










Serviços Personalizados
Journal

Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
 Citado por SciELO
Citado por SciELO Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkAgricultura técnica en México
versão impressa ISSN 0568-2517
Agric. Téc. Méx vol.34 no.2 México Abr./Jun. 2008
Artículos
Dinámica de los tizones común y de halo del frijol en el Valle de México*
Dynamics of the bean common and halo blights in the Valley of Mexico
José Manuel Prudencio–Sains1, Rosa Navarrete–Maya1§, Jorge Navarrete–Maya1 y Jorge Alberto Acosta–Gallegos2
1Facultad de Estudios Superiores–Cuautitlán, UNAM, Apartado Postal 25, Cuautitlán Izcalli, Estado de México.
2Programa de frijol, Campo Experimental Bajío, INIFAP.
§ Autor para correspondencia:
rosa_navarrete@hotmail.com
* Recibido: Agosto de 2006
Aceptado: Febrero de 2008
RESUMEN
Los objetivos de este estudio fueron: 1) evaluar la dinámica de los tizones de halo y común del frijol en el Valle de México, bajo condiciones de temporal y, 2) identificar materiales con resistencia a estas enfermedades. Se establecieron once variedades de frijol, de diverso origen y contrastantes en hábito de crecimiento, el 10 de junio de 2001 en Texcoco, Estado de México. La incidencia y la severidad de las enfermedades se evaluaron en diferentes etapas fenológicas y se obtuvieron la tasa de incidencia y el área bajo la curva del progreso de los tizones (ABCPE). Todas las variedades mostraron incidencia de ambas enfermedades, desde el final de la floración la de tizón de halo fue mayor a 5 0% y el común alcanzó una incidencia similar hasta el inicio del llenado de grano. No se observó relación entre la susceptibilidad a las enfermedades y el hábito de crecimiento de las variedades. El rendimiento fue afectado negativamente por el desarrollo de ambas enfermedades, siendo mayor el impacto negativo del tizón de halo. El ABCPE describió con mayor precisión el efecto de las enfermedades sobre el cultivo, que los porcentajes de incidencia y severidad considerados en forma independiente o por fecha de evaluación. Las variedades que mostraron tolerancia a ambas enfermedades fueron Bayo Madero, Negro 8025 y Pinto Villa.
Palabras clave: Phaseolus vulgaris L., Pseudomonas syringae pv. phaseolicola, Xanthomonas axonopodis pv. phaseoli, condiciones ambientales, resistencia.
ABSTRACT
The objectives of this research were 1) to study the dynamics of the diseases bean common and halo blight during the crop cycle at the Valley of Mexico, under rainfed conditions and 2) to identify resistant cultivars to both bacterial diseases. Eleven bean cultivars, from different origin and contrasting growth habit, were sown on June 10, 2001 at Texcoco, State of Mexico. Incidence and disease severity were recorded at different growth stages and the incidence rate and area under disease progress curve (AUDPC) were also estimated. All cultivars showed incidence of both diseases, that of halo blight was higher than 50% from the end of flowering onwards, and common blight reached the same level until the beginning of seed filling. There was not relationship between disease susceptibility and cultivar growth habit. Seed yield was negatively affected by the development of both diseases and halo blight had a larger negative impact. The AUDPC described with major precision the effect of the diseases on the bean crop, in comparison with punctual readings of incidence and severity. Tolerant cultivars against both diseases were Bayo Madero, Negro 8025 and Pinto Villa.
Key words: Phaseolus vulgaris L., Pseudomonas syringae pv. phaseolicola, Xanthomonas axonopodis pv. phaseoli, weather conditions, resistance.
INTRODUCCIÓN
En México el cultivo del frijol (Phaseolus vulgaris L.) con frecuencia es atacado por enfermedades bacterianas como los tizones común y de halo, los cuales tienen una amplia distribución en las zonas productoras de esta leguminosa (Campos, 1991; Navarrete, 1996). El tizón común está considerado entre los cuatro problemas fitopatológicos más importantes de México (López, 1991; Becerra y López, 1994), se ha estimado que las pérdidas que provoca en el rendimiento varían del 15 al 30% (Campos, 1991). Por otra parte, en Michigan, EUA las pérdidas en el rendimiento causadas por el tizón de halo varían del 23 al 43 % (Schwartz, 1989).
El tizón de halo, inducido por Pseudomonas syringae pv. phaseolicola provoca la defoliación prematura de las plantas y puede llegar a causar la muerte de plantas jóvenes de variedades susceptibles sobre todo en los de hábito determinado, tipo I, como Michigan Dark Red Kidney, Canario 107 y Cacahuate 72 (Campos, 1991; Navarrete y Acosta, 2000). En el Valle de México se ha observado que el tizón de halo ataca severamente las variedades de frijoles azufrados y peruanos, de hábito determinado (raza Nueva Granada), y generalmente ésta es la primera enfermedad foliar que se observa en los cultivos de temporal (Navarrete y Acosta, 2000).
El tizón de halo se desarrolla con temperaturas de 16 a 20 °C, humedad relativa alta (> 80%), lluvia y rocío (Zaumeyer y Thomas, 1957; Patel y Walker, 1963; Campos, 1991; Fourie, 2002). P. syringae pv.phaseolicola provoca manchas circulares, con el centro necrosado rodeado por un halo amarillento verdoso. Posteriormente el área necrótica se rasga y cae, si las condiciones son favorables hay defoliación. La enfermedad puede desarrollarse de manera sistémica, los brotes nuevos muestran clorosis y la planta es de menor tamaño que las plantas sanas. En las vainas se forman manchas circulares de aspecto grasoso, con exudados color crema cuando hay humedad relativa alta, al secarse dejan una lesión café rojiza (Rudolph, 1984; Schwartz, 1989). La bacteria puede causar el aborto de semillas, dejarlas enjutas o causar su muerte. Las semillas que alcanzan su desarrollo y están infectadas pueden transmitir a P. syringae pv. phaseolicola, con lo que facilitan su dispersión al siguiente ciclo (Saettler et al., 1991).
Por otra parte, el tizón común inducido por Xanthomonas axonopodis pv. phaseoli generalmente ataca al frijol en la etapa reproductiva en las siembras de temporal. Se han documentado ataques de tizón común en diversas localidades del Altiplano de México, registrándose incidencias de 40 al 100% (Campos, 1991; Jacobo et al., 1992; Navarrete, 1996).
Los síntomas inducidos por X. axonopodis pv.phaseoli son manchas foliares necróticas irregulares, rodeadas por un delgado halo amarillento, que se desarrollan en el borde o en diferentes áreas de la lámina foliar. Las mayores pérdidas ocurren con temperaturas de 27 a 28 °C, humedad relativa alta (>80%) y lluvias frecuentes (Campos, 1991; Saettler et al., 1991; Fourie, 2002). Los síntomas en vainas y semillas son pequeñas manchas rojizas irregulares, con exudados amarillos cuando hay humedad relativa alta (Allen, 1983; Saettler, 1991). Las semillas pueden tener un desarrollo anormal y quedar arrugadas (Saettler, 1991), cuando tienen un buen desarrollo también constituyen un foco de infección, ya que al sembrarse originan plantas enfermas (Campos, 1991; Saettler, 1991).
Para el combate de estas enfermedades se recomienda el control cultural, especialmente el uso de variedades resistentes, el uso de densidades óptimas de siembra, rotación de cultivos por tres años, la eliminación de residuos de cosecha y producción de semilla libre de la enfermedad (Campos, 1991; Saettler, 1991).
Los objetivos de este estudio fueron: 1) evaluar la dinámica de los tizones de halo y común del frijol en condiciones de temporal en el Valle de México, y 2) identificar variedades con resistencia a estas enfermedades.
MATERIALES Y MÉTODOS
Localización. El experimento se estableció en el Campo Experimental Valle de México del Instituto Nacional de Investigaciones Forestales, Agrícolas y Pecuarias (INIFAP), localizado en Texcoco, Estado de México (19° 29' N y 98° 51' O, 2 240 msnm y precipitación media anual de 644 mm), con clima templado húmedo e inviernos benignos (García, 1981).
Germoplasma utilizado y conducción del experimento.
Se utilizaron once variedades de frijol de diferentes orígenes y hábitos de crecimiento (Cuadro 1).
La siembra se realizó el 10 de junio de 2001 en un diseño de bloques completos al azar con seis repeticiones. La parcela experimental fue de un surco de cinco metros con una separación de 0.6 m. Se efectuaron las labores culturales recomendadas para el cultivo en la región; se dio un riego para facilitar la emergencia (16 de junio) y dos de auxilio (13 de julio y 12 de agosto). Se fertilizó al momento de la siembra con la dosis 40–40–00 de N–P–K ha–1.
Evaluación de las enfermedades. A partir de la emergencia de las plantas se realizaron monitoreos para detectar los síntomas de los tizones de halo y común, por infección natural. Se marcó un metro lineal en cada parcela y se evaluaron las plantas dentro de éste, a los 67, 85 y 99 días después de la siembra (dds), considerando los siguientes indicadores:
a) incidencia. Se obtuvo la incidencia de ambos tizones, de halo (P. syringae pv. phaseolicola) y común (X. axonopodis pv. phaseoli) en cada parcela y a través de repeticiones, utilizando la fórmula de Van der Plank (1975):

donde:
I= índice de incidencia (%)
PE= plantas enfermas
PT= plantas totales
b) severidad. La evaluación se basó en la comparación con una escala visual (Schoonhoven y Pastor–Corrales, 1987), donde los valores de 1 a 3 se consideran resistentes, de 3.1 a 6 intermedios y de 6.1 a 9 son susceptibles. Los datos se transformaron a valores semilogarítmicos, se dividieron entre 100 para obtener el porcentaje en centésimas y se obtuvo su raíz cuadrada (Little y Hills, 1989).
c) área bajo la curva del progreso de la enfermedad (ABCPE) y tasa de intensidad de la enfermedad (TIE).
Se determinaron de acuerdo a las fórmulas de Van der Plank (1975):

donde:
Yi= intensidad de la enfermedad (%), obtenida con base en la severidad observada
ti= período de evaluación en días, por lo que las unidades son % día–1
La tasa de intensidad de la enfermedad (TIE) representa la velocidad con que se desarrolla una epidemia y se expresa en % día–1:
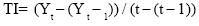
donde:
t y t – 1= fechas en que se cuantificó la enfermedad
Yt y Yt – 1= cantidades de enfermedad registradas en cada fecha.
Condiciones climáticas. Se obtuvieron los datos de precipitación, temperaturas máxima y mínima diarias durante el ciclo de cultivo en una estación meteorológica, localizada a 500 m del sitio experimental.
Análisis estadísticos. Los análisis de varianza se realizaron de acuerdo al diseño experimental con el paquete estadístico MSTATC (Freed et al., 1991) y la separación de medias con la Prueba Múltiple de Duncan al 0.05. Se obtuvieron los coeficientes de correlación simple de Pearson entre los indicadores: severidad, incidencia, área bajo la curva del proceso de la enfermedad, tasa de intensidad de la enfermedad y rendimiento, con los datos obtenidos en cada fecha de evaluación, así como con los datos de máxima severidad y de severidad e incidencia promedio. También se obtuvieron los coeficientes de correlación simple de los datos promedio entre variables climáticas y el desarrollo de las enfermedades.
RESULTADOS Y DISCUSIÓN
Todas las variedades mostraron incidencia de ambas enfermedades, tizón común y de halo. A los 50 dds no se habían detectado síntomas de ninguna de ellas y solamente la variedad Chippata presentaba más del 5 0% de floración. A los 67 dds en todas las variedades se observó una incidencia promedio de tizón de halo mayor al 50%, mientras que el tizón común alcanzó esa incidencia hasta los 85 dds (Cuadro 2). Esto coincide con observaciones anteriores, en que las pudriciones de raíces y el tizón de halo, fueron las primeras enfermedades que se registraron atacando al frijol de temporal en el Valle de México (Acosta y Navarrete, 1996; Navarrete y Acosta, 2000).
Es importante considerar la fecha en que se sembraron las variedades evaluadas en este trabajo, que se considera temprana para la zona, pero se encuentra dentro del lapso recomendado. Es sabido que muchas variedades de frijol son sensibles al fotoperíodo lo que influye en su ciclo biológico acelerando o retrazando la floración (Acosta y White, 1995), tal es el caso de Bayo Madero y Pinto Villa que son sensibles al fotoperíodo, mientras que Negro 8025 es neutral (Rosales et al., 2004). Esta característica aunada a las condiciones climáticas puede explicar porque a los 50 dds aún no había floración en todas las variedades. Cabe mencionar que la presencia del tizón común se asocia con la etapa reproductiva del frijol (Fourie, 2002), de tal forma que si no había floración tampoco se pudo detectar dicha enfermedad en fechas tempranas. Dado que no se realizaron inoculaciones el monitoreo de la incidencia permitió verificar que no hubiera escapes, ya que la incidencia de ambas enfermedades fue alta, la cual puede incrementarse con la precipitacion (Fourie, 2002).
Probablemente las condiciones climáticas promedio (temperatura y precipitación), en el período de los diez días previos a cada evaluación, tiempo suficiente para la incubación de éstos patógenos, fueron favorables para el desarrollo de los tizones (Cuadro 3, Figura 1). En la evaluación a los 67 dds hubo una precipitación acumulada de 15.5mm,temperaturamínimade6.4°C,mediade 15.8°C y máxima de 25.3 °C, condiciones óptimas para el desarrollo del tizón de halo. Posteriormente los valores de temperatura y precipitación fluctuaron, ya que ésta última aumentó mas del doble en la evaluación a los 85 dds y a se redujo a la mitad a los 99 dds (Figura 1, Cuadro 3).
A nivel general, el tizón de halo fue ligeramente mas agresivo que el tizón común a los 85 dds, pero a los 99 dds el tizón común fue mas agresivo que el de halo, como se observa en los promedios de severidad para cada evaluación (Cuadro 4). La severidad máxima del tizón de halo fue de 69.2% en la variedad Chippata. Las variedades mejoradas Bayo Madero, Negro 8025 y Pinto Villa, tuvieron una severidad media de 5.6, 3.7 y 9.1% para el tizón de halo y de 8.4, 11.4 y 13.4% para el tizón común, y se consideran tolerantes a ambas enfermedades. Otras variedades como DON 1013 y Chippata mostraron tolerancia al tizón común, pero fueron intermedias para el de halo. Por el contrario, 97–RS–303 mostró tolerancia al tizón de halo y susceptibilidad al común. Los variedades susceptibles a ambas enfermedades fueron DON 38, Ilomba, HHL 9438–56, BAT 477 y 97–RS–326 (Cuadro 4).
Progreso del tizón de halo
Las variedades que mostraron menor incidencia fueron Bayo Madero y Pinto Villa, ambas de hábito indeterminado tipo III desarrolladas en la región semiárida; mientras que el Pinto 97–RS–326 introducido de EUA, también de hábito indeterminado, mostró 100% de incidencia desde la evaluación realizada a los 67 dds (Cuadro 2).
La severidad inicial de las variedades a los 67 dds varió de 3.4 a 33.2%. A los 85 dds los menores valores de severidad los presentaron las variedades Negro 8025 y Bayo Madero y los mayores Chippata, BAT 477, HHL9438–56 y DON 1013. Por otra parte, en las variedades susceptibles BAT 477, HHL9438–56 y Chippata, la severidad del tizón de halo fue mayor a los 99 dds, en esa etapa varió entre 2.4 a 69.2% y las variedades tolerantes fueron Bayo Madero, Negro 8025 y Pinto Villa, las que mostraron fluctuaciones en la severidad a través del tiempo, a los 99 dds el rango de severidad fue de 1.9 a 3.7% (Cuadro 4). La respuesta de los materiales señalados coincide con trabajos previos que los calificaron como resistentes a moderadamente susceptibles (Beebe y Pastor–Corrales, 1991; Rosales et al., 2004; Beaver et al., 2003).
La variedad Chippata mostró la mayor severidad (Cuadro 4) y tasa de intensidad de la enfermedad (Cuadro 5) a los 85 y 99 dds, la cual fue positiva y ascendente a través del ciclo de cultivo, llegando a cuadruplicar su valor inicial a los 99 dds.
La variedad Pinto Villa confirmó su resistencia al tizón de halo, la que se había observado en un estudio previo efectuado en el mismo sitio, en el que se evaluó el progreso de la enfermedad en frijol sembrado en tres fechas, la última de ellas el 12 de junio de 1997 (Navarrete y Acosta, 2000).
El valor máximo de severidad del tizón de halo fue 69.2%, equivalente a 5.9 en la escala de uno a nueve (Schoonhoven y Pastor–Corrales, 1987), a diferencia del estudio de Navarrete y Acosta (2000), en el que el genotipo Michigan Dark Red Kidney (raza Nueva Granada) alcanzó la máxima severidad 100%, equivalente a nueve con la escala mencionada. Probablemente esto se deba a que las condiciones ambientales fueron diferentes entre los estudios, aún cuando la siembra se realizó en fechas similares, y a que Michigan Dark Red Kidney es más susceptible que cualquiera de las variedades utilizadas en esta investigación.
La tasa de intensidad del tizón de halo en las variedades tolerantes fue disminuyendo conforme avanzó el ciclo de cultivo y en el caso de Bayo Madero fue negativa a partir de los 85 dds (Cuadro 5), es decir la severidad fue menor en esta etapa que al inicio de las evaluaciones, aún cuando el porcentaje de incidencia estuvo en un rango de 52.5 a 78.2% (Cuadro 2). Las variedades Bayo Madero, Ilomba y 97–RS–303 mostraron TIE negativas a partir de los 67 dds; de igual forma se observó para DON 38, DON 1013 y 97 RS–326, lo que significa que un hubo un decremento en la intensidad del tizón de halo, probablemente debido a la combinacion de factores ambientales y a la etapa fenologica avanzada (Cuadro 5).
En relación al hábito de crecimiento de la variedad, la incidencia y la severidad del tizón de halo, las variedades tolerantes son de hábito indeterminado III, mientras que los de mayor severidad incluyeron variedades de los tres hábitos de crecimiento: I, II y III. En esta investgacion sólo hubo una relación directa entre el hábito de crecimiento indeterminado y la resistencia al tizón de halo, a diferencia de lo señalado por Campos (1991) sobre la relación entre la susceptibilidad al tizón de halo y el hábito de crecimiento, ya que no se identificó una relación directa entre ellos. Por otra parte, es probable que esa susceptibilidad si esté asociada al origen de los variedades, las susceptibles de hábito determinado tipo I, por lo general se pueden adscribir a la raza Andina Nueva Granada, las cuales son susceptibles al tizón de halo en ambas etapas vegetativa y reproductiva (Navarrete y Acosta, 2000).
El área bajo la curva del progreso de la enfermedad en las variedades Negro 8025, Bayo Madero y Pinto Villa fluctuó entre 113.9 a 1735.7 % día–1, área equivalente a la novena parte de la registrada por el genotipo más susceptible, Chippata con 1735.7 %día –1 (Cuadro 6).
Las lluvias fueron abundantes durante el período previo a las evaluaciones, lo que permitió que la bacteria se dispersara de manera eficiente hacia las variedades susceptibles vecinas, como se ha documentado sobre el importante papel de la lluvia en la dispersión de esta bacteria (Fourie, 2000). La severidad del tizón de halo y la reacción de los variedades susceptibles estuvo asociada a los cambios ocurridos en las condiciones climáticas (Figura 1). Las condiciones ambientales óptimas para el desarrollo del tizón de halo, que son de >80% de humedad relativa y temperatura media de 19 a 21 °C, se observaron en los 10 días previos al registro de reacción de la enfermedad (Campos, 1991), lo que permitió una adecuada incubación de la bacteria y posteriormente la expresión de síntomas (Cuadro 3).
Por otro lado, no se puede descartar la posibilidad de que exista más de una raza fisiológica de P. syringae pv. phaseolicola en el sitio donde se desarrolló el estudio. Taylor et al. (1996) indicaron que la bacteria presenta a nivel mundial ocho razas fisiológicas; además, se sabe que Bayo Madero es resistente a las razas 1, 2 y 3 (Beebe y Pastor–Corrales, 1991).
Progreso del tizón común
La incidencia del tizón común entre las variedades y las fechas de evaluación fluctuó de 28.0 a 100% y presentó diferencias estadísticamente significativas (p<0.05). La línea 97–RS–326 mostró la mayor incidencia a los 67 dds, de 98.6%, y en las evaluaciones posteriores ésta fue de 100%. La línea 97–RS–303, que como la anterior también fue introducida de Michigan, EUA, alcanzó 100% de incidencia hasta los 99 dds (Cuadro 3). La mayor severidad durante las tres fechas de evaluación la obtuvo la línea 97–RS–326 que alcanzó 75.72% a los 99 dds y la menor 1.9% por la variedad Chippata a los 67 dds (Cuadro 4).
Es importante mencionar que la reacción de resistencia de la variedad Chippata puede ser modificada por la alta susceptibilidad del mismo al tizón de halo (Cuadro 4). Es decir, cuando un genotipo resulta altamente susceptible a la primera enfermedad que ocurre durante el ciclo de cultivo, es difícil determinar con exactitud la reacción de éste a enfermedades subsecuentes. En el caso de P. syringae pv. phaseoli y X. axonopodis pv.phaseoli ambas bacterias se desarrollan a nivel foliar; además como ya se ha mencionado, generalmente el tizón de halo (P. syringae pv.phaseoli) es la primera enfermedad que ocurre y necrosa los tejidos (Navarrete y Acosta, 2000), por lo que al llegar X. axonopodis pv. phaseoli, inductor del tizón común, ya no hay tejido disponible para que éste se desarrolle (Goodwin, 1992).
No se observó ninguna relación entre el hábito de crecimiento, la incidencia y la severidad del tizón común, ya que tanto las variedades con tolerancia como los susceptibles pertenecen a los tres hábitos de crecimiento incluidos en el estudio.
La tasa de intensidad de la enfermedad que mostró la línea 97–RS–326 fue muy elevada desde los 67 dds, casi veinte veces mayor que la variedad Chippata que fue la que mostró la menor severidad y en las siguientes evaluaciones esta tasa también se mantuvo elevada (Cuadro 5). La tasa de intensidad de las variedades DON 38, Bayo Madero e Ilomba mostraron un decremento a partir de los 65 dds.
A los 99 dds en el grupo con los valores más bajos de severidad estuvieron las variedades DON 38, DON 1013, Bayo Madero, Ilomba y Pinto Villa (Cuadro 4). La tasa de intensidad de la enfermedad en DON 38 y Pinto Villa fue disminuyendo a través del ciclo, mientras que en DON 1013, Ilomba y Bayo Madero sufrieron fluctuaciones pero al final su tasa fue menor y negativa (Cuadro 5).
Al realizar los análisis de varianza de la severidad a los 67, 85 y 99 dds se encontraron diferencias significativas entre las variedades evaluadas. Al final de las evaluaciones la mayor severidad la presentó la línea 97–RS–326 (75.72%) a los 99 dds y la menor fue de 1.92% para Chippata a los 67 dds (Cuadro 4).
En el desarrollo del tizón común se observaron fluctuaciones en la severidad; en la mayoría de las variedades ésta se incrementó durante el ciclo de cultivo y alcanzó el máximo porcentaj e a los 99 dds. Sin embargo, en algunas variedades el mayor porcentaje ocurrió a los 85 dds y posteriormente hubo una disminución, tal fue el caso de DON 38, Ilomba y Bayo Madero. Probablemente esto haya ocurrido por los cambios en las condiciones climáticas (temperatura y precipitación) en los días previos a las evaluaciones. Por otra parte, se conoce que esta bacteria presenta variabilidad patogénica (Navarrete y Acosta, 1999), por lo que la respuesta de las variedades puede modificarse debido a esta condición; además de que influya la probable interacción de Xanthomonas axonopodis pv. phaseoli con Pseudomonas syringae pv. phaseolicola.
El ABCPE fluctuó de 299.3 a 2457.6% día–1 el menor correspondió a la variedad DON 1013, lo que representó aproximadamente la octava parte de la mayor ABCPE registrada en la línea 97–RS–326 (Cuadro 6).
Se conoce que el tizón común interactúa negativamente con la sequía, rendimiento y vigor del frijol (Diaz et al., 1991). En este estudio se incluyeron variedades reconocidas como resistentes a patógenos como Pseudomonas syringae pv. phaseolicola, Xanthomonas axonopodis pv. phaseoli, Macrophomina phaseolina, Fusarium spp., Rhizoctonia solani y Colletotrichum lindemuthianum (Cuadro 1). Además de ser resistentes a la sequía, por lo que aquellos que muestran tolerancia al desarrollo del tizón común tienen un amplio potencial para ser utilizados en los programas de mejoramiento de frijol, sobre todo para regiones con baja precipitación como algunas del Altiplano de México, en las cuales el tizón común generalmente se presenta en la etapa reproductiva, favorecida por incrementos en la temperatura (Jacobo et al., 1992; Navarrete, 1996).
Correlaciones
Cuando se analizó la información de cada evaluación, hubo correlaciones positivas y significativas entre la severidad e incidencia del tizón de halo en todas las etapas en las que se determinó la ocurrencia de la misma, a los 67 dds r= 0.659, 85 dds 0.691 y 99 dds 0.689. La correlación entre la severidad del tizón de halo y el común fue positiva y significativa en la evaluación a los 67 dds, posteriormente ya no hubo significancia (Cuadro 7).
A los 67 dds la correlación entre la incidencia y la severidad del tizón común fue altamente significativa (r= 0.855 **), en la segunda evaluación fue significativa y en la tercera evaluación ya no hubo significancia. Por otra parte, en la evaluación a los 67 dds la correlación entre la incidencia del tizón común y la severidad del tizón de halo fue altamente significativa y positiva (r= 0.766 **), mientras que en la evaluación a los 86 dds la correlación fue significativa, pero negativa (r= – 0.766 **). Esto indica que en algunas variedades, como Pinto Villa y Bayo Madero, la incidencia del tizón común sigue incrementandose al tiempo que la severidad del de halo disminuye.
Al realizar correlaciones entre el rendimiento y los demás indicadores evaluados se observó que éste fue principalmente afectado por el tizón de halo. Sin embargo, en el caso del tizón común sólo se observó la influencia de la severidad (r= – 0.562 *) y de la incidencia (r= 0.517 *) sobre el rendimiento en la última evaluación. Esto en concordancia con el hecho de que el tizón común ocurre en la etapa reproductiva. El área bajo la curva del progreso del tizón de halo también mostró una correlación negativa con el rendimiento a los 85 y 99 dds, a diferencia del área de progreso del tizón común en la cual no hubo significancia en ninguna de las evaluaciones (Cuadro 7).
La destrucción del tejido foliar, la defoliación y la senescencia prematura ocasionada por los patógenos necrotróficos puede tener un efecto directo sobre el rendimiento (Gaunt, 1995). Este autor también mencionó que la severidad, número de lesiones o el área de tejido dañado se relacionan con el rendimiento, pero hay mayor relación con el progreso de la enfermedad, aún cuando todos estos indicadores describan a la enfermedad. El ABCPE refleja el efecto del patógeno sobre el crecimiento vegetal ya que muchos patógenos retrazan la emergencia foliar, la extensión foliar, así como el tamaño foliar final, y por lo tanto afectan el rendimiento al haber menor área fotosintética útil. Los tizones de halo y común provocan en el cultivo del frijol efectos como los señalados por Gaunt (1995), por lo que probablemente esta sea la explicación a la correlación negativa entre el rendimiento y la incidencia de tizones observada en esta investigación.
El realizar el análisis por separado del efecto del tizón de halo y del tizón común en cada fecha de evaluación, resultó más revelador que cuando se realiza de manera global, debido a que en el último caso puede enmascararse el efecto individual de cada una de las enfermedades estudiadas. O más aún, cuando se toman en cuenta solamente los valores máximos de severidad de cada enfermedad, independientemente de la fecha en la que ocurran, no se visualizan las diferencias a través del tiempo (Cuadro 8).
El tizón de halo ocurrió al final de la fase vegetativa y el tizón común durante el inicio de la fase reproductiva. La tasa de incremento indicó que el tizón común fue más agresivo ya que desarrolló una severidad mayor que el tizón de halo. Las condiciones climáticas presentes durante el ciclo de cultivo fueron determinantes para el desarrollo de ambas enfermedades (Cuadro 9).
CONCLUSIONES
El tizón de halo incidió significativamente en las variedades susceptibles desde la etapa vegetativa, mientras que el tizón común ocurrió principalmente en la etapa reproductiva.
La interacción entre las variedades susceptibles y la incidencia de estas enfermedades bacterianas, estuvo influenciada por las condiciones ambientales ocurridas durante el período de incubación.
No hubo relación entre el hábito de crecimiento de los variedades y su respuesta al ataque por el tizón común.
El rendimiento fue afectado negativamente por el desarrollo de ambas enfermedades, especialmente por el tizón de halo.
Las variedades de frijol con tolerancia a ambas enfermedades fueron Bayo Madero, Negro 8025 y Pinto Villa, variedades mejorados desarrollados en el Altiplano de México.
LITERATURA CITADA
Acosta–Gallegos, J. A. y Navarrete, M. R. 1996. Identificación de variedades de frijol con resistencia múltiple a enfermedades en la Mesa Central de México. Agric. Téc. Méx. 22(2): 187–197. [ Links ]
Acosta–Gallegos, J. A. and White, J. W. 1995. Phenological plasticity as an adaptation by common bean to rainfed environments. Crop Sci. 35:199–204. [ Links ]
Acosta–Gallegos, J. A. Rosales–Serna, R. Navarrete–Maya, R. y López–Salinas, E. 2000. Desarrollo de variedades mejoradas de frijol para condiciones de riego y temporal en México. Agric. Téc. Méx. 26(1):79–98. [ Links ]
Allen, D. J. 1983. The pathology of tropical food legumes. Disease resistance in crop improvement. J. Wiley & son. Chichester, U. K. 413 pp. [ Links ]
Beaver, J. S. Rosas, J. C. Myers, J., Acosta, J. Kelly, J. D. Nchimbi–Msolla, S. Misangu, R. Temple, S., Arnaud–Santana, E. and Coyne, D. P. 2003. Contributions of the bean/cowpea CRSP to cultivar and germplasm development in common bean. FieldCrops Res. 82:87–102. [ Links ]
Beebe, S. E. and Pastor–Corrales, M. A. 1991. Breeding for disease resistance. In:Van Schoonhiven, A., and Voysest, O. (eds.). Common beans. Research for crop improvement. CIAT, Cali, Colombia. p. 561–617. [ Links ]
Becerra, L. N. y López, S. E. 1994. Enfermedades del cultivo del frijol en el trópico de México. XXI Congreso Nacional de Fitopatología. Soc. Mex. Fitopatol. Cuernavaca, Morelos. p. 56. [ Links ]
Campos, A. J. 1991. Enfermedades de frijol. Editorial Trillas. Primera edición. México. p. 70 – 73. [ Links ]
Díaz, P. R.; Téliz, O. D. y Muñoz, O. A. 1991. Interacción de algunos patógenos del frijol con sequía. Rev. Mex. Fitopatol. 9:121–125. [ Links ]
Freed, R. S. P.; Eisensmith, S.; Goets. D.; Reicosky, V.; Smail, W. and Wolberg, P. 1991. MSTAT–C. A microcomputer program for design, management, and analysis of agronomic research experiments. Michigan State University. [ Links ]
Fourie, D. 2002. Distribution and severity of bacterialdiseases on dry beans (Phaseolus vulgaris L.) in South Africa. J. Pathol.150(4–5):220–226. [ Links ]
García, E. 1981. Modificaciones al sistema de clasificacionesclimáticas de Köeppen. UNAM. México. 252 p. [ Links ]
Gaunt, R. E. 1995. The relationship between plant disease severity and yield. Ann. Rev. Phytopathol. 33:119–144. [ Links ]
Goodwin, P. H. 1992. Effect of common bacterial blight on leaf photosynthesis of bean. Can J. Plant Path. 14:203–206. [ Links ]
Jacobo, J. L.; Ramírez L., M. R. y Fernández, H. P. 1992. Selección y susceptibilidad de germoplasma de frijol al tizón común (Xanthomonas campestris pv. phaseoli (Smith Dye). Rev. Mex. Fitopatol. 11:36–40. [ Links ]
Little, M. T. y Hills, F. J. 1989. Métodos estadísticos para la investigación en la agricultura. Editorial Trillas. Segunda edición. México. 270 p. [ Links ]
López, F. L. C. 1991. Definición de prioridades de investigación fitopatológica para la zona templada del Altiplano Central de México. Agric. Téc. Méx. 17:17–54. [ Links ]
Navarrete, M. R. 1996. Variación patogénica de Xanthomonas campestris pv.phaseoli en México y resistencia del frijol al tizón común. Tesis de Doctorado. Colegio de Postgraduados. Instituto de Fitosanidad. México, p. 1–11. [ Links ]
Navarrete, M. R. y Acosta–Gallegos, J. A. 1999. Pathogenic variability of Xanthomonas campestris pv.phaseoli strains from Mexico. Abstracts 31° Reunión Anual ONTA y 39° Reunión Anúal de la Sociedad Americana de Fitopatología–División Caribe. San Juan, Puerto Rico. p. 84. [ Links ]
Navarrete, M. R. y Acosta–Gallegos, J. A. 2000. Resistencia del frijol al tizón de halo en el Valle de México y progreso de la enfermedad. Rev. Fitotec. Mex. 23:17–28. [ Links ]
Patel, P. N. and Walker, J. C. 1 963. Relation of air temperature and age and nutrition of the host to the development of halo and common bacterial blights of bean. Phytopathology. 53(4):407–411. [ Links ]
Rudolph, K. 1984. Multiplication of Pseudomonas syringae pv. phaseolicola "in planta" I: relation between bacterial concentration and water congestion of different bean cultivars and plant species. Phytopathol. Z. 111:349–352. [ Links ]
Saettler, A. W. 1991. Disease caused by bacteria. In: Hall, R. (ed.). Compendium of bean diseases. APS Press. St Paul, MN. p. 29–30. [ Links ]
Schoonhoven, A. V. y Pastor–Corrales, M. A. 1987. Sistema estándar para la evaluación de germoplasma de frijol. Editado por Centro Internacional de Agricultura Tropical (CIAT). Cali, Colombia. 56 p. [ Links ]
Schwartz, H. F. 1989. Halo blight. In: Bean production problems in the tropics. Schwartz, H. F.,and Pastor–Corrales, M. A. (eds.). CIAT. Cali, Colombia. 654 p. [ Links ]
Taylor, J. D.; Teverson, D. M. and Davis, J. H. C. 1996. Identification and origin of races of Pseudomonas syringae pv. phaseolicola from Africa and other bean growing areas. Plant Pathol. (U. K.) 45(3):469–478. [ Links ]
Van der Plank, J. E. 1975. Principles of plant infection. Academic Press New York. 150 p. [ Links ]
Zaumeyer, W. J. and Thomas, H. R. 1957. A monographic study of bean diseases and methods for their control. Technical Bulletin No. 868. United Stated Department of Agriculture. Washington, D. C. USA. 255 p. [ Links ]

