron ocho progenitores y 56 híbridos simples en dos fechas de siembra, para determinar los efectos de aptitud combinatoria general, específica y efectos recíprocos, se utilizó el método I de Griffing. El estudio se llevó a cabo en Metepec, Estado de México, México en 2005. Los resultados mostraron diferencias entre genotipos y en los efectos de ACG y ACE para la mayoría de los caracteres evaluados. Los efectos genéticos no aditivos fueron de mayor importancia en rendimiento de grano, días a floración masculina y longitud de mazorca, en tanto que los efectos aditivos fueron de mayor relevancia en altura de planta. Los progenitores L1, L2 y L4 presentaron efectos positivos de ACG en rendimiento. El progenitor L4 junto con L3, L6 y L7 mostraron altos promedios en las características agronómicas estudiadas. Entre los híbridos de alto rendimiento, las cruzas 8 X 4, 4 X 1 y 2 X 4 mostraron efectos positivos de ACE; además, el último genotipo mostró la menor altura de planta. Los efectos recíprocos fueron significativos en altura de planta y longitud de mazorca; no obstante, mostraron menores efectos que los de la ACG y ACE.
Palabras clave: Zea mays L., cruzas dialélicas, efectos maternos y no maternos, híbridos simples.
ABSTRACT
The aim of this study was to determine the type of gene action involved and the reciprocal effects upon seed yield and agronomic traits in eight inbred maize lines from the high valleys of central Mexico. Diallel crosses, direct and reciprocals were obtained with the eight lines. Eight parental lines and 56 simple crosses were evaluated in two planting dates in order to determine the general combining ability, specific combining ability and the reciprocal effects using Griffing's method I. The research was carried out during 2005 at Metepec, Estado de Mexico, Mexico. Results show differences among genotypes and in GCA and SCA for most of the evaluated traits. Non–additive genetic effects were important for grain yield, days to anthesis and ear length, whereas the additive effects were significant for plant height. Parents L1, L2 and L4 showed positive effects in GCA for grain yield. Besides L4, parents L3, L6 and L7 showed high values for the agronomic traits considered. Crosses 8 X 4, 4 X 1 and 2 X 4 were among the high yielding hybrids that also showed high positive values of SCA; moreover, the last genotype registered the lowest plant height. The reciprocal effects were significant for plant height and ear length, but of small magnitude than the GCA and SCA parameters.
Key words: Zea mays L. diallel crosses, maternal and non–maternal effects, simple hybrids.
INTRODUCCIÓN
En los Valles Altos del centro de México, situados a más de 2 200 m se siembran 1.5 millones de ha con maíz, lo cual representa 20% de la superficie nacional. De esta superficie, aproximadamente 50% (750 000 ha) se cultivan bajo condiciones de punta de riego y buen temporal en las que se obtienen altos rendimientos; no obstante, el rendimiento promedio regional es menor a cuatro t ha–1; por lo anterior, el uso generalizado de semilla de variedades mejoradas es una alternativa viable para incrementar la rentabilidad del cultivo a corto plazo, por lo que es necesaria la formación de nuevos genotipos superiores a los actuales.
El método de cruzas dialélicas permite estimar la ACG y ACE y se considera eficaz para detectar fuentes de germoplasma útiles para el mejoramiento genético del maíz. El apareamiento de cruzamientos dialélicos es útil para la evaluación de componentes genéticos en la variación del rendimiento de los progenitores y para calcular la capacidad productiva de sus cruzas (Baker, 1978).
La ACG se determina el desempeño promedio de una línea en sus combinaciones híbridas, mientras que, la ACE separa las combinaciones híbridas específicas que resulten mejor o peor de lo que se esperaría en relación con la media de la ACG de las dos líneas progenitoras (Sprague y Tatum, 1942). Con relación al tipo de acción génica que determina la aptitud combinatoria de las líneas, se considera que la ACG determina la porción aditiva de los efectos genéticos, en tanto que la ACE, los efectos no aditivos, esto es, la acción génica de dominancia y epistasis (Poehlman y Allen, 2003). Griffing (1956) propuso cuatro métodos para el análisis dialélico los cuales son de uso más frecuente para estimar los efectos de ACG y ACE, así como para el estudio genético de poblaciones biológicas y la comprensión de la acción génica en caracteres cuantitativos de importancia agrícola; asimismo, suministra al fitomejorador, las herramientas necesarias para la adecuada aplicación de los planes de mejoramiento a emplear (Hallauer y Miranda, 1981). En México, varios fitomejoradores han utilizado algunos de los diseños dialélicos de Griffing, como son los casos de Antuna et al. (2003), De la Cruz et al. (2003) y Reyes et al. (2004) y específicamente en los Valles Altos del centro de México, Pérez et al. (1991), Vasal et al. (1995) y Balderrama et al. (1997) quienes detectaron mayor efecto de la ACG en la manifestación de los caracteres estudiados.
La genética de características ha sido utilizada para el mejoramiento de numerosas especies vegetales, en la mayoría de los estudios se ha resaltado la importancia de los efectos de ACG y ACE e ignoran los factores maternales y recíprocos; es decir, se acepta que existe diferencia en el desempeño de cruzas directas y recíprocas (Mosjidis et al., 1989); sin embargo, en algunas investigaciones se han mostrado valores inconsistentes (en diferentes ambientes) y más pequeños que los obtenidos para ACG o ACE (Velázquez et al., 1992). En algunos casos, el efecto recíproco (ER) ha mostrado ser un factor relevante y de cierta consistencia, por lo que puede tener implicaciones en el mejoramiento genético del maíz (Hansen y Bagget, 1977). La diversidad genética de los progenitores es un aspecto importante que influye en la expresión de los ER en varios caracteres del maíz (Kalsy y Sharma, 1972; Khehra y Bhalla, 1976). Zhang y Kang (1997) y Kang et al. (1999) reportaron que la división del ER en efectos maternos (EMAT) y no maternos (ENMAT) en el análisis dialélico, es útil para determinar si factores maternales o extranucleares están involucrados en la expresión de un carácter. El objetivo de esta investigación fue determinar el tipo de acción génica involucrada, así como la influencia de los efectos recíprocos, en la expresión del rendimiento y algunas características agronómicas de un grupo de líneas endogámicas y sus combinaciones híbridas.
MATERIALES Y MÉTODOS
La investigación se realizó en Metepec, Estado de México, en el Campo Experimental Valle de Toluca del Instituto Nacional de Investigaciones Forestales, Agrícolas y Pecuarias (INIFAP) ubicado a 2 650 m, con clima templado subhúmedo con lluvias en verano, precipitación media anual de 805.4 mm y temperatura media anual de 14.4 °C.
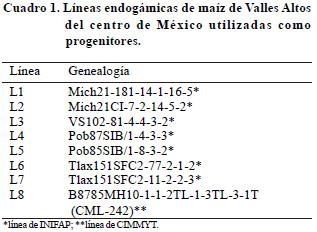
Se incluyeron ocho líneas endogámicas, siete de estas fueron desarrolladas por el INIFAP, con entre tres y cinco generaciones de autofecundación; además, se utilizó una línea del Centro Internacional de Mejoramiento de Maíz y Trigo (CIMMYT) con nivel de endogamia S7. La composición de las poblaciones 85 y 87 (Pob 85 y Pob 87) contienen 60% de germoplasma de Valles Altos, 20% de clima templado y 20% de subtropical–tropical (Eagles y Lothrop, 1994), mientras que el sintético intermedio "B" (SIB) de INIFAP se formó con líneas selectas de Valles Altos (Cuadro 1).
Durante el ciclo primavera–verano de 2004, se realizaron las cruzas dialélicas, directas y recíprocas de las ocho líneas, de las que se generaron 56 cruzas F1; además, se obtuvo semilla de los progenitores por medio de cruzas fraternales. Los 64 genotipos se evaluaron en dos experimentos sembrados el 2 de abril y 6 de mayo de 2005; el primero, se estableció con la aplicación de un riego de presiembra que aportó la humedad necesaria para la germinación y emergencia, el resto del ciclo se mantuvo bajo condiciones de temporal; el segundo, se sembró bajo condiciones de temporal. Cada fecha de siembra (FS) se consideró como un ambiente diferente (AMB) de evaluación. Durante el período de abril a octubre de 2005, la precipitación fue de 820 m, la temperatura media de 14.8 °C y las primeras heladas (–4 °C) ocurrieron el 21 de octubre.
Se utilizó el diseño experimental látice triple 8 X 8, la parcela experimental y útil fue de dos surcos 5 m de largo y 0.80 m de separación. La densidad de población fue de 65 000 plantas por ha–1, el manejo agronómico se realizó con base en las recomendaciones generales del INIFAP para el cultivo de maíz en el valle de Toluca. Se determinaron las siguientes características: días a floración masculina, altura de planta, longitud de mazorca y rendimiento de grano en t ha–1 al 14% de humedad.
En el análisis de la eficiencia del diseño experimental utilizado se observó que fue de 5 a 10% para cada ambiente, por lo que las características agronómicas fueron analizadas como bloques al azar en forma independiente para cada fecha de siembra y se efectuó el análisis combinado. Para la estimación de la ACG, ACE y ER se empleó el análisis dialélico de Griffing (1956) mediante el método I y modelo I, en el que se examinan las líneas parentales, las cruzas F1 directas y recíprocas, para lo cual se utilizó el programa DIALLEL–SAS05 propuesto por Zhang et al. (2005), que permite la división de los ER en maternos (MAT) y no maternos (NMAT).
RESULTADOS Y DISCUSIÓN
El análisis de varianza combinado de los dos experimentos mostró diferencias altamente significativas para los factores ambiente (AMB), genotipo (GEN) y para la interacción GEN X AMB (Cuadro 2). Lo anterior significa que el ambiente (determinado por la fecha de siembra) tuvo efecto sobre los genotipos y las características estudiadas. Lo cual coincide con de la Cruz et al. (2003) en el sentido de que el ambiente modifica las características agronómicas, por lo que es conveniente establecer los ensayos en diferentes ambientales para estimar el valor de los componentes genéticos y separar el efecto del GEN y del AMB.
La cruza 2 X 4 mostró el más alto rendimiento de grano con 11.1 t ha–1, cuatro genotipos (4 X 1, 4 X 2, 1 X 2 y 3 X 1) mostraron un rendimiento superior a 10 t ha–1 (Cuadro 3). En general, los más altos rendimientos se obtuvieron con L4 como progenitor femenino, con más de 9.3 t ha–1 en promedio, resultados similares fueron reportados por Vasal et al. (1995) en el sentido de que la cruza de genotipos de Valles Altos de INIFAP y CIMMYT resulta en alto rendimiento.
Los DFM variaron de 90.5 en la cruza 4 X 3 hasta 97 en la 2 X 5, con media general de 91.7 (Cuadro 3). De las cruzas con alto rendimiento, la más precoz fue 8 X 4 con 91.8 DFM y las más tardías 2 X 1 y 3 X 1 con 97 DFM. La máxima altura de planta se observó en la cruza 7 X 1 con 279.1 cm y la mínima en la 8 X 4 con 204 cm. La altura de la cruza de más alto rendimiento (2 X 4) fue 220.1 cm, considerada baja con respecto a las que mostraron alto rendimiento (Cuadro 3). Resultados similares fueron reportados por Hallauer y Miranda (1981). La mínima longitud de mazorca 10.87 cm se observó en la cruza 5 x 6 y la máxima 14.40 cm en la 2 X 8. Las cruzas 2 X 8 y 4 X 2 mostraron alto rendimiento y longitud de mazorca superior a 14 cm.
Las líneas progenitoras L3, L6, L7 y L1 mostraron rendimiento superior a 3.5 t ha–1, en contraste con L8 con 1.13 t ha–1 (Cuadro 4). Las líneas más precoces fueron L3, L6 y L7 con 96.5 días a floración masculina y las más tardías L1, L2 y L8 con 103.5 días; en general, las líneas fueron mas tardías que sus híbridos, lo cual coincide con de la Cruz et al. (2003) en el sentido de que el desarrollo de los híbridos es más rápido que el de sus progenitores (Cuadro 4). La máxima altura de planta se observó en L1 (243.6 cm) y la mínima (129.8 cm) en L8. La máxima longitud de mazorca (12.73 cm) se observó en L3 y la mínima (7.61 cm) en L2, la mayor correspondió a las líneas que mostraron el más alto rendimiento.
El análisis dialélico combinado de los dos ambientes mostró diferencia significativa en ACG para rendimiento y días a floración masculina y altamente significativa en altura de planta, lo que indica que los progenitores poseen diferente ACG, esto debido principalmente a efectos aditivos. La ACE fue altamente significativa en todas las características evaluadas, lo que indica que al menos una de las cruzas fue diferente a las otras en la ACE a causa de los efectos no aditivos (Cuadro 5).
Los ER fueron significativos en altura de planta y longitud de mazorca, lo que permite inferir que para estas características el valor que se obtiene de la cruza (i, j) es diferente al de la cruza inversa (j, i); Hansen y Bagget (1977), reportaron diferencias recíprocas en altura de planta y longitud de mazorca. Los efectos maternos fueron significativos sólo en rendimiento, en este sentido, Velázquez et al. (1992) observaron EMAT para rendimiento sólo en una de tres localidades (Valles Altos del centro de México). Los efectos maternos fueron significativos y altamente significativos en altura de planta y longitud de mazorca, respectivamente (Cuadro 5). Es importante mencionar, que de acuerdo con Zhang y Kang (1997) el factor materno (recíproco general) se refiere al efecto del genotipo o tejido materno sobre alguna característica de su descendencia, mientras que, Kang et al. (1999) señalan que el componente no materno (recíproco específico) es la interacción entre factores extranucleares y nucleares en las cruzas.
La interacción ACG X AMB fue significativa para altura de planta y altamente significativa para rendimiento y longitud de mazorca. La interacción ACE X AMB fue significativa para días a floración masculina y altamente significativa para rendimiento y altura de planta; lo anterior, resalta la importancia de repetir los experimentos bajo diferentes ambientes para obtener estimaciones consistentes, Vasal et al. (1995). Asimismo, el factor ER X AMB fue significativo en días a floración masculina y altamente significativo en rendimiento. Mientras que EMAT X AMB fue sólo significativo en días a floración masculina y altura de planta; además, ENMAT X AMB fue altamente significativo en rendimiento (Cuadro 5); lo anterior, permite inferir que para estas características, la manifestación del efecto recíproco, materno y no materno dependen del ambiente, lo que coincide con lo reportado por Kang et al. (1999).
Al desglosar las cruzas en ACG y ACE y no obstante de haber mostrado las diferencias significativas mencionadas (Cuadro 5), se determinó que la ACE contribuyó con más de 80% a la suma de cuadrados de genotipos en rendimiento y con más de 58% en días a floración masculina y longitud de mazorca. La importancia de los efectos no aditivos en estas características coincide con de la Cruz et al. (2003); sin embargo, es contrario a lo reportado por Pérez et al. (1991) y podría deberse a que estos últimos autores utilizaron variedades criollas de Valles Aaltos. La ACG contribuyó con 51% en altura de planta, la relevancia de los efectos aditivos en altura de planta también fue reportada por Antuna et al. (2003).
Los ER fueron significativos en altura de planta y longitud de mazorca; en ambos casos, se debió a los efectos no maternos ya que estos fueron significativos en altura de planta y altamente significativos en longitud de mazorca. En general, los ER mostraron valores bajos y aún más bajos que los observados para ACG y ACE los que sólo contribuyeron con 10% en promedio en el desglose de la suma de cuadrados de genotipos del análisis dialélico en las variables evaluadas (Cuadro 5), lo anterior coincide con lo observado por Hansen y Bagget (1977).
La estimación de los efectos de ACG de las líneas endogámicas mostraron que en rendimiento, los valores positivos más altos fueron para la las líneas L4, L1 y L2 con 0.766, 0.443 y 0.378 respectivamente, esto sugiere que estos materiales contienen genes de efectos aditivos que se expresan favorablemente en el rendimiento de grano (Cuadro 6). En contraste, L5, L6, L7 y L8 mostraron valores negativos, lo anterior coincide con Velázquez et al. (1992) que también estimó ACG positivas en líneas derivadas de Mich. 21 y Pérez et al. (1991) quienes reportaron valores altos de ACG en la población Mich. 21 (Cuadro 6). En días a floración masculina, L5 obtuvo el efecto negativo más alto (–1.244 días) y L2 el más alto efecto positivo con 2.244 días. La línea L1 mostró el efecto positivo más alto en altura de planta con 23.567 cm y L4 el más negativo con –16.734 cm. El análisis dialélico combinado no se observó diferencia para ACG en longitud de mazorca (Cuadro 5), por lo que no se presentan estos efectos en el Cuadro 6.
La estimación de los efectos de ACE de las cruzas dialélicas en rendimiento mostraron el más alto efecto positivo en 2 X 8 y 8 X 4 con 7.52 y 7.22 t ha–1, respectivamente (Cuadro 7). Se observó que en siete de las cruzas de más alto rendimiento mostraron efectos positivos superiores a 1.0 t ha–1, lo que coincide parcialmente con lo reportado por Reyes et al. (2004) en el sentido de que una cruza simple será de alto rendimiento si sus dos líneas progenitoras son de alta ACG o si el efecto de ACE es alto y al menos uno de sus progenitores es de alta ACG.
El efecto negativo más alto de ACE para días a floración masculina se observó en la cruza 8 X 4 (–10.30), lo que resulta favorable ya que podría aportar precocidad y rendimiento de grano (9.58 t ha–1); no obstante, también mostró efecto positivo alto en altura de planta (47.35 cm ), lo cual demerita esta característica. Otras cruzas que mostraron efectos positivos en altura de planta fueron 1 X 2 y 4 X 7, (Cuadro 7). Las cruzas 2 X 8 y 8 X 4 mostraron los efectos positivos de ACE más altos para longitud de mazorca con 5.383 y 3.695 cm, respectivamente. Entre los híbridos con alto rendimiento, 2 X 4 mostró efecto positivo de ACE en longitud de mazorca de 1.189 cm.
En la característica altura de planta, el análisis dialélico mostró diferencia significativa en ER, lo cual se debió principalmente a los ENMAT, los híbridos 1 X 4 y 7 X 8 mostraron ER negativos de –14.7 y –22.4 cm, respectivamente (Cuadro 8); asimismo, los mismos genotipos mostraron los mayores ENMAT negativos con –10.5 y –18.0 cm, respectivamente, por lo que utilizados en ese sentido contribuyen al menor porte de planta; por el contrario, si se utilizan las cruzas recíprocas (4 X 1 y 8 X 7), los ER serían los mismos pero positivos y en este caso incrementarían la altura de planta.
En longitud de mazorca el análisis dialélico detectó diferencias en ER, lo cual se debió a la alta significancia de los ENMAT, se observó que los mas altos ER y ENMAT positivos se presentaron en el genotipo 2 X 8 con 1.38 y 0.97 cm, respectivamente; además, la cruza 3 X 5 mostró efectos negativos en estos dos factores con –1.03 y –0.68 cm, respectivamente; es decir, que al utilizar el híbrido 2 X 8 se incrementa la longitud de mazorca; en la cruza recíproca (8 X 2) se obtendría mismo efecto de ER y ENMAT pero negativo y aportaría una reducción en longitud de mazorca (Cuadro 9).
En los híbridos que mostraron valores positivos y negativos significativos de los ER y de los ENMAT en altura de planta y longitud de mazorca y en la mayoría de las cruzas (1 X 4, 7 X 8, 2 X 8, 3 X 5, 4 X 8, 5 X 8 y 1 X 8), intervinieron como progenitores femeninos o masculinos las líneas 4, 5 y 8 las cuales fueron formadas con la integración de germoplasma de Valles Altos, subtrópico y trópico; además, como contrapartes participaron las líneas L1, L2, L3 y L7 de Valles Altos, lo que coincide parcialmente con Kalsy y Sharma (1972) y con Khehra y Bhalla (1976) en el sentido de que la diversidad genética y el diferente origen geográfico de los progenitores influyen en la manifestación de los efectos recíprocos y/o citoplásmicos.
CONCLUSIONES
En las líneas endogámicas de Valles Altos estudiadas, los efectos genéticos no aditivos son relevantes en las características de rendimiento de grano, días a floración masculina y longitud de mazorca en tanto que los efectos aditivos son para altura de planta.
Los progenitores L1, L2 y L4 mostraron los más altos efectos positivos de ACG en rendimiento de grano y sobresalen las cruzas de 8 X 4, 4 X 1 y 2 X 4 con valores altos positivos de ACE.
El híbrido 2 X 4 destaca por las características de rendimiento, altura de planta y longitud de mazorca; asimismo, las líneas endogámicas L3, L6, L7 y L1 mostraron los valores más altos en las características evaluadas.
Los ER observados fueron significativos en altura de planta y longitud de mazorca; sin embargo, presentaron valores menores a los observados en ACG y ACE.
LITERATURA CITADA
Antuna, G. O.; Rincón, S. F.; Gutiérrez del R. E.; Ruiz, T. N. A. y Bustamante, G. L. 2003. Componentes genéticos de caracteres agronómicos y de calidad fisiológica de semillas en líneas de maíz. Rev. Fitotec. Mex. 26:11–17. [ Links ]
Baker, R. J. 1978. Issues in diallel analysis. Crop Sci. 18:533–536. [ Links ]
Balderrama, C. S.; Mejía, C. A.; Castillo, G. F. y Carballo, C. A. 1997. Efectos de aptitud combinatoria en poblaciones de maíz nativas de Valles Altos de México. Rev. Fitotec. Mex. 20:137–147. [ Links ]
de la Cruz, L. E., Gutiérrez del R., E.; Palomo, G. A. y Rodríguez, H. S. 2003. Aptitud combinatoria y heterosis de líneas de maíz en la Comarca Lagunera. Rev. Fitotec. Mex. 26:279–284. [ Links ]
Eagles, H. A. and Lothrop, J. E. 1994. Highland maize from Central Mexico. Its origin, characteristics and use in breeding programs. Crop Sci. 34:11–19. [ Links ]
Griffing, B. 1956. Concept of general and specific combining ability in relation to diallel crossing systems. Aust. J. Biol. Sci. 9:463–493. [ Links ]
Hallauer, A. R. and Miranda, F. J. B. 1981. Quantitative genetics in maize breeding. Iowa State University Press. Ames, Iowa. USA. [ Links ]
Hansen, L. A. and Bagget, J. R. 1977. Reciprocal differences for plant and ear characteristics in sweet corn. Hort. Sci. 12:60–62. [ Links ]
Kalsy, H. S. and Sharma, D. 1972. Study of cytoplasmic effects in reciprocal crosses of divergent varieties of maize (Zea mays L.). Euphytica 21:527–533. [ Links ]
Kang, M. S.; Kushairi, D. A.; Zhang, Y. and Magari, R. 1999. Combining ability for rind puncture resistance in maize. Crop Sci. 39:368–371. [ Links ]
Khehra, A. S. and Bhalla, S. K. 1976. Cytoplasmic effects on quantitative characters in maize (Zea mays L.). Theor. Appl. Genet. 47:271–274. [ Links ]
Mosjidis, J. A.; Waines, J. G.; Yermanos, D. M. and Rosielle, A. A. 1989. Methods for the study of cytoplasmic effects on quantitative traits. Theor. Appl. Genet. 77:195–199. [ Links ]
Pérez, T. R. A.; Carballo, C. A.; Castillo, G. F. y Covarrubias, P. J. 1991. Identificación de patrones heteróticos en un grupo de variedades precoces de maíz. Agrociencia serie Fitociencia 2:69–79. [ Links ]
Poehlman, J. M. y Allen, D. 2003. Mejoramiento genético de las cosechas. Guzmán, O. M.; Hernández, C. M. A. y Serrano, C. L. M. (Trad.). Editorial Limusa. México. [ Links ]
Reyes, L. D.; Molina, G. J. D.; Oropeza, R. M. A. y Moreno, P. E. C. 2004. Cruzas dialélicas entre líneas autofecundadas de maíz derivadas de la raza tuxpeño. Rev. Fitotec. Mex. 27:49–56. [ Links ]
Sprague, G. F. and Tatum, L. A. 1942. General vs specific combining ability in single crosses of corn. J. Am. Soc. Agron. 34:923–932. [ Links ]
Vasal, S. K.; Srinivasan, G.; Vergara, A. N. y González, C. F. 1995. Heterosis y aptitud combinatoria en germoplasma de maíz de Valles Altos. Rev. Fitotec. Mex. 18:123–139. [ Links ]
Velázquez, C. G. A.; Castillo, G. F.; Molina, G. J. D. y Arellano, V. J. L. 1992. Aptitud combinatoria y efectos recíprocos en líneas de maíz (Zea mays L.) con diferente nivel de endogamia. Agrociencia serie Fitociencia 3:111–123. [ Links ]
Zhang, Y. and Kang, M. S. 1997. DIALLEL–SAS: A SAS program for Griffing's diallel analyses. Agron. J. 89:176–182. [ Links ]
Zhang, Y.; Kang, M. S. and Lamkey, K. R. 2005. DIALLEL–SAS05: A comprehensive program for Griffing's and Gardner–Eberhart analyses. Agron. J. 97:1097–1106. [ Links ]

