










Servicios Personalizados
Revista
Articulo
 Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkAgricultura técnica en México
versión impresa ISSN 0568-2517
Agric. Téc. Méx vol.33 no.3 México sep./dic. 2007
Artículos
EFECTO DE LA ALIMENTACIÓN DE CINCO ESPECIES DE CHINCHES (HEMIPTERA: PENTATOMIDAE, COREIDAE) EN FRUTOS DE NOGAL PECANERO*
FEEDING EFFECT OF FIVE STINK BUG SPECIES (HEMIPTERA: PENTATOMIDAE, COREIDAE) ON PECAN FRUITS
Socorro Héctor Tarango Rivero1 , Mónica Liliana García Bañuelos2 y María del Carmen Candia Plata3
, Mónica Liliana García Bañuelos2 y María del Carmen Candia Plata3
1 Campo Experimental Delicias, INIFAP. Km. 2 carretera Delicias–Rosales, Ciudad Delicias, Chihuahua.
2 Centro de Investigación en Alimentación y Desarrollo, Cuauhtémoc, Chihuahua.
3 Facultad de Ciencias Químico Biológicas, Universidad de Sonora, Hermosillo, Sonora. México.
Autor para correspondencia:
tarango.hector@inifap.gob.mx
* Recibido: Noviembre de 2005
Aceptado: Septiembre de 2007
RESUMEN
El objetivo de esta investigación fue evaluar el efecto de la alimentación de cinco especies de chinches (Hemiptera: Pentatomidae y Coreidae) sobre frutos de nogal pecanero, en distintas fases fenológicas de crecimiento y desarrollo de la nuez, en Delicias, Chihuahua, México. Mediante la técnica de inclusión–exclusión de insectos en campo y con el análisis electroforético del tracto digestivo de las chinches en laboratorio. El experimento se llevó a cabo de 2000–2003 en la región de Delicias. La especie Brochymena sulcata (Van Duzee) no causó ni caída de frutos ni manchado de la almendra; en contraste, Leptoglossus zonatus (Dallas), Leptoglossus clypealis Heidemann y Chlorochroa ligata (Say) sí causaron caída de nueces, cuya intensidad varió con la fase de crecimiento y desarrollo del fruto. Durante el período de llenado de la almendra y apertura del ruezno, L. zonatus y C. ligata causaron manchado significativo de la almendra, mientras que Euschistus servus (Say) sólo lo hizo de manera leve. El análisis del tracto digestivo de B. sulcata mostró la presencia de isoenzimas de masa molecular similar a las del brote del nogal y diferentes a las del ruezno o la almendra. En la digesta de L. zonatus se encontraron isoenzimas similares a las de la almendra, pero no a las del brote. Los estudios demostraron que B. sulcata no es plaga del fruto, mientras que L. zonatus, L. clypealis y C. ligata son plagas potenciales del fruto durante la fase final de crecimiento del mismo, en el llenado de la almendra y hasta la cosecha. E. servus es una especie de importancia menor en las huertas de nogal en Delicias, Chihuahua.
Palabras clave: Brochymena sulcata (Van), Chlorochroa (Say), Euschistus servus (Say), Leptoglossus clypealis (Heidemann), isoenzimas, nuez pecanera.
ABSTRACT
The objective of this study was to evaluate the feeding damage that five different bug species (Hemiptera: Pentatomidae and Coreidae) cause on pecan nuts at different fruit phenological stages. Throughout the technique of including–excluding, stink bugs in pecan shoot terminals were sampled in the field and electrophoretical techniques were used to analyze the content of the digestive track of the bugs in the laboratory. The experiment was carried out from 2000 to 2003 at Delicias, Chihuahua, Mexico. The species Brochymena sulcata (Van Duzee) did not provoke fruit drop or staining on the pecan kernel. On the contrary, Leptoglossus zonatus, L. clypealis, and Chlorochroa ligata caused nut drop whose intensity was related to nut development phase. During the period from nut filling to shuck opening, L. zonatus and C. ligata caused most of the kernel staining damage, while Euschistus servus showed a slight staining effect during this period. The analysis of the digestive track of B. sulcata, showed isoenzymes closely related to the plant growing points and different from these of nut shuck or kernel. While for L. zonatus isozymes closely resembling kernel isoenzymes were found, and different to these from the shoot tissue. Results indicate that B. sulcata does not feed on nuts, whereas L. zonatus L. clypealis and C. ligata are potential fruit pests during the late development stage of the nut, kernel filling and until harvest maturity. E. servus should not be considered an important pecan pest in Delicias, Chihuahua.
Key words: Brochymena sulcata (Van), Chlorochroa (Say), Euschistus servus (Say), Leptoglossus clypealis (Heidemann), isoenzymes, pecan nut.
INTRODUCCIÓN
El nogal pecanero Carya illinoinenensis (Wang.) K. Koch se cultiva en más de 45 000 ha en el estado de Chihuahua (SAGARPA, 2007), donde la principal plaga que ataca a este frutal son los áfidos Monellia caryella (Fitch),Monelliopsis pecanis Bissell y Tinocallis caryaefoliae (Davis) (Homoptera: Aphididae) (Tarango, 2005). En segundo lugar está el barrenador del ruezno Cydia caryana (Fitch) (Lepidoptera: Tortricidae) y el barrenador de la nuez Acrobasis nuxvorella Neunzing (Lepidoptera: Pyralidae) (Tarango et al., 2003a). En los últimos años se ha incrementado la presencia de varias especies de chinches (Hemiptera: Pentatomidae y Coreidae) en las huertas y su daño ha afectado de manera significativa la calidad de la nuez, por lo que son consideradas plagas importantes (Tarango et al., 2003b).
Yates et al. (1991) citan los primeros estudios que describen el daño que causan las chinches a la nuez pecanera. Estos insectos atacan los frutos en la etapa de crecimiento rápido y durante el estado acuoso. Dos días después del ataque la cáscara se torna necrótica y el interior del fruto lo hace a los cuatro o cinco días posteriores, tiempo al cual cae. Después de que la cáscara ha endurecido el daño se presenta en la almendra en desarrollo o completamente llenay se manifiesta en forma de manchas oscuras y amargas. El pico de las chinches apestosas y de patas laminadas tiene la capacidad de penetrar la cáscara endurecida de la nuez, por lo que puede dañar la almendra aun después de que el ruezno ha abierto. La incidencia económicamente importante de las chinches ocurre en la época de llenado de almendra y apertura del ruezno (Smith, 1998).
En las regiones productoras de nuez pecanera de los Estados Unidos de América las especies de chinches que se reportan como plagas importantes son: Euschistus servus (Say), Nezara viridula (L.), Chlorochroa ligata (Say), Acrosternum hilare (Say), Leptoglossus phyllopus (L.) y L. oppositus (Say) (Dutcher y Todd, 1983; Yates et al., 1991; Mizell et al., 1997; Smith, 1998; McPherson y McPherson, 2000; Cottrell et al., 2000). En la región centro–sur del estado de Chihuahua se ha determinado que la chinche gris Brochymena sulcata (Van Duzee) es la más abundante y se encuentra durante todo el ciclo vegetativo, aparece a principios de marzo, su mayor densidad poblacional ocurre desde mediados de mayo hasta finales de junio y permanece en los nogales hastamediados de octubre. La chinche de patas laminadas Leptoglossus zonatus (Dallas) es otra especie común que llega a las huertas en agosto, alcanza un pico poblacional a principios de septiembre y permanece en los árboles hasta mediados de octubre. La conchuela Chlorochroa ligata (Say) se presenta a principios de septiembre, su mayor densidad poblacional se detecta a mediados de octubre y se localiza en las huertas hasta fines de ese mes. La chinche café Euschistus servus (Say), la verde Nezara viridula (L.) y las de patas laminadas L. clypealis Heidemann, L. phyllopus (Linneo) y L. oppositus (Say) se presentan de manera esporádica y en bajo número en las huertas de nogal (Tarango et al., 2003b).
No obstante ser insectos de importancia económica, en las regiones productoras de nuez de Chihuahua es muy poco el conocimiento bioecológico que se tiene de las chinches asociadas al nogal pecanero. En la práctica, cualquier especie de chinche se combate de manera discrecional con plaguicidas, aunque se desconozca el tipo de daño que causa cada una; además, no se cuenta con los métodos de muestreo apropiados ni con los umbrales de acción. Cabe señalar que la implementación exitosa de programas de Manejo Integrado de Plagas (MIP) requiere de información suficiente sobre fenología y potencial productivo de los cultivos, biología y ecología de las plagas y de sus enemigos naturales, relación de las plagas con el cultivo y con los factores biológicos y físicos del ecosistema, potencial de daño económico, métodos de muestreo, umbrales de acción y manejo y aplicación de las diferentes técnicas de control (Henneberry et al., 1991; Gut, 1993). En este trabajo se planteo como objetivo determinar de qué parte del nogal se alimentan las principales especies de chinches que concurren a las nogaleras y la época en que causan daño económico importante. De manera particular se estudió la relación trófica de B. sulcata con el nogal, por ser la especie más abundante en las huertas y sobre la que no hay referencias en la literatura de plagas del nogal.
MATERIALES Y MÉTODOS
Se utilizó la técnica de exclusión e inclusión de insectos con jaulas para los estudios de campo (Dutcher y Todd, 1983; Coombs, 2000). Para los estudios de laboratorio se empleó la técnica de electroforesis, que permite determinar cuál es la fuente de alimento de un amplio número de especies de insectos mediante la detección de isoenzimas en su tracto digestivo (Lucket al., 1999; Amalin et al., 2000).
Estudio de campo. El trabajo se realizó de 2000 a 2002, en la huerta Santa María, en Delicias, Chihuahua. Se utilizaron 10 nogales (repeticiones) de 12 años de edad, vigorosos y con abundante fructificación, de las variedades Western y Wichita. La unidad experimental fue un brote fructífero de 20 a 30 cm de longitud, el cual se cubrió con una jaula de tergalina de 20 cm de diámetro y 60 cm de longitud. Los tratamientos fueron: 1) testigo sin chinches (mediante exclusión), y 2) cinco diferentes especies de chinches (mediante inclusión), cada especie evaluada por separado. En cada jaula se incluyeron dos adultos de cada especie, los cuales permanecieron 10 días en contacto con el brote, hojas y frutos. En cada árbol se colocó una jaula de cada uno de los tratamientos, en un brote seleccionado de la misma longitud y con el mismo número de frutos.
Se evaluó el efecto de la alimentación de las especies B. sulcata, C. ligata, L. zonatus, L. clypealis y E. servus. Durante los años de estudio las chinches se incluyeron en distinto grado de avance de las fases fenológicas de crecimiento y desarrollo, según los criterios de Herrera (1988) y Tarango (1988). En cada experimento se incluyeron las especies fácilmente disponibles en la fecha de su establecimiento. Las chinches de B. sulcata se recolectaron en la misma huerta y en algunos casos en árboles de pinabete (Tamarix sp.) y de huizache (Acacia farnesiana L. Willd) vecinos a las huertas. Los de L. zonatus se obtuvieron de una huerta en Rosales, Chihuahua. Los de C. ligata se recolectaron en mezquites (Prosopis sp.) y en nogales. Los de L. clypealis se obtuvieron de árboles de pistacho (Pistacia vera L.). Los de E. servus se tomaron de campos de maíz (Zea mays L.).
El tiempo de exclusión o inclusión fue de 10 días. A 1, 15 y 30 días después de retirar la jaula se contó el número de frutos retenidos en los tratamientos aplicados en junio, julio y agosto. Para los tratamientos aplicados en septiembre y octubre las nueces se cosecharon en madurez fisiológica, se curaron a la sombra (para fij ar el color del integumento de la almendra) y se midió el número de nueces con manchas de punción por chinches por racimo y el número de manchas por almendra. Los datos se analizaron con un diseño experimental completamente aleatorizado, con el paquete estadístico SAS 6.03 (SAS Institute, 1988).
Estudio de laboratorio. Consistió en analizar y comparar los perfiles de esterasas y peroxidasas del tracto digestivo de las chinches y de tejidos de nogal (brote, ruezno y almendra). El trabajo se realizó en 2002, en la huerta Santa María y en el Laboratorio de Bioquímica del Centro de Investigación en Alimentación y Desarrollo, en Ciudad Cuauhtémoc, Chihuahua.
Insectos. En dos nogales de la variedad Western se cubrieron varios brotes con racimo de 4 o 5 nueces, con jaulas de tergalina. En cada jaula se incluyeron 10 adultos de una misma especie de chinche, durante tres días. Posteriormente se cortaron los brotes con la jaula y se transportaron al laboratorio, donde las chinches permanecieron dos días más. Un grupo control se mantuvo en j aulas de plástico y tapa de tergalina y fue alimentado con una solución de sacarosa al 20%, proporcionada en esferas de algodón de 1.5 cm de diámetro. Las especies evaluadas fueron B. sulcata, L. zonatus y C. ligata.
Material vegetal. En septiembre se recolectaron brotes del año y frutos de nogal de la variedad Western. Una vez cortado, dicho material fue inmediatamente lavado y colocado en hielo a 4 °C para su transporte al laboratorio, donde fue congelado con nitrógeno líquido y almacenado a –80 °C hasta su análisis.
Extracción de proteínas de insectos. El análisis del contenido de esterasas y peroxidasas intestinales de los insectos se efectuó de acuerdo con la metodología utilizada por Amalin et al. (2000). Para ello, el tracto digestivo de las chinches fue disectado íntegramente y la extracción de las proteínas tisulares y las contenidas en la digesta fue realizada mediante la homogenización del tracto digestivo en buffer TBE (Tris–borato 0.09 y EDTA 0.002 M, pH 8.3, con Tritón X–100 al 0.2% y sucrosa al 10%) y una mezcla de inhibidores de proteasas (tripsina y quimiotripsina 0.02 M). Posteriormente, el extracto proteico crudo fue centrifugado a 35 000 g por 20 min a 4 °C y el sobrenadante fue separado y mezclado con un volumen igual de buffer muestra 2X (Tris–sulfato 100 mM, glicerol al 20%, azul de bromofenol al 0.05%, pH 6.5), para realizar el ensayo electroforético.
Extracción de proteínas de tejidos de nogal. Se procesaron muestras de la corteza y las primeras capas de madera del brote, del ruezno y de la almendra, de acuerdo con la metodología reportada por García et al. (2000), que consiste en extraer las proteínas mediante la homogenización de 4 g de tej ido rebanado en 12 ml de buffer de extracción (Trizma base 5 0 mM, ácido cítrico 7 mM, ácido ascórbico al 0.1 %, MgCl2 10 mM, ácido dietilditiocarbámico 10 mM, mercaptoetanol 1 mM y polietilenglicol Mr 3500 al 1%, pH 8) y 0.5 g de PVPP. El homogenizado fue centrifugado durante 20 min a 25 000 g a 4 °C; el sobrenadante se separó y se centrifugó por 12 min en las mismas condiciones. El extracto proteico fue usado inmediatamente para el análisis electroforético o almacenado a –20 °C hasta su utilización.
Electroforesis de isoenzimas y comparación de los patrones electroforéticos. Las proteínas extraídas del tracto digestivo de las chinches y del brote, ruezno y almendra, fueron separadas mediante electroforesis en geles de poliacriamida bajo condiciones nativas (PAGE), usando una minicámara de electroforesis (Bio Rad®) a 4 °C y 60 V por 2.5 h, con un sistema de geles continuos del tipo superior al 5 % de poliacriamida en buffer Tris–sulfato 100 mM, pH 6.5 y geles de separación al 10% o 12% de poliacriamida con buffer Tris–HCl 300 mM, pH 8.3. Como buffer cámara se empleó una solución de Tris–Base 37 mM y glicina 40 mM, pH 8.9 y como buffer cámara inferior Tris–HCl 63 mM, pH 7.5. Las concentraciones de proteína aplicadas a los geles se ajustaron de acuerdo con la actividad y claridad de las bandas. Las muestras fueron aplicadas al gel mezcladas con un volumen igual de buffer muestra (Tris–sulfato 100 mM, glicerol al 20%, azul de bromofenol al 0.05%, pH 6.5). Para estimar la masa molecular de las isoenzimas se usaron estándares de proteínas de 14 200 a 545 000 kDa (Sigma Chemical Co., St. Louis MO.). Al finalizar la corrida electroforética, los geles fueron incubados en la oscuridad y a temperatura ambiente, en soluciones amortiguadoras conteniendo los sustratos enzimáticos alfa y beta naphthyl acetato para esterasas y 3–amino 9 etil carbazol para peroxidasas. La reacción se detuvo mediante la adición a los geles de solución fijadora (ácido acético al 10% y metanol al 25%). Los patrones de isoenzimas detectados fueron visualmente identificados y digitalizados con un escáner HP ScanJet® 3300C.
RESULTADOS Y DISCUSIÓN
Estudio de campo
Caída de frutos
El grado de daño por chinches varía en función de la fase de desarrollo del fruto del nogal que ataque el insecto (Dutcher y Todd 1983; Coombs, 2000). La presencia de B. sulcata al inicio del crecimiento rápido de la nuez y la fase líquida del endospermo (estado acuoso) del 12 al 20 de junio, no tuvo efecto en la caída del fruto, tanto en la variedad Western como en la Wichita (Cuadro 1). En parte, estos resultados son similares a los reportados por Coombs (2000), de que N. viridula no causa caída de frutos Western y Wichita en pospolinización, cuando éstos comienzan su desarrollo.
En un estado más avanzado de desarrollo del fruto se observó que cuando la nuez está en plena fase de crecimiento rápido y del estado acuoso, del 27 de junio al 6 de julio, B. sulcata no causa caída de frutos, pero más notable es que tampoco lo hace la conchuela C. ligata, en las dos variedades de nogal evaluadas (Cuadro 2). Al respecto, se ha demostrado que en esta época N. viridula sí causa caída de nueces (Dutcher y Todd, 1983), pero también que del 12 al 16% de frutos picados por la chinche no caen (Coombs, 2000). Lo anterior sugiere que el fruto del nogal al inicio de su crecimiento rápido no es un alimento de preferencia para las chinches o que éstas tienen otras fuentes de alimentación más accesibles en dicha época del año. En términos de los programas de MIP, esto significa que durante la primera parte del crecimiento de la nuez el muestreo de las chinches no es una práctica necesaria.

Ante la ausencia de efecto en los frutos por la alimentación de C. ligata (Cuadro 2), se repitió el experimento en 2002 en la época de plenitud de crecimiento del fruto y del estado acuoso, del 11 al 22 de julio. Se encontró que B. sulcata no causa caída de nueces, y que tampoco lo hizo L. zonatus. Sin embargo, en comparación con el testigo, en C. ligata se observó 35% de caída de frutos, diferencia altamente significativa (Cuadro 3). Esto sugiere que en la segunda parte del período de crecimiento la nuez aumenta la suceptibilidad al daño por la alimentación de las chinches, época en la cual se justifica efectuar muéstreos y en su caso aplicar medidas de control.

Un cuarto experimento mostró que al final del crecimiento e inicio del estado mucilaginoso de la nuez Wichita, del 22 de julio al 2 de agosto, B. sulcata no causa caída de frutos, en cambio C. ligata lo hizo en 49% y L. clypealis en 64% (Cuadro 4). Este resultado muestra que durante la fase de llenado de la almendra, cuando el fruto tiene altas necesidades nutrimentales (Herrera 1988), la punción por insectos causa el aborto de la nuez. Una respuesta diferencial al daño por chinches fue observada por Coombs (2000), quien observó que durante toda la época de crecimiento de la nuez, la alimentación de N. viridula causó alto porcentaj e de caída en la variedad Western, pero en Wichita se retuvo 90% de los frutos con punción por este insecto en el último tercio de dicha fase fenológica.

Se ha señalado que una nuez dañada por un hemíptero cae a los 4 o 5 días después de la punción (Woodroof y Woodroof citados por Yates et al., 1991); no obstante, en este efecto también existe una amplia variación, ya que como se observa en el Cuadro 4 una nuez Wichita dañada por chinches puede tardar hasta 16 a 3 0 días para caer. Por otro lado, L. clypealis se muestra como una especie con potencial de plaga importante (Cuadro 4), aunque su localización en Chihuahua ha sido en unas cuantas huertas de nogal en centro–sur del estado (Tarango et al., 2003b). Su importancia económica ya ha sido demostrada en huertos de pistachos, donde en dos días de contacto con los frutos puede causar de 23 a 40% de manchado de la almendra y un 33% de caída de frutos (Bolkan et al., 1984).
Los resultados de este estudio y de Dutcher y Todd (1983) y Coombs (2000), se puede inferir que la caída de nuez dañada por un hemíptero depende de varios factores: fase de crecimiento del fruto (y posiblemente de su estado nutricional), especie de chinche y variedad de nogal. También, una observación importante de este trabajo es que B. sulcata no causa caída de nueces, no obstante que es la especie más abundante y permanente en las huertas; además, su población es eficazmente regulada por un conjunto de parasitoides de huevos y de adultos (Tarango et al., 2003b). Las referencias alimenticias de esta chinche indican que se alimenta en el tronco de manzanos y moreras (Ruckes citado por Welter y Purcell, 1987), en el pistachero lo hace de ramas y raquis, pero no del fruto (Welter y Purcell, 1987), y Eger (1981) menciona que esta especie se mantiene en laboratorio con ramas de sauce como alimento.
Manchado de la almendra
En plenitud de la fase fenológica de llenado de almendra de las variedades Western y Wichita, de 10 al 24 de septiembre, B. sulcata no causó manchas en la almendra (Cuadro 5). De manera notoria se encontró que la alimentación de E. servus dio lugar a un manchado muy leve, lo cual en parte puede deberse a que esta especie no tolera el cautiverio, toda vez que al final de la inclusión sólo sobrevivieron 0.4 insectos en promedio por jaula (de dos introducidos); aunque también puede significar que la chinche café es menos dañina para el nogal en la región. En cambio, en 10 días de contacto con la nuez L. zonatus manchó de 75 al 81% los frutos de un racimo, con un promedio de dos a tres manchas por almendra. Un potencial de daño diferente entre especies de chinches sugiere que la determinación de umbrales de acción debería ser específicapara cada unade ellas. El experimento se repitió en 2002 en la variedad Western y se observó que B. sulcata no causó daño, mientras que C. ligata y L. zonatus mancharon más de 80% los frutos de un racimo con más de tres manchas por almendra (Cuadro 6).

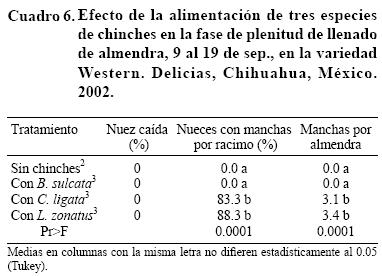
Al final de la etapa de llenado de la almendra e inicio de la apertura del ruezno del 24 de septiembre al 2 de octubre, B. sulcata no mancha la nuez. El número de frutos manchados por C. ligata y por L. zonatus se reduce, pero el número de manchas por almendra es superior a 2.5 (Cuadro 7). Esto sugiere que la almendra llena provee más alimento a las chinches, por lo que requieren menos nueces pero un número similar de punciones a la almendra para su alimentación. Esta observación también sugiere que el umbral de acción no sólo varía entre especies de chinches, sino también con la fase fenológica del fruto.
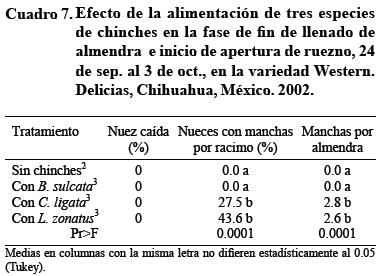
Una vez que la almendra completa el llenado y la mayoría de las nueces ha abierto su ruezno, a mediados de octubre, C. ligata puede manchar 4 nueces d ecada 10 y L. zonatus 8 de cada 10, con más de 3 manchas por almendra en promedio en ambas especies (Cuadro 8). Es decir, la nuez es susceptible de daño por chinche aun sin ruezno que la cubra, cuando está lista para la cosecha. Por su parte, la chinche gris B. sulcata no causa daño en esta época.
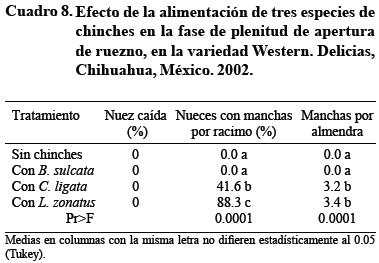
Los últimos cuatro experimentos muestran que la nuez puede ser dañada por punción de hemípteros durante toda la fase de llenado de almendra y de apertura de ruezno. Así, el período crítico de muestreo y protección contra ciertas especies de chinches abarca del inicio de llenado de almendra hasta la recolección de la nuez. Como actualmente no hay plaguicidas convencionales que puedan aplicarse en dichas fases fenológicas del nogal pecanero, por problemas de residuos en la nuez, es necesario el desarrollo de herramientas y/o métodos alternativos de control para estos insectos.
Estudio de laboratorio
Se comparó el patrón electroforético de esterasas de la almendra, ruezno y brote (Figura 1, carriles 2, 3 y 4, respectivamente) con el patrón electroforético de la digesta de L. zonatus y B. sulcata alimentadas de brotes fructíferos del nogal. El patrón de esterasas endógenas del tracto digestivo (Figura 1, carriles 5 y 8) fue obtenido con el análisis de la digesta de chinches alimentadas con sacarosa. En el caso de L. zonatus alimentada de tejidos del nogal (Figura 1, carril 6) se detectaron tres bandas similares a las encontradas en la almendra de la nuez (Figura 1, carril 2), de masa molecular 146, 119 y 87 kDa, mientras que en la digesta de B. sulcata alimentada de tejidos del nogal (Figura 1, carril 7) no se observaron bandas comparables a las del brote, ruezno o almendra.

Por otro lado, la comparación del patrón electroforético de los tejidos de nogal, originado por la detección de peroxidasas, con respecto del patrón obtenido con la digesta de las chinches L. zonatus, C. ligata y B. sulcata (Figura 2), demostró la presencia de una banda en la digesta de C. ligata (Figura 2, carril 6) de masa molecular aproximada a 128 kDa comparable con una de las detectadas en el patrón generado de brotes del nogal (Figura 2, carril 4). De manera similar a lo encontrado en la detección de esterasas, el análisis de la digesta de L. zonatus alimentada de tej idos del nogal (Figura 2, carril 9) mostró dos bandas principales que no aparecen en la digesta de las chinches testigo alimentadas con sacarosa (Figura 2, carril 10) y que son similares a dos bandas del patrón electroforético de la almendra y del ruezno (Figura 2, carriles 2 y 3). En el caso de las isoenzimas extraídas de B. sulcata se observó un patrón similar al generado a partir del análisis del brote del nogal (Figura 2, carril 4), con preponderancia de una banda de masa estimada en 202 kDa (Figura 2, carril 8).

Los resultados anteriores, que coinciden con los observados en campo, muestran que B. sulcata no se alimenta de la nuez ya que no se encontraron isoenzimas del ruezno ni de la almendra en la digesta de esta especie. En contraparte se demostró la presencia de isoenzimas de masa similar a las del brote, lo cual coincide con las referencias de la literatura, que señalan que dicho insecto se alimenta del tallo de diferentes especies arbóreas (Eger, 1981; Welter y Purcell, 1987). En cambio, en la digesta de L. zonatus aparecieron principalmente isoenzimas de esterasas presentes en la almendra, tal como corresponde a una especie que causa fuerte daño a las nueces.
CONCLUSIONES
Durante la época de crecimiento de la nuez el riesgo de daño por chinches es de bajo a moderado. B. sulcata se alimenta de los brotes del nogal y por lo tanto no daña las nueces.
En las fases fenológicas de llenado de almendra y de apertura del ruezno el riesgo de daño por chinches es de moderado a alto. L. zonatus y C. ligata son las especies de hemípteros con más potencial de daño a la nuez en la región. En algunas áreas cercanas a vegetación de matorral L. clypealis puede también ser de importancia económica.
E. servus mostró un potencial de daño más bajo que las especies L. zonatus y C. ligata, para el nogal en la región.
AGRADECIMIENTOS
Al Sr. Alfredo Chávez V. por permitirnos trabajar en su huerta. Por la identificación de las especies de hemípteros al Dr. Harry Braylowsky y a la M. C. Guillermina Ortega L. del Instituto de Biología–Universidad Nacional Autónoma de México (UNAM) y al Dr. Edward Riley del Department of Entomology, Texas A & M University. A Nogales La Trinidad, Campuzano Hermanos, SPR Río Torio, Sr. Carlos Martínez Ch. y Sr. Óscar Rodríguez L. por su apoyo económico para realizar los estudios de electroforesis. Al Sr. Carlos Pérez F. y al Sr. Guerrero Campos G. del Campo Experimental Delicias (CEDEL), Instituto Nacional de Investigaciones Forestales, Agrícolas y Pecuarias (INIFAP), por su excelente trabajo de campo. Al Dr. Carlos Blanco del USDA–ARS por la revisión y sugerencias al manuscrito.
LITERATURA CITADA
Amalin, D. M.; Peña J. E. and McSorley R. 2000. Gut content analysis of three species of sac spiders by electrophoresis. Florida Entomol. 83 (4):489–492. [ Links ]
Bolkan, H. A.; Ogawa J. M., Rice R. E.; Bostock R. M. and Crane J. C. 1984. Leaffooted bug (Hemiptera: Coreidae) and epicarp lesion of pistachio fruits. J. Econ. Entomol. 77:1163–1165. [ Links ]
Coombs, M. T. 2000. Seasonal phenology, parasitism, and evaluation of mowing as a control measure for Nezara viridula (Hemiptera: Pentatomidae) in australian pecans. Environ. Entomol. 29(5): 1027–1033. [ Links ]
Cottrell, T. E.; Yonce C. E. and Wood B. W. 2000. Seasonal occurrence and vertical distribution of Euschistus servus (Say) and Euschistus tristigmus (Say) (Hemiptera: Pentatomidae) in pecan orchards. J. Entomol. Sci. 35 (4):421–431. [ Links ]
Dutcher, J. D. and Todd J. W. 1983. Hemipteran kernel damage of pecan. Misc. Public. Entomol. Soc. Amer. 13:1–11. [ Links ]
Eger, J. E. 1981. Oviposition site preferences and related aspects of the parasitism of Brochymena spp. by Trichopoda plumipes in College Station, Texas. Environ. Entomol. 10:468–471. [ Links ]
García B., M. L.; Tarango R. S. H.; Llamas Ll. J. J. y Gardea B. A. A. 2000. Identificación de portainjertos en pistachero mediante análisis de isoenzimas. En: Memorias. XVIII Congreso Nacional de Fitogenética. Soc. Mex. Fitogenética. p.382. [ Links ]
Gut, L. L. 1993. Integrated pest management overview. In: Orchard pest management. A resource book for the pacific northwest. Good Fruit Grower, p. 17–28. [ Links ]
Herrera, E. 1988. Pecan nut development of 'Ideal' and 'Western Schley' cultivars (Carya illinoensis (Wang.) K. Koch) in the Mesilla Valley, New Mexico. Bulletin 737. AES–CAHE. New Mexico State University. 24 p. [ Links ]
Henneberry, T. J.; Glass E. H.; Gilbert R. G.; King E. G.; Miller R. W. and Whitten C. J. 1991. Integrated pest management, a sustainable technology. In: Agriculture and environment. The 1991 yearbook of agriculture. U.S. Government Print. Off. p. 150–159. [ Links ]
Luck, R. F.; Shepard B. M. and Kenmore P. E. 1999. Evaluation of biological control with experimental methods. In: T.S. Bellows and T.W. Fischer (eds.). Handbook of biological control. Academic Press, p. 225–242. [ Links ]
McPherson, J. E. and McPherson R. M. 2000. Stink bugs of economic importance in America north of Mexico. CRC Press. p.72–162. [ Links ]
Mizell, R. F.; Tedders W. L. and Aldrich J. A. 1997. Stink bug monitoring– an update. Southeast. Pecan Grow. Ass. 90:50–52. [ Links ]
Secretaría de Agricultura, Ganadería, Pesca y Alimentación (SAGARPA). 2007. Avances de siembras y cosechas 2006. Chihuahua. www.sagarpa.gob.mx. [ Links ]
Statistical Analysis Systems Institute (SAS Institute). 1988. SAS/STAT users' guide, release 6.03. SAS Institute, Cary, N.C. [ Links ]
Smith, M. T. 1998. Low input management of stink bugs in pecan: combining the best of conventional control methods and a new trap cropping approach for cost effective IPM. Southeast. Pecan Grow. Ass. 91:52–58. [ Links ]
Tarango R., S. H. 1988. Fenología de algunas variedades de nogal pecanero en Ojinaga, Chihuahua. Informe de investigación. México. Campo Experimental Delicias (CEDEL), Instituto Nacional de Investigaciones Forestales, Agrícolas y Pecuarias (INIFAP). [ Links ]
Tarango R., S. H.; Aguilar P. H. y Quiñones P. F. J. 2003a. Biología, muestreo y control de los barrenadores del ruezno y de la nuez. México. Campo Experimental Delicias (CEDEL), Instituto Nacional de Investigaciones Forestales, Agrícolas y Pecuarias (INIFAP). 26 p. (Folleto técnico No. 12). [ Links ]
Tarango R., S. H.; García M. L. B. y González H. A. 2003b. Especies, daño y control natural de chinches en nogal pecanero. México. Campo Experimental Delicias (CEDEL)–Instituto Nacional de Investigaciones Forestales, Agrícolas y Pecuarias (INIFAP). 39 p. (Folleto técnico No . 14). [ Links ]
Tarango R., S. H. 2005. Control biológico de áfidos del nogal pecanero. México. Campo Experimental Delicias (CEDEL), Instituto Nacional de Investigaciones Forestales, Agrícolas y Pecuarias (INIFAP). 37 p. (Folleto técnico No. 22). [ Links ]
Welter, S. C. and Purcell, M. 1987. A progress report on biological control of hemipteran pests and habitat manipulation in California pistachios. California Pistachio Industry. Ann. Rept. Crop Year 1986–87. p.140–146. [ Links ]
Yates, I. E.; Tedders W. L. and Sparks, D. 1991. Diagnostic evidence of damage on pecan shells by stink bugs and coreid bugs. J. Amer. Soc. Hort. Sci. 116 (1):42–46. [ Links ]

