










Serviços Personalizados
Journal
Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkAgricultura técnica en México
versão impressa ISSN 0568-2517
Agric. Téc. Méx vol.34 no.1 México Jan./Mar. 2008
Artículos
Interacción genotipos X prácticas de manejo en el rendimiento de grano de híbridos de maíz*
Genotypes X crop management interaction grain yield of corn hybrids
Aristeo Barrios–Ayala1 , Antonio Turrent–Fernández2, Rafael Ariza–Flores1, Marco Otero–Sánchez3 y Alejandro Michel–Aceves3
, Antonio Turrent–Fernández2, Rafael Ariza–Flores1, Marco Otero–Sánchez3 y Alejandro Michel–Aceves3
1 Campo Experimental Iguala, INIFAP. km 2.5 carretera Iguala–Tuxpan, Iguala, Guerrero.
2 Área de Productividad, Programa de Edafología, IRENAT, Colegio de Postgraduados.
3 Colegio Superior Agropecuario del estado de Guerrero, Iguala, Guerrero.
Autor para correspondencia:
aristeo_barrios@yahoo.com.mx.
* Recibido: Julio de 2006
Aceptado: Diciembre de 2007
RESUMEN
Una alternativa al método de campo para generar recomendaciones de manejo en México son los modelos modulares dinámicos del crecimiento de los cultivos, que son aditivos en su concepción e ignoran al genotipo. El objetivo de este estudio fue: 1) demostrar que no se pueden generalizar las recomendaciones de manejo del cultivo de maíz para todos los genotipos (de maíz) disponibles para ello y 2) cotejar el efecto de las interacciones genotipo x factores de manejo sobre el rendimiento de grano 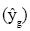 en tres experimentos de campo conducidos bajo riego en el sureste de México, San Gregorio y Villa Flores, Chiapas y Piedras Negras, Veracruz. El diseño experimental fue en parcelas divididas con dos repeticiones, en el que las parcelas grandes se asignaron a 25 tratamientos de un compuesto central rotatorio para N, P, K y densidad de población. Las parcelas chicas fueron asignadas a tres o cuatro híbridos modernos de maíz, todos con desarrollo agronómico similar pero diferente origen genético. El rendimiento de grano se ajustó por regresión a un modelo cuadrático aditivo que ignoró a los genotipos y a otro no aditivo para manejo y genotipo; este último mediante variables mudas con valor 0 ó 1. Los resultados evidencian la interacción genotipo x manejo para el rendimiento de grano, que se manifiesta en diferentes efectos lineales, curvaturas e interacciones entre híbridos, que son cambiantes entre localidades. El modelo no aditivo mejoró entre 30 y 71% el coeficiente de determinación (R2) para rendimiento de grano en relación al modelo aditivo.
en tres experimentos de campo conducidos bajo riego en el sureste de México, San Gregorio y Villa Flores, Chiapas y Piedras Negras, Veracruz. El diseño experimental fue en parcelas divididas con dos repeticiones, en el que las parcelas grandes se asignaron a 25 tratamientos de un compuesto central rotatorio para N, P, K y densidad de población. Las parcelas chicas fueron asignadas a tres o cuatro híbridos modernos de maíz, todos con desarrollo agronómico similar pero diferente origen genético. El rendimiento de grano se ajustó por regresión a un modelo cuadrático aditivo que ignoró a los genotipos y a otro no aditivo para manejo y genotipo; este último mediante variables mudas con valor 0 ó 1. Los resultados evidencian la interacción genotipo x manejo para el rendimiento de grano, que se manifiesta en diferentes efectos lineales, curvaturas e interacciones entre híbridos, que son cambiantes entre localidades. El modelo no aditivo mejoró entre 30 y 71% el coeficiente de determinación (R2) para rendimiento de grano en relación al modelo aditivo.
Palabras clave: Modelos dinámicos, híbridos modernos de maíz; modelo cuadrático no aditivo, modelo cuadrático aditivo.
ABSTRACT
An alternative to the field method for generating recommendations of management in Mexico is the use of modular, dynamic models of crop growth, which are additives in its conception and ignore the genotype. The objetive in this study was: 1) to prove that the recomendations for the use of maize crop cultivation can not be used for all genotypes of maize available for this and 2) data from three maize field experiments conducted in southeastern Mexico, San Gregorio and Villa Flores, Chiapas and Piedras Negras, Veracruz, was analyzed in order to evaluate the genotype x crop management interactions on grain yield . Twenty–five treatments of a Rotable, Central Composite Design for N, P, K and population stand were assigned to main plots of a split plot design replicated twice. Small plots were assigned to three or four modern maize hybrids that had similar agronomic performance but different genetic background. Grain yield was fitted by regression with an additive model that ignored maize genotype and an interactive quadratic model for management factors and maize hybrids, in the last model dummy variables valued 0 or 1 accounted for maize hybrids. Results reflected genotype by management interaction that varied significantly among hybrids and locations in the interactive model. The interactive model consistently improved 30% to 71% the determination coefficient (R2) of grain yield as compared to the additive model.
Key words: Dynamicmodels, modern maize hybrids, quadratic interactive model, quadratic additive model.
INTRODUCCIÓN
Poco se ha estudiado la forma cómo influyen las diferencias genéticas de un cultivo, incluyendo al maíz, en la eficiencia y optimación de diversos factores de manejo. En cuanto al nitrógeno, se menciona que tal capacidad es el producto de numerosos procesos metabólicos controlados genéticamente, que por su complejidad, son difíciles y costosos de medir. Así, Bramel–Cox et al. (1991), encontraron evidencias de la interacción genotipo por nitrógeno entre híbridos comerciales de maíz de los EE.UU. Explican que esta interacción se manifiesta en diferente capacidad genética para absorber y utilizar el nitrógeno. Algunos investigadores suponen que estas adaptaciones controladas genéticamente también existen para los otros insumos y factores tales como: P y K, densidad de población, entre otras. Sin embargo, de estos insumos existen menos estudios (Lafitte y Edmeades, 1991; Zobel, 1991).
La alternativa actual para generar recomendaciones son los modelos dinámicos modulares que en el cultivo de maíz (además de otros parámetros) utilizan el rendimiento de grano para modelar el crecimiento, un ejemplo es el modelo sistémico racional que calcula la fertilización a partir de rendimientos máximos alcanzables y otros parámetros (Etchevers et al., 1991; Johnson, 1991; Rodríguez, 1993). Sin embargo, debido a que estos métodos, al igual que los métodos de modelaje dinámico modular empíricos en uso actual, presuponen la propiedad de aditividad entre los factores suelo, clima, planta y manejo, de la ley de la respuesta de los cultivos de Jenny (1941), las recomendaciones sobre manejo de cultivos, generadas con estos modelos, pueden ignorar al genotipo y tratar como independientes a los factores de manejo, de suelo y clima (Mutsaers y Wang, 1999); lo cual no siempre sucede.
En la presente investigación se evalúan algunos efectos de las interacciones genotipo x manejo, sobre el rendimiento de grano. En estos experimentos se involucra de manera factorial además de los genotipos y factores de manejo, los factores incontrolables asociados a tres localidades. Se plantea la hipótesis de que los híbridos modernos de maíz que alcanzan rendimientos y homogeneidad fenotípica similares, pero que provienen de distinto origen genético, interaccionan significativamente con el manejo y los factores incontrolables de producción, de una región o agrosistema determinado. El objetivo de esta investigación fue demostrar que no se pueden generalizar las recomendaciones de manejo del cultivo de maíz para todos los genotipos (de maíz) disponibles para ello.
MATERIALES Y MÉTODOS
Para el cotejo de la hipótesis de este estudio se aprovecharon tres experimentos que fueron establecidos en el trópico bajo del sureste de México, sembrados bajo condiciones de riego en el ciclo otoño invierno 1997–1998. Las localidades y su clasificación taxonómica del suelo son: San Gregorio, Chiapas. Vertic Ustifluvents; Villaflores, Chiapas. Udic Ustifluvents y Piedras Negras, Veracruz. Udic Ustifluvents, (Soil Survey Staff, 1995). Los híbridos de maíz evaluados fueron: H–512, H–515 y H–516 en Villaflores y Piedras Negras y además de estos tres, el C–343 de Cargill, en San Gregorio, todos similares en rendimiento, altura de planta y ciclo biológico. Para ejemplificar su diversidad genética, se mencionan los programas de mejoramiento que dieron origen a sus líneas progenitoras: del H–512, dos líneas experimentales provienen del trópico húmedo, otra proviene del subtrópico seco (LRB) y la cuarta se obtuvo en el trópico subhúmedo (Iguala, Guerrero.). Las tres líneas progenitoras del híbrido H–515, fueron desarrolladas por el programa de mejoramiento del Instituto Nacional de Investigaciones Forestales, Agrícolas y Pecuarias (INIFAP) para el trópico subhúmedo. Dos líneas progenitoras del H–516, también lo son del H–515, la tercera procede de una población subtropical del programa de mejoramiento de INIFAP. El C–343 proviene de la empresa privada (Noel O. Gómez Montiel, comunicación personal).
El manejo agronómico se realizó con la tecnología que recomienda el (INIFAP, 1988). Durante el ciclo del cultivo, se tomaron datos de altura de planta, días a floración, madurez fisiológica y síntomas de enfermedad que se presentaron previo a la madurez fisiológica. Las parcelas se cosecharon manualmente, tan pronto llegó a su madurez fisiológica, considerando sólo plantas con competencia completa dentro de su surco y a los flancos. Se contaron las mazorcas y se valoró la sanidad, peso, muestra de peso de grano y contenido de humedad de grano, para expresar el rendimiento de grano en t ha–1 con 14% de humedad.
Diseño experimental. Con la fertilización (N, P2O5 y K2O) y la densidad de población (DP) se generaron los tratamientos de parcela grande (TPG) de un diseño de parcelas divididas, seleccionados de un Diseño Compuesto Central Rotable en bloques incompletos e información homogénea (Cochran y Cox, 1957) con dos repeticiones. El espacio de exploración fue: 60>N<220; 0>P2O5<160; 0>K2O<120 y 30>DP<70, en cinco niveles igualmente espaciados y codificados de –2 a +2 de la siguiente manera: n= (N–140)/40, p= (P2O5–80)/40, k= (P2O5–60)/30 y d= (DP–50)/10. El N, P2O5 y K2O se expresan en kg ha-1; la DP en miles de plantas ha-1. Los híbridos de maíz fueron asignados de forma aleatoria a las parcelas subdivididas (chicas). La sub–parcela asignada al híbrido que le correspondió un borde de la parcela grande, constó de dos surcos de ocho m de largo, uno de ellos usado como barrera de protección; el resto (tres o cuatro según el caso) constó de un sólo surco.
Análisis de varianza y de regresión. En cada experimento se ajustaron dos modelos para la variable rendimiento de grano: un cuadrático aditivo sin variables de genotipo y otro cuadrático no aditivo o interactivo con la inclusión de 2 y 3 variables mudas que representan a los 3 ó 4 híbridos de maíz (Allen, 1984). También se realizaron análisis de varianza, regresión y pruebas de falta de ajuste, para el modelo cuadrático no aditivo mencionado. Así mismo, se realizaron selección de variables con (p<0.05) para permanecer en la ecuación.
RESULTADOS Y DISCUSIÓN
Análisis de varianza. El análisis de varianza individual de los tres experimentos, indicó que para factores de manejo e híbridos de maíz, la prueba de F fue significativa a [(p>F)<0.01]. Respecto a la interacción factores de manejo por híbridos de maíz en un experimento la significancia estadística fue de [(p>F)<0.01] (Villaflores) y en dos de [(p>F)<0.05] (San Gregorio y Piedras Negras).
Análisis de regresión. Los modelos generados se presentan en el Cuadro 1. Se incluyen también los coeficientes de determinación (R2) y el cuadrado medio de las desviaciones de regresión (CMDR). La bondad del ajuste de cada uno de los modelos se determinó en función del mayor R2 y menor CMDR. Se observa en el Cuadro 1 que el incremento en la precisión y en la variabilidad explicada al pasar de un modelo cuadrático aditivo que no incluye a los genotipos como variables (CA–SG) a un modelo cuadrático no aditivo que si los incluye (CI–CG), es de al menos 30% como sucedió en San Gregorio, dándose la diferencia más grande en el experimento de Villaflores que fue de 71%, al pasar de un R2 de 0.096 a 0.81 con disminuciones sustantivas en el CMDR. Este incremento en la precisión y sobre todo en la variabilidad explicada, se debió a la variabilidad que define los genotipos y sus interacciones con los factores de manejo. Esta no aditividad mostrada respecto al genotipo remarca la importancia de reconocer que cada híbrido puede manifestar una respuesta estadísticamente diferente en rendimiento de grano a niveles iguales de factores de manejo. Este resultado contrasta con lo propuesto por Volke y Etchevers (1994) quienes sugieren aplicaciones similares de insumos a variedades con potencial de rendimiento similar con el modelo de balance nutrimental (Modelo Sistémico Racional; Rodríguez, 1993).
La inclusión en el modelo de los genotipos y las interacciones con el manejo, consistentemente obtuvo incrementos de R2 y así mismo, al entrar con una presión alta de permanencia en el modelo (p<0.05) además de precisión respecto al modelo aditivo le proporcionan sensibilidad, la cual se reflejará en diferentes óptimos en las dosis de fertilizantes y densidad de población para cada híbrido y por lo tanto en diferentes rendimientos estimados. En este sentido, Mutsaers y Wang (1999) indican que los modelos dinámicos que simulan el crecimiento de los cultivos con la ayuda de equipos de cómputo, son aditivos y no consideran la respuesta individual de los genotipos. Por lo tanto, no explican con eficiencia un distinto potencial de rendimiento en los híbridos y la interacción genotipo x ambiente (Johnson, 1991; Baker, 1996). Un ejemplo de estos modelos es el sistémico racional o de balance nutrimental (Rodríguez, 1993).
Para explicar las manifestaciones de la interacción genotipos x factores de manejo, se tomaron las ecuaciones de regresión ajustadas de dos experimentos, ambos consideraron tres híbridos y cuatro factores de manejo (Piedras Negras y Villaflores). El ajuste se realizó a partir de los rendimientos promedio de grano. Así, el mejor ajuste de los datos generados con el experimento de Piedras Negras incluyó cinco términos que explican 62% de la variabilidad de esta respuesta observada; el CMDR fue de 0.1333 (t ha-1). Los límites de la condición numérica son 1.807 y 33.084 e indican que no se tienen problemas de multicolinealidad; el Fc fue de 27.30, significativa a 0.0001 y prueba la falta de ajuste que no fue significativa. La contraparte, el modelo cuadrático aditivo sin los genotipos como variables (CA–SG), sólo explicó 8.8% de la variación del rendimiento observado y el CMDR fue de 0.305 (t ha-1).
En la ecuación, los términos w1 y w2 corresponden a las variables "mudas o dummy" que para este caso de tres híbridos, toman los valores de: w1= 0 y w2= 0 para el híbrido de referencia, H–516; w1= 1 y w2= 0 para el H–512 y w1= 0 y w2= 1 para el H–515. En este caso, la respuesta de los híbridos H–512 y H–515 será la suma algebraica de los términos sin alguna w, más los términos homólogos que correspondan al efecto de los factores de manejo; esto es, que todos los términos con w son diferencias de respuesta entre híbridos (Barrios et al., 2003; Barrios et al., 2004; Turrent et al., 2005). Por ejemplo, en Piedras Negras la interacción genotipo por densidad de población se detecta porque el H–512 y el H–515 presentan el término d y el H–516 no lo presenta. Lo mismo sucede con la interacción genotipo por N cuadrático (n2) que en esta localidad sólo el H–512 respondió a este término. En Villaflores, cuyo mejor ajuste incluyó siete términos [R2= 0.81, CMDR= 0.4022 (t ha-1), Fc= 50.32 significativa a 0.0001 y límites de la condición numérica de 1.22 y 54.89] sucedió al contrario: el H–515 presenta esta respuesta a n2 y el H–512 y H–516 no; lo mismo sucede con los efectos p2, k2 y d2 del mismo híbrido H–515.
En este experimento, los híbridos H–516 y H–512 son un ejemplo de no–interacción entre híbridos de maíz y factores de manejo. El término –2.667778w1 (significativo al 0.0001) le corresponde al intercepto del H–512 e implica que sólo difiere del H–516 en rendimiento y presenta los mismos efectos de n y d. Estas respuestas son evidencias de la presencia de la interacción genotipos x factores de manejo entre ambos híbridos; las diferencias en la estructura de sus ecuaciones son reflejo de la diferente intensidad con que estos factores afectan a los genotipos, lo que también puede verse como capacidad diferencial para optimizar su uso.
Estos resultados pueden ser una evidencia de que existen diferencias genéticas para la respuesta a estos factores, ya mencionada para N por algunos investigadores (Lafitte y Edmeades, 1991; Zobel, 1991), quienes aluden que los rendimientos potenciales de los híbridos de maíz no sólo son producto de sus requerimientos de manejo, sino también de sus ale los que le confieren tolerancia a factores bióticos (plagas y enfermedades) y a tensiones ambientales, que influyen en la expresión de esta variable. Con el modelo aditivo que ajusta una ecuación genérica para todos los híbridos sería imposible detectar alguna capacidad diferencial que puede existir entre ellos para responder a factores de manejo en función de su composición genética.
En el caso de N, el uso eficiente relacionado con las diferencias genéticas entre los híbridos de maíz se atribuye básicamente a: 1) la capacidad diferencial para acumular N en prefloración, 2) la actividad de enzimas en períodos de llenado de grano y 3) diferencias en estructura y densidad de la raíz (Cretans y Berkum, 1984; Bramel–Cox et al., 1991; Feil et al., 1993).
Es un hecho que los fitomejoradores ahora le confieren a los híbridos componentes genéticos que les den ventaja para enfrentarse a diferentes factores limitantes bióticos (Robinson, 2000) y abióticos (Short, 1991; Jewell et al., 1995). Esto ha propiciado que los híbridos modernos de maíz incluyan en su constitución genética capacidad para tolerar tensiones ambientales y a que no dependan exclusivamente de la "cantidad" de manejo para expresar el potencial de producción. Por lo tanto, el uso del modelo aditivo sólo proporciona información limitada del rendimiento de grano, evidenciando sesgos potenciales en su ecuación y alto grado de generalidad, que se refleja en un R2 bajo, con alto CMDR. Por esta razón considerar al rendimiento de grano como función aditiva de los factores de manejo, puede afectar considerablemente los resultados que se generan con los modelos dinámicos que simulan el crecimiento y la producción de los cultivos (Baker, 1996; Boote et al., 1996; Passioura, 1996). Mutsaers y Wang (1999) analizan en parte esta situación y manifiestan la necesidad de incorporar a estos modelos mayor conocimiento básico que les proporcione la flexibilidad necesaria para adecuarse a situaciones más específicas, no sólo las debidas al componente genético del cultivo, sino también del suelo y clima que aún no estén consideradas.
CONCLUSIONES
La ecuación de regresión ajustada con el modelo no aditivo que incluye a los genotipos y sus interacciones con los factores de manejo mejoró consistentemente la explicación de la variabilidad observada en el rendimiento de grano respecto al modelo aditivo que ignora al genotipo. El ajuste en el rendimiento de grano observado se mejoró entre 30 y 71%.
No se rechaza la hipótesis de que el componente genético de los híbridos de maíz es una parte activa en el diseño de recomendaciones al interactuar con los factores de manejo, por lo que no se debe generalizar una recomendación de manejo para el cultivo de maíz.
LITERATURA CITADA
Allen, E. L. 1984. An introduction to linear regression and correlation. 2ª ed. University of Washington, W. H. Freeman. New York, USA. p. 35–117. [ Links ]
Baker, J. V. 1996. Use and abuse of crop simulation models. Agron. J. 88:681–689. [ Links ]
Barrios A., A.; Turrent F., A.; Cortés F., J. I.; Ortiz S., C. A. y Martínez G., A. 2003. Optimización tecnológica simultánea para tres híbridos de maíz con precisión definida a priori. Terra 21:127–136. [ Links ]
Barrios A., A.; Turrent F., A.; Cortés F., J. I.; Ortiz S., C. A.; Gómez M., N. O. y Martínez G., A. 2004 Interacción genotipos x prácticas de manejo en híbridos de maíz: Efectos sobre el diseño de recomendaciones. Rev. Fitot. Mex. 27(4): 3 99–407. [ Links ]
Boote, K. J.; Jones, J. W. and Pickering, N. B. 1996. Potential uses and limitations of crop models. Agron. J. 88:704–716. [ Links ]
Bramel–Cox, P. J.; Baker, T.; Zavala–Garcia, F. and Eastin, J. D. 1991. Selection and testing environments for improved performance under reduced–input conditions. In: D. A. Sleper et al. (Eds.). Plant breeding and sustainable agriculture: Considerations for objectives and methods. CSSA Special Publ. No. 18. ASA/CSSA/SSSA, Madison, WI. p. 29–56. [ Links ]
Cretans, P. B. and Van Berkum, P. 1984. Genetics of nitrogen metabolism and physiological/biochemical selection for increased grain crop productivity. Theor. Appl. Genet. 67:97–111. [ Links ]
Etchevers B., J. D.; Rodríguez S., J. y Galvis S., A. 1991. Generación de recomendaciones de fertilización, mediante un enfoque sistémico racional. Terra [ Links ]
Feil, B.; Thiraporn, R. and Stamp, P. 1993. In vitro nitrate reductasa activity of laboratory–grown seedlings as an indirect selection criterion for maize. Crop Sci. 33:1280–1286. [ Links ]
Instituto Nacional de Investigaciones Forestales Agrícolas y Pecuarias (INIFAP). 1998. Tecnologías llave en mano. División Agrícola, tomo I , INIFAP. Mexico, D. F. 392 p. [ Links ]
Jenny, H. 1941. Factors of soil formation. McGraw–Hill. New York, USA. p. 450–600. [ Links ]
Jewell, D. C.; Waddington S. R.; Ransom J. K. and Pixley K. V. 1995. Maize research for stress environments. Proceedings of the fourth eastern and southern Africa regional maize conference, held at Harare, Zimbabwe, 28 March – 1 April 1994. Centro Internacional de Mejoramiento de Maíz y Trigo (CIMMYT), México. p. 306. [ Links ]
Johnson, G. W. 1991. General model for predicting crop response to fertilizer. Agron J. 83:367–373. [ Links ]
Lafitte, H. R. and Edmeades, G. O. 1991. Three cycles of recurrent selection in maize for performance under low N conditions. Agron. Abst. 1991:102. [ Links ]
Mutsaers, H., J. W. and Zaoqian Wang. 1999. Are simulation model ready for agricultural research in developing countries? Agron. J. 91:1–4. [ Links ]
Passioura, J. B. 1996. Science, snake oil, education or engineering? Agron. J. 88: 690–694. [ Links ]
Robinson, R. A. 2000. Retorno a la resistencia. Fitomejoramiento para depender menos de los plaguicidas. Trad. F. Romero (2000). Colegio de Postgraduados. Montecillo, México. p. 15–103. [ Links ]
Rodríguez S., J. 1993. La fertilización de los cultivos: un método racional. Facultad de Agronomía, Pontificia Universidad Católica de Chile, Santiago, Chile. [ Links ]
Short, K. E. 1991. Recurrent selection for nitrogen use efficiency in maize. Agron. Abst. 1991:116–116. [ Links ]
Turrent F., A.; Laird J. R.; Cortés F., J. I. y Barrios A., A. 2005. Un reencuentro con la productividad de agrosistemas: II. Validez para adaptar tecnología al maíz en México. Agrociencia 39:149–159. [ Links ]
Volke H., V. y Etchevers B., J. D. 1994. Recomendaciones de fertilización para cultivos: necesidad y perspectivas de una mayor precisión. Cuaderno de Edafología 21. IRENAT, Programa de Edafología, Colegio de Postgraduados, Montecillo, México. p. 43. [ Links ]
Zobel, R. W. 1991. Genetic Control of systems. In: Y. Waisel et al. (Eds.) Plant Roots: The Hidden Half. Marcel Dekker, N.Y. [ Links ]

