










Serviços Personalizados
Journal
Artigo
 Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkBoletín de la Sociedad Botánica de México
versão impressa ISSN 0366-2128
Bol. Soc. Bot. Méx no.88 México Jun. 2011
Ecofisiología
Microclimate and sapling survival under organic and polyethylene mulch in a tropical dry deciduous forest
Microclima y supervivencia de plántulas bajo acolchados orgánicos y de polietileno en un bosque tropical caducifolio
María Guadalupe Barajas–Guzmán and Víctor L. Barradas1
Universidad Nacional Autónoma de México, Instituto de Ecología. 1 Corresponding author: vbarrada@ecologia.unam.mx
Received: December 20, 2010.
Accepted: April 11, 2011.
Abstract
Mulches effect on net radiation (RN), air (Ta) and soil (TS) temperature, vapour pressure deficit (VPD), soil water content (SW), stomatal conductance (gS) and survival of three native species in a degraded tropical deciduous forest area was examined. The study year was dry, with half of the average annual rainfall. Three blocks were chosen, each block contained sixteen plots. Mulches with alfalfa straw, forest litter, white polyethylene and bare soil, were used. Twenty plants of each species one year–old were transplanted in each of 16 plots (5 × 6 m). The experiment was conducted during one year. RN was higher in plots with forest litter than in other treatments; Ta, TS and VPD were higher in bare soil plots than in mulched plots, and SW was higher in mulched plots than in bare soil plots. Ipomoea wolcottiana and Caesalpinia eriostachys registered the highest values of gS and survival, whilst Lonchocarpus eriocarinalis registered the lowest. In polyethylene mulch the three species showed the greatest survival. Mulches mitigate extreme microclimates and increase plant survival during the restoration of dry tropical areas.
Key words: low precipitation, microclimate mitigation, native plants, reforestation, stomatal conductance.
Resumen
Se examinó el efecto de acolchados en la radiación neta (RN), temperatura del aire (Ta) y del suelo (TS), déficit de presión de vapor (DPV), contenido de agua en el suelo (AS), conductancia estomática (gS) y supervivencia de tres especies nativas en un área degradada de bosque tropical caducifolio. El año de estudio fue seco, con la mitad del promedio de la precipitación anual. Se establecieron 48 parcelas con acolchados de alfalfa, mantillo, polietileno blanco y suelo desnudo. Se transplantaron 20 plantas de cada especie de un año de edad en cada una de las parcelas (5 × 6 m). El experimento se llevó a cabo durante un año. RN fue más alta en los acolchados de mantillo que en los otros tratamientos; AS fue más alto en los acolchados que en el control, mientras que Ta, TS, y DPV fueron más altos en el suelo desnudo. Ipomoea wolcottiana y Caesalpinia eriostachys presentaron las gS y supervivencias más altas, y Lonchocarpus eriocarinalis las más bajas. Las tres especies registraron la mayor supervivencia en polietileno. Los acolchados mitigan los microclimas extremos e incrementan la supervivencia vegetal al restaurar las áreas tropicales secas.
Palabras clave: baja precipitación, conductancia estomática, mitigación del microclima, plantas nativas, reforestación
Deforestation is one of the most serious environmental problems in the world mainly throughout the tropics, this is estimated in tropical areas between 110,000 and 240,000 km2 year–1, however, reforestation only includes 11,000 km2 year–1 (PNUMA, 2002). The demands made on forest resources by the human and animal (cattle raising) populations, further accentuated by climatic fluctuations, have lead not only to the deterioration of the ecosystems in which they are found but also to genetic exhaustion of many vegetal species (Cony, 1995). Furthermore, it is necessary to add the destruction and fragmentation of the natural habitat and the acceleration of soil loss due to hydric and aeolian erosion. These processes provoke climate change on a micro and intermediate level (Barradas and Fanjul, 1986) and a loss of soil fertility (Maass and García–Oliva, 1990), thus generating a new and more extreme microenvironment, which limits reforestation, reduces land productivity and leads to the degradation of natural resources.
In Mexico the deforestation is approximately 6,680 km2 year–1 (Masera et al., 1997) and reforestation is only 334 km2 year–1 (Carabias and Arizpe, 1993), and survival of plants is as much as 35% (Arriaga et al., 1994). The tropical dry deciduous forests constitute approximately 60% of Mexico's tropical vegetation (Rzedowski, 1978) and they have not been studied as much as conifers and rainforests, despite the fact that 3,020 of the 6,680 km2 annually deforested belong to the former category (Masera et al., 1997).
When tropical forests are deforested the microclimate is extremely severe (high incoming radiation, high evaporation, elevated and strong seasonally and daily variations in temperature, low air humidity) (Barradas and Fanjul, 1986), and these conditions affect stomatal conductance a physiological key for the growth and survival of plants. In general, stomatal closure occurrences in response to decreasing light and can be enhanced by water stress, or in response to increased leaf–to–air vapour pressure difference; also stomata aperture is dependent on many other factors such as leaf age, nutrition and disease (Jones, 1992).
All these concerns have lead to the need to explore different and diverse techniques for ensuring the successful survival of the plants used in reforestation programs and the restoration of ecosystems. One of these is mulching, an agricultural technique which could favour the success of the planting, survival and growth of plants during reforestation, because it protects the soil structure from the impact of raindrops, reduces changes in temperature (Price et al., 1998), and the rate of evaporation decreases, while infiltration rate increases, losses due to erosion and water running off the surface are reduced (Rathore et al., 1998). However, in areas with water deficiencies, very little is known about the influence of mulches on the physical variables of the soil, as well as the changes in the microclimate and the plants survival (Tilander and Bonzi, 1997).
In this study, we investigated the effects of mulching on survival saplings of tropical dry deciduous forest (TDF) trees and on air and soil microenvironment in a perturbed TDF area of the Pacific coast of Mexico. Our aim was to determine the effects of different mulches (polyethylene, alfalfa straw, and forest litter of a mature TDF) on sapling stomatal conductance and survival, soil water content, and the air and soil temperature, the deficit pressure vapor and net radiation.
Materials and methods
Site description. The study was carried out in the village of San Mateo, on the Pacific coast of Mexico in the state of Jalisco (19° 34' N, 105° 04' W), in a lowland cattle raising area where TDF was the original vegetation and was removed approximately 15 years ago. The area had previously been used for maize cultivation, as is common in these seasonally dry areas. The landscape of this area is presently covered by Acacia farnesiana (L.) Wild (500 trees per ha) and some Caesalpinia eriostachys Benth (one tree per ha), with plants of the Asteraceae family dominating the herbaceous stratus.
The climate in the region is characterized by a seasonal rainfall pattern, with a rainy season (June to October) that provides 90% of the total annual rainfall (mean annual rainfall average 740 mm; García–Oliva et al., 1991). Monthly air temperatures are in the range 22–27 °C, with a diurnal variation of 9 °C in summer and 14 °C in winter (de Ita–Martínez and Barradas, 1986). During the study year the cumulative precipitation was 391.6 mm (i.e., approximately 50% of the long–term mean annual rainfall), and the annual average temperature minimum was 17.2 °C and the maximum 28.8 °C.
The landscape in the area is characterized by convex hillside located 80 m asl facing south; its slope ranges from 5 to 25°. The soil (Haplic Ustarents) is shallow (generally less than 0.6 m in depth), fine and kaolinitic; the parent material is rhyolitic volcanic rock (Campo et al., 2001). The proportions of clay, silt, and sand in the soil to a depth of 10 cm were respectively 25 ± 3, 24 ± 3, and 51 ± 3% (mean ± 1 SE).
The vegetation in the region is the TDF (Rzedowski, 1978). The forest is diverse in species composition, with approximately 750 species grouped in 108 families. Leguminosae is the most common family, accounting for 15% of species (Lott et al., 1987). The more common species are Caesalpinia eriostachys Benth Lonchocarpus eriocarinalis Micheli, Ipomoea wolcottiana Rose, Bursera instabilis McVaugh & Rzedowski, Jatropha malacophylla Standl., Croton chamelensis E.J. Lott, Cordia alliodora (Ruiz & Pav.) Oken and Spondias purpurea L.
Experimental design. The experiment was carried out from July 2001 to July 2002, in a 90 × 50 m area on a slope facing mainly south (the slope angle ranges from 5 to 25°). Three blocks were established; each block contained 16 plots of size 5 × 6 m in which three types of mulch were placed (alfalfa straw, forest litter and white polyethylene) as well as control plots of bare soil. The blocks were spaced 2 m apart, and the plots were spaced 1.5 m apart, and set out lengthwise at 4 m intervals along the slope. To prevent possible leaching effects and water running from one plot into another, canals were dug (85 m long, 0.75 m wide and 0.60 m deep) running perpendicular to the slope.
Each plot with organic mulch was covered with 900 g dry matter per m2 (forest litter or alfalfa straw; Medicago sativa L.). This amount of mulch covered between 70 and 75% of the soil area. Medicago sativa is abundant in the region from cattle raising practices. The forest litter was a mixture of the TDF plant species and was collected from the soil of the forest in the dry season at the end of May 2001. The forest litter consisted of all dead plant material lying on the forest floor, including freshly fallen forest litter and the more finely decomposed forest litter fraction. Saplings of three native species were used: Ipomoea wolcottiana Rose (Convolvulaceae), Lonchocarpus eriocarinalis Micheli (Leguminoseae) and Caesalpinia eriostachys Benth (Leguminoseae). Ipomoea wolcottiana is a deciduous early successional species with fast growth rate; L. eriocarinalis a deciduous intermediate successional and N–fixer species with an intermediate growth rate, and C. eriostachys is a facultative deciduous, late successional species with a slow growth rate (Huante et al., 1995). Twenty one–year–old plants (0.66 plant m–2), were transplanted into each plot with mulch or bare soil; a total of 240 plants of each species were transplanted on July 22–25 2001, with 1.6 m distance between them. Each plot contained a single plant species. The experimental design was therefore a combination of 4 (3 mulches and bare soil) × 4 (3 species and without plants) factors with three replicates (3 blocks) = 48 plots. The selection of species was based on their high abundance in TDF (Lott et al., 1987), their growth rates (Huante et al., 1995), and high production of seeds and high germination percentage (I. Acosta, pers. comm.).
Mulch characteristics. The mulches differed in their color, polyethylene was white, alfalfa straw (Medicago sativa) was light yellow and TDF forest litter was dark brown. The polyethylene besides was impermeable with thickness of 0.4 and the mean time of utility as mulching was of six months. The polyethylene was perforated where plants were transplanted with a diameter of 25 cm and fastened with stones.
Net radiation, air temperature and humidity. All measurements were made in three periods of ten days each one, on September, October, and November of 2001 year. In the first moment of measurements, net radiation was registered every hour from 7:00 to 17:00 local standard time (LST) with a net radiometer (Q7_1, REBS, Campbell Scientific, Logan, Utah, USA) in three different mulches and bare soil plots.
Net radiation, air temperature and relative humidity were measured from 11:00 to 13:00 LST (period when maximum net radiation was registered) in all mulches and bare soil plots in all three times of measurements at 30–35 cm above the soil surface with a net radiometer (Q7_1, REBS, Campbell Scientific, Logan, USA) and a temperature and humidity probe (HMP35C, Vaisala, Helsinki, Finland) connected to a data logger (21X, Campbell Scientific, Logan, Utah, USA).
Vapour pressure deficit (VPD) was calculated from air temperature (Ta) and relative humidity (RH) from the relationship VPD = eS (1 – RH), where eS is vapour pressure at saturation and RH is given from zero to one.
Soil temperature and soil gravimetric water content (SW). Soil temperature at surface (TS0) and at 10 cm depth (TS10) were measured between 11:00 and 13:00 LST, with a temperature probe (HMP107, Campbell Scientific, Logan, Utah, USA) connected to a data logger (21X Campbell Scientific, Logan, Utah, USA) in mulches and bare soil plots.
For the evaluation of SW three soil samples (5–10 cm depth) were randomly collected from within each plot, combined in the field, and stored at 4 °C for up to 48 h until processing. The upper 5–10 cm of the soil profile in this region concentrates root biomass (~40% of total root biomass in the upper 0.6 m of the soil profile, or until rock contact; Barajas–Guzmán, unpublished data). In the laboratory, soils were hand homogenized and sieved (to 2 mm) for measurement of SW, determined by the differential weight between field moist and dried samples at 105 °C during 48 h.
Sampling stomatal conductance. Similarly to net radiation, stomatal conductance was measured every hour from 7:00 to 17:00 LST in the first period of measurements in at least three leaves of three saplings of the study species in each one plots with a steady state porometer (LI–1600, LI–COR, Lincoln, Nebraska, USA). Afterwards, stomatal conductance (gS) was only measured between 11:00 and 13:00 LST when maximum stomatal conductance (gSMAX) was registered.
Sapling stuvival. Sapling survival (as percentages from the initial values) was registered in rainy season (2001) and at the end of the experiment (July 2002). During the dry season it was not possible to determine if plants were alive with exactitude since they were leafless.
Statistical analysis. A non–parametric statistical analysis (Kruskal–Wallis and Friedman test) was used to analyse daily net radiation. A two–way (mulch and time) analysis of variance (ANOVA) was applied to the microclimate variables, and a three–way (mulch, time and species) analysis of variance was applied to stomatal conductance and sapling survival (these data were transformed to arcsines Vsurvival percentage) (Zar, 1999). Each plot included the slope as a co–variable. Finally, multiple regression analysis was applied between stomatal conductance and microclimate and soil variables.
Results
Microclimate. Daily net radiation was different between mulch treatments (P < 0.01), and the largest difference was registered between 11:00 and 13:00 LST. Cumulative values of net radiation from 7:00 to 17:00 LST indicate that the mulch with the lowest net radiation value was polyethylene (3.28 MW m–2), followed by bare soil (3.74 MW m–2) and alfalfa straw (3.96 MW m–2). The forest litter showed the highest value of net radiation (4.29 MW m–2).
The net radiation registered between 11:00 and 13:00 LST was different between mulches (P < 0.0001). All treatments were different between them, and again polyethylene showed the lowest (478.8 ± 16.4 W m–2) net radiation value, followed by bare soil (566.4 ± 12.9 W m–2), alfalfa straw (584.8 ± 13.5 W m–2), and on forest litter (601.2 ± 15.2 W m–2). Differences in net radiation were also found at different measurements (P < 0.0001). The highest radiation level was found in September (671.0 ± 6.3 W m–2), and was different of October (524.0 ± 10.9 W m–2) and November (478.3 ± 5.3 W m–2). The interaction between mulches and periods of measurement was significant (P < 0.001), and the lowest net radiation values was observed in plots with polyethylene mulch in October and November and the highest values with forest litter and alfalfa straw in September.
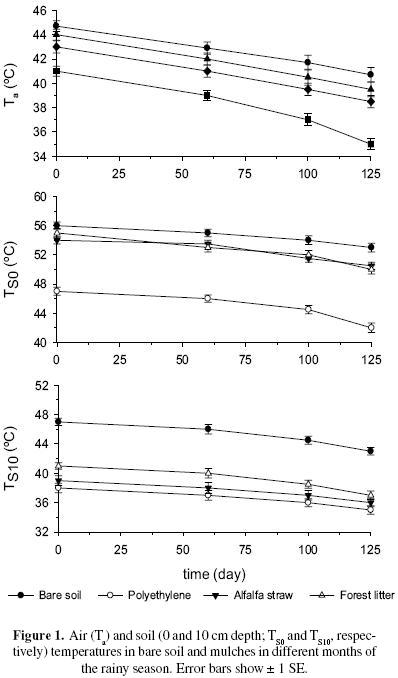
The air temperature differed among mulches (P < 0.0001) (Figure 1). Plots with polyethylene registered the lowest temperature and was different of the others treatments, alfalfa straw and forest litter were similar, and the highest temperature was registered on bare soil, this treatment and forest litter were not different. Significant differences were also registered over time (P < 0.00001). September was the hottest period (43.7 ± 0.4 °C), and was different of October (40.7 ± 0.2 °C) and November (40.6 ± 0.3 °C). The interaction mulches x measurement was significant (P < 0.01); the lowest air temperature was registered in plots with polyethylene mulch in November and the highest in bare soil in September.
Significant differences of VPD were found between mulches (P < 0.0001); polyethylene mulch presented the lowest VPD (4.45 ± 0.11 kPa), and was different of bare soil plots (5.67 ± 0.15 kPa). Forest litter (5.36 ± 0.09 kPa) and alfalfa straw (5.14 ± 0.11 kPa) were not different. All times of measurements were different (P < 0.0001), the highest VPD (5.62 ± 0.15 kPa) was registered in September, followed by November (5.22 ± 0.08 kPa) and the lowest VPD was registered in October (4.64 ± 0.07 kPa). The interaction mulches x periods of measurement was significant (P < 0.001), the lowest VPD values were observed in plots with polyethylene mulch in October and November and the highest in bare soil in September.
Soil temperature between 11:00 and 13:00 at the soil surface TS0 was different among mulches (P < 0.0001) (Figure 1) and between periods (P < 0.05). The greatest difference on TS0 (8.3 °C) was found between polyethylene mulch, with the lowest temperatures, and bare soil, with the highest temperature. Forest litter and alfalfa straw plots presented medium values. October showed the highest TS0 (51.8 ± 0.6 °C), and September and November (50.5 ± 0.5 °C and 50.5 ± 0.7 °C) were similar. The interaction mulches and times of measurement was significant (P < 0.05), and the lowest soil surface temperature was observed in soils under polyethylene mulch in November and the highest in bare soil in October and November.
Soil temperature at 10 cm depth (TS10) between 11:00 and 13:00 was different among mulches (P < 0.00001) (Figure 1), and periods of measurements (P < 0.00001). TS10 greatest difference (8.0 °C) again was found between polyethylene and bare soil, whereas alfalfa straw was similar to polyethylene and forest litter was different of others treatments. October and November showed the highest temperatures (41.3 ± 0.5 °C and 42.6 ± 0.6 °C) whereas September the lowest (35.5 ± 0.5 °C). The interaction mulches and measurement was significant (P < 0.01), and the lowest soil temperature at depth of 10 cm was in soils under polyethylene mulch in September and the highest in bare soil in October and November.
Soil gravimetric water content (SW) registered differences among mulches (P < 0.00001) and periods (P < 0.00001) (Figure 2). Polyethylene retained more water in the soil, followed by forest litter, whereas alfalfa straw retained as much water in the soil as forest litter, and bare soil showed the lowest values. SW was higher in September than October and November. However, soil was drier in July when soil was uncovered (Figure 2). The interaction between mulches and measurement was significant (P < 0.01), the highest SW was observed in soils under polyethylene mulch in September and the lowest in bare soil in November.

Sapling. The mulches affected maximum stomatal conductance (gSMAX) (P < 0.01) (Figure 3), plants with forest litter and polyethylene mulches were able to open more their stomata than plants on bare soil and with the less opening in plants with alfalfa straw mulch. The statistical analysis also showed that gSMAX was different among periods (P < 0.00001) (Figure 3), the highest values were registered on September to decrease on October and November. There were not interactions between the studied factors. The gSMAX also was different among species (P < 0.0001), Caesalpinia eriostachys registered the highest value of gSMAX, followed by Ipomoea wolcottiana and Lonchocarpus eriocarinalis (Figure 3).

Multiple regression analysis showed that gSMAX was controlled mainly negatively by air temperature and vapour pressure deficit, and positively by soil gravimetric water in Caesalpinia eriostachys (gSMAX = 4.30SW – 28.3Ta – 6.10VPD + 1300; R2 = 84.46.2; P < 0.00001) and in Lonchocarpus eriocarinalis (gSMAX = 2.13SW –18.70T – 4.22VPD + 798; R2 = 71.16; P < 0.00001), and the stomatal conductance in Ipomoea wolcottiana was related negatively with vapor pressure deficit, and positively by soil gravimetric water and net radiation (gSMAX = 13.70RN + 4.5SW – 5.8VPD – 250; R2 = 84.2; P < 0.00001).
Survival saplings varied significantly among mulches (P < 0.00001), months (P < 0.00001) and species (P < 0.00001). Survival increased in the order bare soil < alfalfa straw = forest litter < polyethylene. In contrast survival decreased after eight months in all treatments and species survival followed the order Lonchocarpus eriocarinalis < Caesalpinia eriostachys < Ipomoea wolcottiana (Figure 4).

Discussion
Bare soil plots showed a high amount of net radiation and the highest values of VPD, and air and soil temperature, and the lowest of SW, these conditions may be unfavorable to sapling survival. Caesalpinia eriostachys, Lonchocarpus eriocarinalis and Ipomoea wolcottiana presented a range survival of 0 to 10% in this treatment, this range is very low (3–4 times less) in contrast to reported by Kennard et al. (2002) and Liberman and Li (1992) in mature forest of TDF. This fact may be explained by the importance of the extreme temperatures and low content of soil water in the sapling establishment and survival in dry areas. In this treatment the highest air and soil temperatures decreased the SW, and these events increased the saplings mortality. Soil mulch evidently mitigates of the microenvironment that surrounds the transplanted plants. This mitigation could be mainly due to the reduction of net radiation and the physical barrier to conduct heat to and from the soil.
Differences among net radiation levels by mulches could be due to the reflective power of the materials used; polyethylene possibly reflected most of the radiation and, therefore, showed the lowest net radiation values (Rosset et al., 2001), whilst forest litter, which was dark brown in color, absorbed a great amount of radiation, reflected less and, therefore, showed the highest net radiation values (Montague et al., 2000). Alfalfa straw mulch showed intermediate values because was yellow in color. Therefore, it is possibly to conclude that the mulch color had a direct effect on the quantity of registered net radiation (Price et al., 1998; Iziomon and Mayer, 2002).
The amount of net radiation has an effect on the temperature and humidity of the air and of the soil (Rosset et al., 2001). The lowest air temperature was registered in the polyethylene mulch and this could be due to the fact that this mulch showed the lowest net radiation readings as well as the fact that this material heats up a great deal and cools down slowly. Consequently, the two organic mulches tended to heat up less energy than polyethylene and cool down more quickly, releasing their energy (in the form of heat) into the air immediately surrounding them. These temperature readings and the availability of water influenced the VPD because higher temperatures and/or low air humidity lead to greater VPDs, so the plots containing polyethylene mulch had the lowest VPD, followed by alfalfa straw, and after that, the forest litter.
Differences in soil temperature also could have an effect on SW (Tilander and Bonzi, 1997) because higher temperatures lead to greater evaporation and transpiration (Acharya et al., 1998; Novak et al., 2000) and, indeed, there is a low SW in plots with high temperatures. Soil beneath polyethylene mulch retained the greatest amount of water because soil temperatures were lower and polyethylene prevented water from evaporating quickly (Rathore et al., 1998; Mathews et al., 2002). In the case of the organic mulches, the values of the three parameters described above fall between those shown by polyethylene mulch and bare soil, because, although they absorbed energy and showed a greater SW than bare soil, they still lacked the ability of the polyethylene to reduce water loss.
The analysis of the microenvironmental variables which was carried out over time, shows that the reduction in net radiation and air temperature could be due to the time of year because, as winter approaches (Woodward, 1987). Soil gravimetric water was also reduced as time advanced because the rainy season finished at the end of October (Monks et al., 1997; Price et al., 1998). Although stomatal conductance was greater in September, VPD showed the highest recordings in this month probably due to air temperature effect was more marked than the effect of stomatal conductance.
A multiple regression analysis showed that stomatal conductance was controlled mainly by VPD, SW, air temperature and net radiation (Montague et al., 2000; Lagergren and Lindroth, 2002; Cochard et al., 2002). Actually there is a great concern to determine the response of stomata to environmental variables (e.g. Meinzer et al., 1997), and in general the studies showed that the decrease of stomatal conductance has been attributed to an increasing VPD and air temperature (Schulze, 1986). In this study it is interesting that Caesalpinia eriostachys and Lonchocarpus eriocarinalis showed contrasting values of gSMAX, and has been affected by the same microclimate variables, air temperature, VPD and SW. However, stomatal conductance of Ipomoea wol–cottiana was not affected by air temperature.
Ipomoea wolcottiana and Caesalpinia eriostachys registered the highest values of stomatal conductance and also the highest values of survival, while Lonchocarpus eriocarinalis registered the lowest values of survival and stomatal conductance. In polyethylene mulch the stomatal conductance and SW were high, whilst air temperature, DPV, net radiation and soil temperature were low, and the three species showed greatest survival. The mitigation of the extreme microenvironment by the mulches, has repercussions on stomatal conductance and therefore on gas exchange with a direct effect on plant growth (Barajas–Guzmán et al., 2006) and indirectly on plant establishment and survival.
In conclusion, our results show that mulches absorbed and reflected radiation differentially, reduced evaporation from the soil, reduced soil temperature at the surface and at 10 cm depth, and increased the sapling survival. On the other hand, this study strengthens the idea that the stomatal conductance is a good predictor for the plants survival. Finally, the use of mulches is recommended to restore highly perturbed zones and special in those areas with high water deficit as they mitigate extreme microclimate conditions and limit direct evaporation from the soil.
Acknowledgements
We thank to L. Hernández, G. Prado, R. León and D. Juárez for technical assistance. This research was supported by a grant from DGAPA–PAPIIT, UNAM (IN204599) and CONACyT (MGBG).
Literature cited
Acharya C.L., Kapur O.C. and Dixit S.P. 1998. Moisture conservation for rainfed wheat production with alternative mulches and conservation tillage in the hills of north–west India. Soil and Tillage Research 46:153–163. [ Links ]
Arriaga–M V., Cervantes–G V. and Vargas–Mena A. 1994. Manual de Reforestación con Especies Nativas: Colecta y Preservación de Semillas, Propagación y Manejo de Plantas. Secretaria de Desarrollo Social, Instituto Nacional de Ecología, Universidad Nacional Autónoma de México, México, D.F. [ Links ]
Barajas–Guzmán M.G. Campo J. and Barradas V.L. 2006. Soil water, nutrient availability and sapling survival under organic and polyethylene mulch in a seasonally dry tropical forest. Plant Soil 287:347–357. [ Links ]
Barradas V.L. and Fanjul L. 1986. Microclimatic characterization of shaded and open–grown coffee (Coffea arabica L.) plantations in Mexico. Agricultural and Forest Meteorology 38:101112. [ Links ]
Campo J., Maass M. and de Pablo L. 2001. Weathering in a Mexican tropical dry forest. Agrociencia 35:245–254. [ Links ]
Carabias J. and Arizpe L. 1993. El deterioro ambiental: Cambios nacionales, cambios globales. En: Azuela A. Carabias J. Provencio E. and Cuadri G. Eds. Desarrollo Sustentable. Hacia una Política Ambiental. pp. 43–59. Universidad Nacional Autónoma de México, México, D.F. [ Links ]
Cochard H., Coll L., Le–Roux X. and Améglio T. 2002. Unraveling the effects of plant hydraulics on stomatal closure during water stress in walnut. Plant Physiology 128:282–290. [ Links ]
Cony M.A. 1995. Reforestación racional de zonas áridas y semiáridas con árboles de múltiples propósitos. Interciencia 20:249-252. [ Links ]
de Ita–Martínez C. and Barradas V.L. 1986. El clima y los patrones de producción agrícola en una selva baja caducifolia de la costa de Jalisco, México. Biotica 11:237–245. [ Links ]
García–Oliva F., Ezcurra E. and Galicia L. 1991. Pattern of rainfall distribution in the Central Pacific Coast of Mexico. Geogra–phiscar Annalizer. Series A, Physical Geography 73:179–186. [ Links ]
Huante P., Rincón E. and Acosta I. 1995. Nutrient availability and growth rate of 34 woody species from a tropical deciduous forest in Mexico. Functional Ecology 9:849–858. [ Links ]
Iziomon M. and Mayer H. 2002. On the variability and modelling of surface albedo and long–wave radiation components. Agricultural and Forest Meteorology 111:141–152. [ Links ]
Jones H.G. 1992. Plants and Microclimate. A Quantitative Approach to Environmental Plant Physiology. Cambridge University Press, Cambridge. [ Links ]
Kennard D.K., Gould K., Putz F.E., Fredericksen T.S. and Morales F. 2002. Effect of disturbance intensity on regeneration mechanisms in a tropical dry forest. Forest Ecology and Management 162:197–208. [ Links ]
Lagergren F. and Lindroth A. 2002. Transpiration response to soil moisture in pine and spruce trees in Sweden. Agricultural and Forest Meteorology 112:67–85. [ Links ]
Lieberman D. and Li M. 1992. Seedling recruitment patterns in a tropical dry forest in Ghana. Journal of Vegetation Science 3:375–382. [ Links ]
Lott E., Bullock H. and Solís–Magallanes A. 1987. Floristic diversity and structure of upland and arroyo forest of coastal Jalisco. Biotropica 19:228–235. [ Links ]
Maass M. and García–Oliva F. 1990. La investigación sobre la erosión de suelos en México. Un análisis de la literatura existente. Ciencia 41:209–228. [ Links ]
Masera O.R., Ordóñez M.J. and Dirzo R. 1997. Carbon emissions from Mexican Forest: Current situation and long–term scenarios. Climate Change 35:265–295. [ Links ]
Mathews C.R., Bottrell D.G. and Brown M.W. 2002. A comparison of conventional and alternative understory management practices for apple production: multitrophic effects. Applied Soil Ecology 21:221–231. [ Links ]
Meinzer F.C., Hinckley T.M. and Ceulemans R. 1997. Apparent responses of stomata to transpiration and humidity in a hybrid poplar canopy. Plant, Cell and Environment 20:1301–1308. [ Links ]
Monks C.D., Monks D.W., Basden T., Selders A., Poland S. and Rayburn E. 1997. Soil temperature, soil moisture, weed control and tomato (Lycopersicon esculentum) response to mulching. Weed Technology 11:561–566. [ Links ]
Montague T., Kjelgren R. and Rupp L. 2000. Surface energy balance affects gas exchange and growth of two irrigated landscape tree species in an arid climate. Journal of the American Society of Horticulture Science 125:299–309. [ Links ]
Novak M.D., Chen W. and Hares M.A. 2000. Simulating the radiation distribution within a barley–straw mulch. Agricultural and Forest Meteorology 102:173–186. [ Links ]
Price J., Rochefort L. and Quinty F. 1998. Energy and moisture considerations on cutover peatlands: surface microtopography, mulch cover and Sphagnum regeneration. Ecology Engineering 10:293–312. [ Links ]
Programa de las Naciones Unidas para el Medio Ambiente. 2002. Perspectivas del Medio Ambiente Mundial. GEO–3. Ed. Mundi–Prensa, Madrid. [ Links ]
Rathore A.L., Pal A.R. and Sahu K.K. 1998. Tillage and mulching effects on water use, root growth and yield of rainfed mustard and chickpea grown after lowland rice. Journal of the Science of Food and Agriculture 78:149–161. [ Links ]
Rosset M., Montani M., Tanner M. and Fuhrer J. 2001. Effects of abandonment on the energy balance and évapotranspiration of wet subalpine grassland. Agriculture, Ecosystems and Environment 86:277–286. [ Links ]
Rzedowski J. 1978. Vegetación de México. Limusa. México, D.F. [ Links ]
Schulze E.D. 1986. Carbon dioxide and water vapour exchange in response to drought in the atmosphere and in the soil. Annual Review of Plant Physiology 37:247–274. [ Links ]
Tilander Y. and Bonzi M. 1997. Water and nutrient conservation through the use of agroforestry mulches, and sorghum yield response. Plant Soil 197:219–232. [ Links ]
Woodward F.I. 1987. Climate and Plant Distribution. Cambridge University Press, Cambridge. [ Links ]
Zar J.H. 1999. Biostatistical Analysis. Prentice–Hall, Upper Saddle River. [ Links ]

