










Servicios Personalizados
Revista
Articulo
 Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkAgricultura técnica en México
versión impresa ISSN 0568-2517
Agric. Téc. Méx vol.33 no.1 México ene./abr. 2007
Ensayo
MODELO CONCEPTUAL DE COEXISTENCIA ENTRE Phyllophaga crinita y Anomala flavipennis EN AGROECOSISTEMAS DEL NORESTE DE MÉXICO*
A CONCEPTUAL MODEL FOR COEXISTENCE BETWEEN Phyllophaga crinita AND Anomala flavipennis IN AGROECOSYSTEMS OF NORTHEASTERN MEXICO
Luis Angel Rodríguez–del–Bosque1
1 Campo Experimental Río Bravo, INIFAP . Apartado Postal 172, Km 61 carretera Matamoros–Reynosa. 88900 Río Bravo, Tamaulipas, México.
Autor para correspondencia:
rodriguez.luis@inifap.gob.mx
* Recibido: Abril de 2005
Aceptado: Marzo de 2007
RESUMEN
La diversidad de gallina ciega en el norte de Tamaulipas, México se compone de una decena de especies en los géneros Phyllophaga y Anomala. Sólo dos especies, Phyllophaga crinita (Burmeister) y Anomala flavipennis (Burmeister), forman 97% de los adultos capturados en trampas de luz y larvas colectadas del suelo en cultivos de sorgo y maíz. Un estudio de 25 años (1979–2003) en Río Bravo, Tamaulipas, México, informa de una captura alta muy similar de adultos de P. crinita y A. flavipennis en la misma trampa de luz durante la última semana de abril y primera semana de mayo. De ser éste el único dato a considerar, se supondría una gran competencia entre ambas especies en esta región. Si estas dos especies comparten nichos ecológicos (rizofagia) y hábitats (subsuelo) similares, ¿por qué coexisten? El presente ensayo expone los mecanismos y estrategias de P. crinita y A. flavipennis que les permiten coexistir en los agroecosistemas del norte de Tamaulipas. Se discuten las diferencias en la distribución temporal y espacial de P. crinita y A. flavipennis, así como sus atributos poblacionales. Se propone mediante un modelo conceptual, que la coexistencia de ambas especies en esta región se deriva principalmente de la baja probabilidad de competencia entre ellas (4%) en tiempo y espacio por el recurso raíz.
Palabras Clave: Anomala flavipennis, Phyllophaga crinita, competencia interespecífica, ecología, nicho, hábitat.
ABSTRACT
The diversity of white grubs in northern Tamaulipas, Mexico, includes ten species in the genus Phyllophaga and Anomala. However, only two species, Phyllophaga crinita (Burmeister) and Anomala flavipennis Burmeister, comprise 97% of the adults captured in light traps and larvae collected from soil samples in commercial fields of grain sorghum and maize. A study conducted during 25 years (1979–2003) at Río Bravo, Tamaulipas, Mexico, showed similar high captures of P. crinita y A. flavipennis adults in the same light trap during late April and early May. If this was the only available information considered, a strong competition between the two species might be suggested. However, if these two species share similar ecological niches (root feeding) and habitats (underground), why do they coexist?. The present report discusses the mechanisms and strategies of P. crinita y A. flavipennis that allow them to coexist in the agroecosystems of northern Tamaulipas. Differences in their temporal and spatial distribution and population attributes are discussed. A conceptual model is proposed that indicates that the coexistence of both species in this region is mainly due to a low probability for competition (4%) in time and space for food (roots).
Key words: Anomala flavipennis, Phyllophaga crinita, interspecific competition, ecology, niche, habitat.
INTRODUCCIÓN
Las especies con nichos ecológicos similares tienden a competir entre sí hasta provocar el desplazamiento de una de ellas, de acuerdo con el principio ecológico conocido como la Ley de Gauss (1934), el cual establece que dos o más especies que ocupan el mismo nicho ecológico no pueden coexistir indefinidamente en el mismo hábitat. El desplazamiento es el resultado más severo de la competencia interespecífica y es más frecuente entre organismos taxonómicamente relacionados (Reitz y Trumble, 2002).
En el norte de Tamaulipas, se ha practicado el monocultivo de gramíneas (sorgo y maíz) durante las últimas cuatro décadas en cerca de un millón de hectáreas. Entre los insectos dañinos más importantes en esta región, están los que atacan el sistema radicular y la base del tallo, principalmente gallinas ciegas, gusanos de alambre, alfilerillo, trozadores y pequeño barrenador. De éstas, sobresalen las gallinas ciegas por sus altas densidades, daños severos y amplia distribución (Rodríguez–del–Bosque 1980, 1988, 1996a, 1998, Rodríguez–del–Bosque et al., 1995).
La diversidad de gallinas ciegas en el norte de Tamaulipas, se compone de una decena de especies en los géneros Phyllophaga y Anomala. Sin embargo, sólo dos contribuyen con 97% del total de adultos capturados en trampas de luz y de larvas colectadas del suelo en sorgo y maíz, lo que sugiere que el resto de las especies no tienen importancia agrícola. Las dos especies de gallinas ciegas más abundantes en esta región son Phyllophaga crinita (Burmeister) y Anomala flavipennis Burmeister (Coleptera : Scarabaeidae) (Rodríguez–del–Bosque et al., 1995).
Un estudio de 25 años (1979–2003) en Río Bravo, Tamaulipas, indica que la captura total de P. crinita y A. flavipennis adultos en la misma trampa de luz negra (similar a la reportada por Harding et al., 1966) durante la última semana de abril y primera semana de mayo ha sido de 8400 y 8600, respectivamente, lo que representa una proporción muy equilibrada de ambas especies en tiempo y espacio. De ser éste el único dato a considerar, se supondría una gran competencia entre ambas especies en la región. Si éstas comparten nichos ecológicos (rizofagia) y habitats (subsuelo) similares, ¿por qué coexisten y no aplica la Ley de Gauss en este caso? La presente investigación tuvo como objetivo explicar mediante un modelo conceptual los mecanismos y estrategias de P. crinita y A. flavipennis que les permiten coexistir en los agroecosistemas del norte de Tamaulipas.
CICLO BIOLÓGICO
Las dos especies, P. crinita y A. flavipennis, difieren notablemente en sus respectivos ciclos de vida en el noreste de México. A su vez, los mismos ciclos difieren para las mismas especies en otras áreas de distribución geográfica. En esta región, P. crinita tiene un ciclo biológico anual. Los adultos emergen con mayor abundancia desde finales de abril hasta finales de mayo (Figura 1). La presencia de huevos en el suelo se observa comúnmente durante mayo. El primer y segundo estadios larvales se desarrollan normalmente durante junio y julio, respectivamente. El tercer estadio se observa comúnmente a partir de agosto hasta fines de marzo y principios de abril, cuando comienzan a pupar (Rodríguez–del–Bosque 1982, 1988, 1996b). En latitudes mayores P. crinita puede tener un ciclo de vida cada dos años (Teetes et al., 1976).

Por su parte, A. flavipennis tiene un ciclo de vida bivoltino en esta región, con una generación que emerge en primavera y la segunda en otoño. Los adultos de la primera generación emergen principalmente desde fines de abril hasta mediados de mayo y la segunda desde finales de agosto hasta finales de septiembre. Las larvas de la primera generación ocurren en junio–julio y las de la segunda generación desde octubre hasta principios de marzo. Las pupas ocurren comúnmente en marzo y agosto en la primera y segunda generación, respectivamente. En contraste, A. flavipennis se comporta como una especie univoltina en latitudes mayores, incluyendo los estados de Nebraska, Kansas y Georgia, EE. UU., en donde los adultos emergen solamente en junio–julio (Hayes and McColloch, 1924; Forschler y Gardner, 1991; Ratcliffe, 1991). La adaptación al bivoltinismo en A. flavipennis al sur de su distribución, en comparación con el univoltismo en el norte, se atribuye a la acumulación del doble de unidades calor a través del año, lo que le permite completar dos generaciones por año (Rodríguez–del–Bosque, 1998).
ABUNDANCIA, ECOLOGÍA Y HÁBITOS
Es notoria la mayor abundancia de P. crinita y A. flavipennis en el norte de Tamaulipas, en comparación con otras localidades en sus respectivos rangos de distribución. En esta región, se han capturado hasta 47 000 y 129 000 adultos de A. flavipennis durante toda la época de emergencia en la primera y segunda generación, respectivamente, muy por encima de las capturas en otras localidades de EE. UU. (Rodríguez–del–Bosque, 1998). La captura de adultos de P. crinita ha alcanzado hasta 45000 en un año, cifras también superiores a las reportadas en otras localidades de México y EE. UU. (Rodríguez–del–Bosque 1988, 1993 y Rodríguez del Bosque et al., 1995). Los registros diarios en la trampa de luz ubicada en Río Bravo, Tamaulipas, indican una captura récord de 12 000 adultos de P. crinita el 8 de mayo de 1992 y de 25 000 adultos de A. flavipennis el 20 de agosto de 1992 (Rodríguez–del–Bosque, 1993).
Una de las probables causas de las altas poblaciones de P. crinita en esta región es la práctica del monocultivo de gramíneas (sorgo y maíz) en casi un millón de hectáreas durante las últimas cuatro décadas. Lo anterior se basa en que P. crinita prefiere ovipositar en sorgo y maíz que en otros cultivos como girasol, soya y frijol (Rodríguez–del–Bosque, 1984). Aunque no se ha determinado la preferencia de A. flavipennis a diversas plantas hospederas, es probable que esta especie no sea tan atraída a las gramíneas como P. crinita, debido a la limitada presencia de larvas observada (menos del 20%) en muestreos intensivos en sorgo y maíz, en donde P. crinita domina con más del 80% de las larvas colectadas (Rodríguez–del–Bosque et al. 1995). Aún se desconoce cuáles son las plantas hospederas o hábitats preferidos por A. flavipennis, cuya abundancia en esta región es explicada más por su bivoltinismo que por el monocultivo de gramíneas.
Invariablemente, la precipitación o el agua de riego estimulan la emergencia de adultos de P. crinita entre tres y cuatro días después del riego o una precipitación significativa (Gaylor y Frankie, 1979; Rodríguez–del–Bosque, 1993). Esta condición le permite a las hembras ovipositar en suelos húmedos pero no saturados para garantizar una mayor sobrevivencia de huevos y larvas (Gaylor y Frankie, 1979). En contraste, el efecto de la lluvia o riego no e s determinante en A. flavipennis a pesar de que, en la mayoría de los casos, la precipitación estimula la emergencia de A. flavipennis en promedio seis días después, es frecuente observar gran actividad de adultos aún bajo condiciones de sequía. El impacto de las condiciones de suelo seco sobre la oviposición y sobrevivencia de A. flavipennis se desconocen (Rodríguez–del–Bosque, 1998).
La sobrevivencia de larvas y pupas de A. flavipennis es significativamente mayor que la de P. crinita bajo condiciones controladas. En contraste, la longevidad de adultos de P. crinita es mayor que la de A. flavipennis. Se localizó que una hembra de P. crinita vivió 87 días bajo condiciones extremadamente secas, lo que demuestra la capacidad de esta especie de sobrevivir en la celda pupal por largos períodos, en espera de la ocurrencia de lluvia para emerger y así encontrar condiciones propicias para ovipositar (Rodríguez–del–Bosque, 1996b).
ACTIVIDAD ALIMENTICIA
La actividad alimenticia estacional de P. crinita es diferente a la de A. flavipennis como resultado de sus diferentes ciclos biológicos (voltinismo) en el norte de Tamaulipas (Figura 2). Con base en el porcentaje de daño a la raíz, la actividad alimenticia de las larvas de segundo estadio de P. crinita es moderada (41%) durante julio. El tercer estadio tiene la máxima actividad desde julio a septiembre (66–88%), para después disminuir gradualmente de octubre (33%) a diciembre (5%) y cesar su alimentación en enero y febrero (Rodríguez–del–Bosque, 1996a). La mayoría de las especies de gallinas ciegas cesan su alimentación y emigran a mayores profundidades del suelo, particularmente en zonas templadas (Ritcher, 1958). Sin embargo, esto no ocurre en los trópicos, como el caso de Ligyrus subtropicus Blatchely, la cual se alimenta de las raíces de la caña de azúcar durante todo el año en Florida, EE. UU. (Cherry, 1991).

En contraste, las larvas de primera generación de A. flavipennis tienen una mayor actividad alimenticia durante julio (72%), la cual decrece rápidamente en agosto (14%), para dar inicio al estado pupal. Las larvas de la segunda generación se alimentan activamente en noviembre (53%), para decrecer gradualmente de diciembre (16%) a enero (6%). Las larvas permanecen inactivas en febrero y marzo, para después iniciar el período pupal (Rodríguez–del–Bosque, 1996a).
UN MODELO CONCEPTUAL DE COEXISTENCIA
Ambas especies de gallinas ciegas, P. crinita y A. flavipennis, tienen en común el mismo nicho ecológico: la rizofagia. La gran capacidad de estas dos especies de dañar el sistema radicular ha sido demostrada en esta región (Rodríguez–del–Bosque, 1996a). Además, son muy abundantes en esta localidad. Estas dos características per se sugerirían una gran competencia entre las dos especies por el recurso en esta zona. Sin embargo, existen otras características biológicas y ecológicas que las diferencian y que, evidentemente, les permiten la coexistencia en esta región. Dichas diferencias han sido detalladas en las secciones anteriores.
Con base en la información generada durante 25 años sobre la biología y ecología de P. crinita y A. flavipennis en el norte de Tamaulipas, se propuso un modelo conceptual para explicar su coexistencia mediante el análisis de dos características diferentes entre ambas especies: a) la actividad alimenticia estacional (tiempo) y b) el hábitat (espacio). Si ambas especies coinciden en la utilización del recurso raíz (alimento) en el mismo tiempo (x) y espacio (y) estarían bajo una presión de competencia determinada en función de la magnitud de dicha coincidencia (xy).
Tomando como base la Figura 2, donde se muestra la probabilidad de actividad alimenticia de larvas de P. crinita y A. flavipennis (Rodríguez–del–Bosque, 1996a), a continuación se discuten tres escenarios conceptuales de competencia entre ambas especies:
a) Competencia máxima. Supongamos dos dimensiones en el eje de las "yâ para indicar dos posibles hábitats: 1) el agroecosistema de sorgo–maíz, que es el más común en el norte de Tamaulipas; y 2) otros cultivos diferentes a sorgo–maíz u otras plantas hospederas en áreas sin sembrar, como en canales, drenes, caminos, o en pequeñas áreas silvestres, cerca de las parcelas comerciales. Si ambas especies de gallina ciega tuvieran la misma oportunidad de aprovechar ambos hábitats, la competencia entre ellas por el alimento tendría una probabilidad máxima, representado por la Figura 3. Sin embargo, aunque ambas especies se traslapan mutuamente en el 100% en los hábitats de sorgo–maíz y otras plantas hospederas (y), la competencia (coincidencia) entre ellas es de sólo 28%, debido a su diferente ocurrencia temporal (x). Este valor de competencia fue calculado con base en el área de traslape de las dos especies de gallinas ciegas (PA) dividido entre la suma de las áreas de ambas especies (P+A) multiplicado por 100:
Área de traslape de ambas especies (PA)= 53
Área de P. crinita (P)= 100 Área de A. flavipennis (A)= 88
% Competencia entre ambas especies (PA)= (53/188) x 100= 28
Las áreas fueron calculadas de acuerdo a los datos presentados en la Figura 2 y estandarizadas con base a 100 para el área de P. crinita.

b) Competencia nula. Supongamos que ambas especies son mutuamente excluyentes al preferir diferentes hábitats: P. crinita a sorgo–maíz y A. flavipennis a otras plantas hospederas. Bajo este escenario (Figura. 4), la competencia entre ambas especies de gallina ciega sería nula, debido a que no existe traslape alguno entre ellas en los dos hábitats:
Área de traslape de ambas especies (PA)= 0
Área de P. crinita (P)= 100
Área de A. flavipennis (A)= 88
% Competencia entre ambas especies (PA)= (0/188) x 100= 0

c) Competencia mínima. Supongamos que ambas especies se traslapan mutuamente en 20% en los hábitats: sorgo–maíz y otras plantas hospederas de acuerdo al reporte de Rodríguez–del–Bosque et al., (1995), discutido previamente. En este escenario (Figura 5), la competencia entre ambas especies de gallina ciega sería de tan sólo 4%, debido al mínimo traslape entre ellas en los dos hábitats (20%) y a su diferente ocurrencia temporal:
Área de traslape de ambas especies (PA)= 7
Área de P. crinita (P)= 100
Área de A. flavipennis (A)= 88
% Competencia entre ambas especies (PA)= (7/188) x 100= 4
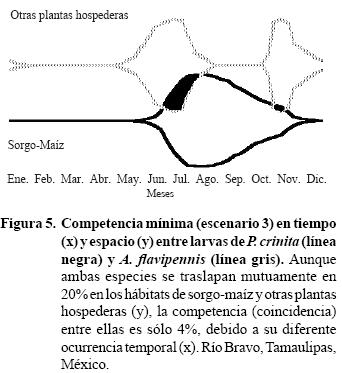
Se propone este escenario de competencia mínima como el responsable de la coexistencia de P. crinita y A. flavipennis en el norte de Tamaulipas. Este mecanismo de mínimo traslape en tiempo y espacio, aunado al subsidio de energía característico de los grandes agroecosistemas, ha permitido a las dos especies coexistir en forma abundante en esta región, comparado con las menores poblaciones observadas en otras áreas de su respectiva amplia distribución geográfica.
LITERATURA CITADA
Cherry, R. H. 1991. Feeding rates of different larval instars of a sugarcane grub, Ligyrus subtropicus Blatchely (Coleoptera: Scarabaeidae). J. Agric. Entomol. 8:163–168. [ Links ]
Forschler, B. T. and Gardner, W. A. 1991. Flight activity and relative abundance of phytophagous Scarabaeidae attracted to blacklight traps in Georgia. J. Agric. Entomol. 8:179–187. [ Links ]
Gauss, G. F. 1934. The struggle for existence. Williams and Williams. Baltimore, USA. [ Links ]
Gaylor, M. J. and Frankie, G. W. 1979. The relationship of rainfall to adult flight activity; and of soil moisture to oviposition behavior and egg and first instar survival in Phyllophaga crinita. Environ. Entomol. 8:591–594. [ Links ]
Harding, W. C. Jr.; Hartsock, J. G. and Rohwer, G. G. 1966. Blacklight trap standards for general insect surveys. Bull. Entomol. Soc. Am. 12:31–32. [ Links ]
Hayes, Wm. P. and McColloch, J. W. 1924. The biology of Anomala kansana (Scarabaeidae, Coleop.) J. Econ. Entomol. 17:589–594. [ Links ]
Ratcliffe, B. C. 1991. The scarab beetles of Nebraska. Bull. Univ. Nebr. Stati. Mus. 12:1–333. [ Links ]
Ritcher, P. O. 1958. Biology of Scarabaeidae. Annu. Rev. Entomol. 3:311–334. [ Links ]
Reitz, S. R. and Trumble, J. T. 2002. Competitive displacement among insects and arachnids. Annu. Rev. Entomol. 47:435–465. [ Links ]
Rodríguez–del–Bosque, L. A. 1980. Evaluación de pérdidas en maíz por plagas del suelo en el norte de Tamaulipas, México. Folia Entomol. Mex. 45:97–98. [ Links ]
Rodríguez–del–Bosque, L. A. 1982. Aspectos sobre la biología y comportamiento de la gallina ciega, Phyllophaga crinita Burm. (Col.: Scarabaeidae). Folia Entomol. Mex. 54:43–44. [ Links ]
Rodríguez–del–Bosque, L. A. 1984. Oviposición de Phyllophaga crinita Burmeister sobre diferentes cultivos en el norte de Tamaulipas, México. Southwest. Entomol. 9:184–186. [ Links ]
Rodríguez–del–Bosque, L. A. 1988. Phyllophaga crinita Burmeister (Coleoptera: Melolonthidae): Historia de una plaga del suelo (1855–1988). Memorias de la III mesa redonda sobre plagas del suelo. Soc. Mex. Entomol., 24 de Mayo, Morelia, Michoacán, México. p. 53–79. [ Links ]
Rodríguez–del–Bosque, L. A. 1993. Abundancia estacional y ecología de coleópteros rizófagos: Un estudio durante 15 años en agroecosistemas del norte de Tamaulipas, México. En: M. A. Morón (ed.) Diversidad y manejo de plagas subterráneas. Publicación Especial de la Sociedad Mexicana de Entomología e Instituto de Ecología. p. 7–15. [ Links ]
Rodríguez–del–Bosque, L. A., R. L. Crocker, and E. J. Riley. 1995. Diversity and abundance of Phyllophaga and Anomala in agroecosystems of northern Tamaulipas, Mexico. Soutwest. Entomol. 20:55–59. [ Links ]
Rodríguez–del–Bosque, L. A. 1996a. Seasonal feeding by Phyllophaga crinita and Anomala spp. (Coleoptera: Scarabaeidae) larvae in northeastern Mexico. J. Entomol. Sci. 31:301–305. [ Links ]
Rodríguez–del–Bosque, L. A. 1996b. Pupation and adult longevity of Phyllophaga crinita, Anomala flavipennis and A. foraminosa (Coleoptera: Scarabaeidae). Southwest. Entomol. 21:55–58. [ Links ]
Rodríguez–del–Bosque, L. A. 1998. A sixteen–year study on the bivoltinism of Anomala flavipennis (Coleoptera: Scarabaeidae) in Mexico. Environ. Entomol. 27:248–152. [ Links ]
Teetes, G. L., Wade, R. J., McInyre, C. and Schaefer, C. A. 1976. Distribution and seasonal biology of Phyllophaga crinita in the Texas High Plains. J. Econ. Entomol. 69:59–63. [ Links ]

