










Services on Demand
Journal

Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
 Cited by SciELO
Cited by SciELO Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkBoletín de la Sociedad Botánica de México
Print version ISSN 0366-2128
Bol. Soc. Bot. Méx n.83 México Dec. 2008
Taxonomía y florística
Plant diversity assessment in the Yaxchilán Natural Monument, Chiapas, Mexico
Evaluación de la diversidad vegetal en el Monumento Natural Yaxchilán, Chiapas, Mexico
Jorge A. Meave1,4, Marco A. Romero–Romero1, Andrés Valle–Doménech1,2, Armando Rincón–Gutiérrez1, Esteban Martínez3 and Clara H. Ramos3
1 Departamento de Ecología y Recursos Naturales, Facultad de Ciencias, Universidad Nacional Autónoma de México, México 04510, D.F., México. 4 Author for correspondence. E–mail: jorge.meave@ciencias.unam.mx
2 Present address: El Colegio de la Frontera Sur, Unidad Villahermosa, Villahermosa, Tabasco, México.
3 Departamento de Botánica, Instituto de Biología, Universidad Nacional Autónoma de México, México 04510, D.F, México.
Received: August 22, 2008.
Accepted: October 6, 2008.
Abstract
We assessed the diversity of vascular plants in the Yaxchilán Natural Monument, a nature protection area located in Chiapas State, southern Mexico. A checklist including 547 species (337 genera, 102 families) was produced by combining information derived from three sources: (1) an initial list derived from an ecological study on forest structure; (2) a systematic survey conducted over an 18–month period; and (3) an extra visit to the site as part of a more ambitious project covering the entire Lacandon rain forest region. February was the month in which the largest number of fertile vouchers was collected in two consecutive years. The families with greatest richness were Fabaceae (47 species), Orchidaceae (34), Rubiaceae (23), and Asteraceae (22). The most speciose genera were Piper (13 species), Psychotria (9), Adiantum and Peperomia (8). Trees were the largest growth form group (203 species), followed by terrestrial herbs (129). Most species (459) were encountered in primary, closed forest, but secondary vegetation contributed 125 species. An estimation of the total size of Yaxchilán flora obtained through the Chao 2 algorithm indicated that this protected area may host more than 850 plant species. Despite its relatively small area, the Yaxchilán Natural Monument hosts a considerable biodiversity, representing over 16.2% of the recorded flora in the entire Selva Lacandona region, and over 7.2% of the State flora.
Key words: biodiversity assessment, floristics, Lacandon rain forest, nature preserve, Usumacinta River, tropical rain forest.
Resumen
Se evaluó la diversidad de plantas vasculares presentes en el Monumento Natural Yaxchilán, un área natural protegida del estado de Chiapas, sur de México. La lista florística, que incluye 547 especies (337 géneros y 102 familias) de plantas vasculares, fue elaborada a partir de la compilación de información derivada de tres fuentes: (1) una lista parcial obtenida a partir de un estudio ecológico de la estructura del bosque en una parcela de 1 ha en esta localidad; (2) una exploración sistemática realizada en siete viajes al sitio en un periodo de 18 meses; y (3) una visita extra al sitio como parte de una investigación más ambiciosa que abarca toda la Selva Lacandona. Los mayores números de ejemplares fértiles por visita fueron recolectados en el mes de febrero de dos años consecutivos. Las familias con mayor riqueza de especies fueron Fabaceae (47 especies), Orchidaceae (34), Rubiaceae (23), y Asteraceae (22). Los géneros más ricos en especies fueron Piper (13 especies), Psychotria (9), Adiantum y Peperomia (8). La categoría de forma de crecimiento más grande fue la de los árboles (203 especies), seguido por el de las hierbas terrestres (129). La mayoría de las especies (459) fueron halladas en la vegetación de selva primaria, aunque la vegetación secundaria también tuvo una contribución sustancial (125 especies). Se estimó que el tamaño de la flora total en Yaxchilán puede ser de alrededor de 850 especies de plantas, según el procedimiento de Chao 2. A pesar de su tamaño relativamente pequeño, el Monumento Natural Yaxchilán alberga una biodiversidad vegetal considerable, cuyo número representa 16.2% de la flora en toda la Selva Lacandona y alrededor de 7.2% de la flora conocida para el estado.
Palabras clave: área natural protegida, evaluación de la biodiversidad, florística, río Usumacinta, Selva Lacandona.
Since the second half of the 20th century, the national strategy for biodiversity conservation in Mexico has focused largely on the establishment of nature protection areas. Currently, the conservation network of areas under federal protection (known as Sinanp, the National System of Protected Areas), which is under the responsibility of the Secretariat of the Environment and Natural Resources (SEMARNAT), encompasses an archipelago of 161 protected areas distributed across all ecological zones and in all 31 States plus the Federal District of the country (Conanp, 2008).
Based on the recognition of differences in conservation status, degree of human occupancy, size, and other features of the nature preserves, the Sinanp comprises a range of categories, including Biosphere Reserves, National Parks, Special Fauna and Flora Protection Areas, and other kinds of areas enjoying official protection. "Natural Monument" (Monumento Natural) is one of these categories, and is defined in the Mexican Environmental Regulations as any area where protection is justified based on the fact that it contains one or several unique natural components, and a large aesthetic, historic or scientific value. For these reasons they are given the status of absolute protection. Natural Monuments cannot be included in other categories because they lack the variety of ecosystems that usually characterizes them or because they are too small (Conanp, 2008).
Despite the importance and the potential benefits of this strategy for the conservation of the national fauna and flora, a full appreciation of the value, and particularly of the efficacy of nature protection areas in protecting the rich biodiversity of Mexico, cannot be fully achieved without having comprehensive inventories for all biological groups that occur and are able to maintain viable populations within their limits. in addition, implementation of management programs is required by law for all protected areas within Sinanp. However, in the case of several nature protection areas the lack of adequate biological inventories has hindered the making of these management plans. Ultimately, the risk is that the lack of a management plan may trigger some disturbing processes that may potentially result in the decline of the biological integrity of the protected area. Considering their small size compared to other nature protection areas included in the Sinanp, this is of particular concern in the case of Natural Monuments.
The protected status of Yaxchilán began with a Presidential Ordinance granted to an area of 2,261 ha on August 24, 1994 (SEDESOL, 1992; Anonymous, 1995). In 1997, a collaborative study was undertaken in the Yaxchilán Natural Monument (hereafter YNM), in order to provide a comprehensive assessment of its biological diversity based on inventory work for several animal groups, as well as for vascular plants (Meave and Luis–Martínez, 1999; Meave, 2008). The ultimate goal of this effort was to provide a comprehensive evaluation of the biological diversity existing in this area, with the hope that this would serve as the basis for a future management plan. In this paper we provide the first account of the vascular plant diversity of the YNM. The floristic checklist presented here adds to those already published for the avifauna (Puebla–Olivares et al., 2002), mammals (Escobedo–Morales et al., 2005), and the herpetofauna (Ferreira–García and Canseco–Márquez, 2006). The list is not limited to the taxonomic information, but also includes data on growth form and the timing of reproductive phenology.
Study site
Location and physical environment. The YNM is located in the State of Chiapas (Ocosingo county, southern Mexico; 16°50'29"–16°54'05" N; 90°56'48"– 91°00'38" W), in the easternmost portion of the region known as Selva Lacandona (Lacandon Rain Forest). The Natural Monument is located on the left bank of the Usumacinta River, at the point where it marks the Mexico–Guatemala border; part of the YNM territory is encircled by a large bend of the river, known as "Yaxchilán's omega" (figure 1). Based on discharge volume, the drainage basin of this river ranks no. 1 in Mexico (de Vos, 1994). At Yaxchilán the Usumacinta flows with a SE–NW direction towards the Gulf of Mexico.
Predominant surface geology is limestone of Upper Cretaceous age (SPP, 1981); under the tropical climate a karstic terrain has originated, creating a moderately abrupt landscape dominated by circular hills, short escarpments and dissolution basins (García–Gil and Lugo–Hupb, 1992; Ali–phat–Fernández, 1996). in the area surrounded by the river's omega, elevation ranges between 80 and 260 m a.s.l.; further south, in the portion of the YNM abutting the El Tornillo range, maximum elevation is around 320 m. Prevailing soils are shallow rendzinas with high clay and organic matter contents in the upper layer; shallow lithosols, less than 10 cm deep and having many rock outcrops also occur (SPP, 1981). Some areas located in low topographic positions are poorly drained and often become flooded during the rainy season. This heterogeneity seems to be common in the Selva Lacandona region, where considerable differences in soil depth and associated water retention capacity have been reported between the higher parts of the hills and the intervening lower terrain (Miranda, 1961; Siebe et al., 1996).
The Selva Lacandona is a region of high precipitation; water is mostly brought in by the Easterlies after their passage over the Gulf of Mexico. in late summer and early autumn precipitation peaks due to cyclonic disturbances originated both in the Gulf of Mexico and the Pacific Ocean. Climate in Yaxchilán is of the Amw"ig type, i.e. the least humid of tropical humid climates. Mean annual temperature is ca. 25.5(C and total annual precipitation ranges between 1,560 and 2,380 mm (Herbert–Pesquera, 1995), with a mean of ca. 1,950 mm (SPP, 1981). Winter precipitation accounts for over 10% of total annual rainfall (Cardoso–D., 1979).
Vegetation. The Selva Lacandona is the Chiapan portion of a large region, measuring ca. 3 million hectares, that was originally covered by a mosaic of rain and evergreen lowland and montane forests, which is shared between the states of Chiapas and small portions of Campeche and Tabasco in Mexico, and the abutting El Petén department in Guatemala (Martínez et al, 1994; Meave, 1995; Medellín, 1996). The Selva Lacandona is one of the largest rain forest regions in Mexico, and originally it covered nearly one and a half million hectares. Despite being considered as one of the most important centers of biodiversity in Mexico (de la Maza and de la Maza, 1991; Medellín, 1991; de la Maza, 1998) as a result of the extremely high deforestation rates in the past four decades, the extent of the rain forest has been reduced to less than 500,000 ha (Medellín, 1996), of which 331,200 are protected in the large Montes Azules Biosphere Reserve.
Despite a relatively low precipitation for tropical rain forest standards (Whitmore, 1990; Richards, 1996), the impression that vegetation in the YNM gives to first–time visitors is that of a lush, well–developed tropical rain forest. Trees about 50 m in height are common, particularly Ceiba pentandra and Ficus spp. However, after several walking expeditions through the area and a closer inspection of the vegetation, a different perspective is gained, as a considerable heterogeneity in many vegetation characteristics can be appreciated. The taller vegetation is found in ravines and in the many relatively small valleys that occur throughout the YNM. This widespread community may be classified as tropical evergreen forest (Breedlove, 1973; selva alta perennifolia after Miranda and Hernández–X. [1963], or bosque tropical perennifolio after Rzedowski [1978]). Elsewhere, however, the forest is not so tall; on upper slopes and hilltops the trees are shorter and never reach heights of 25 m, apparently as a consequence of the notably drier environment that results from the shallow soils on these slopes. This shorter community is also distinguished by a more open canopy, and may be classified as semi–evergreen medium–height forest (selva mediana subperennifolia after Miranda and Hernández–x. [1963] or bosque tropical subperennifolio after Rzedowski [1978]). It is likely that this shorter community has led some authors to classify the Yaxchilán vegetation in general as a medium–height forest (selva mediana) (Aliphat–Fernández, 1996). However, it is clear that the mature vegetation of the YNM should be better described as an intricate mosaic composed of patches of tall and medium–height forest that respond to topographic and edaphic variation across the landscape. Valle–Doménech (2000) performed a detailed analysis of this variation at the 1–ha level and found that both forest structure and floristic composition varies greatly between these two communities.
In periodically flooded areas located on the banks of the Usumacinta, a further plant community is present that may be classified as riparian or gallery forest. This community occupies a small area but has a distinctly different appearance, probably as a consequence of periodic disturbances due to the large seasonal fluctuations in water levels. Finally, in the area surrounding the research camp built by the National Institute of Anthropology (INAH) near the archeological zone and around the small landing strip existing there, vegetation displays a varying degree of human alteration, including one old orchard and some abandoned milpas (traditional maize fields).
Martínez et al. (1999) listed 13 vegetation types for the entire Selva Lacandona region including several variants and transitions; at a smaller scale, the vegetation occurring in Yaxchilán closely matches this mosaic pattern. From a phytogeographic perspective, Yaxchilán forms part of the Gulf of Mexico Floristic Province (Rzedowski, 1978); however, Wendt (1993) and Ibarra–Manríquez et al. (2002) consider the areas located in Tabasco, northern Chiapas, and western Campeche as transitional between the Gulf of Mexico and the Yucatan Peninsula floristic provinces.
As in other marginal rain forest regions, leaf phenology is characterized by a large proportion of trees shedding their leaves during the short dry season. In particularly dry years, the leafless period may last until mid June or early July. Again, deciduousness is more conspicuous and of longer duration in those areas where soil is shallower and vegetation is shorter and more open.
Human occupation. A remarkable feature of the YNM is the presence of the remains of an ancient city build by the lowland Maya people. Apparently, the first settlement in Yaxchilán took place sometime in the Late Preclassic Period (200 BC–AD 200). By AD 600, Yaxchilán had become a powerful city–state (García–Moll, 1996), probably assisted by its strategic location on the Usumacinta River, which may have been an important route for trade. Yaxchilán architecture is renowned for its fine carved stellae, altars and lintels (Garza and Tommasi, 1987). Like other lowland Maya cities, Yaxchilán was abandoned between 800 and 900 AD, at the time of the Maya classic collapse (García–Moll, 1996).
An interesting implication of human occupation of the site is the impact that the expansion of a major urban center such as Yaxchilán may have had on the natural environment. Many scholars have suggested that the abandonment of the classic Maya cities (see Challenger, 1998 for a review) resulted from extensive land degradation due to soil erosion from agricultural practices. In contrast, Gómez–Pompa (1993) claims that the productive systems used at that time did not affect soils irreversibly, and that deforestation cannot have been total, as the fast forest recovery could not have taken place without a multitude of natural or managed vegetation islands containing the necessary germplasm. The land around Yaxchilán was used for subsistence agriculture, likely based on corn, beans and squash polyculture, or for cash or other important crops, such as cotton, cacao and copal (an incense–like resin; Challenger, 1998). After abandonment, the vegetation recovered such that by the 20th Century, the forest structure described in the nearby archaeological zone in Bonampak was indistinguishable from that of in primary tropical forest found elsewhere in the Americas (Meave–del Castillo, 1990).
At the time of the European contact between the Americas and Europe, the Selva Lacandona was inhabited by a few choltí communities that were exterminated by the Spanish conquerors during the late 17th Century (de Vos, 1991, 1996). Thereafter, the region remained almost completely uninhabited except for a few Yucatec–Maya speaking groups who had little impact on the natural environment (Nations, 1988).
After the discovery of the ancient city of Yaxchilán in the early 19th Century (García–Moll and Juárez–Cossío, 1986), a massive exploitation of precious woods began in the Selva Lacandona during the second half of that century. The fluvial network of the Usumacinta and its tributaries served for transporting large pieces of mahogany (Swietenia macrophylla) and red cedar (Cedrela odorata) to the ports in the Gulf of Mexico. Several logging camps developed along the Usumacinta and its tributaries; Yaxchilán formed part of the territories leased to timber enterprises until the beginning of the 20th Century (de Vos, 1994), and must have suffered the impact of this activity, although this is uncertain (SEMARNAT, 2003).
At present, the community of Frontera Corozal, inhabited by people of the chol ethnic group and located 20 km upstream, has the largest influence on the YNM. The town was founded in 1976, at a time when an aggressive campaign by the Federal Government was taking place to colonize the rain forest border regions of southern Mexico (Diechtl, 1987). Currently, surveillance of the archeological zone and of the entire YNM is carried out by inspectors from Frontera Corozal.
Methods
Three sources of information were combined to prepare the checklist of vascular plants for the YNM. First, a modest set of specimens were collected as part of an ecological study in which heterogeneity of forest structure and composition were assessed in a 1–ha plot located in a central portion of Yaxchilán's "omega" (Valle–Doménech, 2000), relatively distant from the archeological site. These specimens were collected between October 1996 and June 1997. Second, as part of a large collaborative project, a systematic survey was conducted bimonthly over a 15–month period from December 1997 to February 1999. During this time, a minimum of three persons visited the site seven times for periods ranging between 8 and 12 days; they systematically walked through most of the territory of the YNM to collect samples. On one occasion, locations that were difficult to access in the western end of the preserve were reached by boat. Finally, an additional trip to the site was made in March 2005, as part of a more ambitious floristic project covering the entire Lacandon rain forest region.
Plant specimens were preserved by soaking them in a 50:50 alcohol/water solution and transported to the Universidad Nacional Autónoma de México (UNAM) in Mexico City. Species determinations were done by some of the authors, but many specialists in several plant groups (listed in table 1) took part in this process. The first voucher collection was deposited at MEXU, the National Herbarium of Mexico hosted at the Institute of Biology (UNAM). Additional sets will be distributed to other herbaria shortly.

The species in the checklist are organized according to the Cronquist System (with the modifications presented in Brummit, 1992) because of its wide use and comparability. Recent taxonomic revisions were used as authorities for the delimitation of species in some groups (e.g. Pteridophytes: Moran and Riba, 1995; Mickel and Smith, 2004; Lauraceae: Lorea–Hernández, 2002; Orchidaceae: Hágsater et al., 2005; Rubiaceae: Borhidi, 2006). When possible, genera nomenclature was standardized according to Brummitt (1992). Species author names follow Brummit and Powell (1992) and Villaseñor (2001); species names of others groups were verified in the Tropicos.org (2008) and IPNI (2008) web sites.
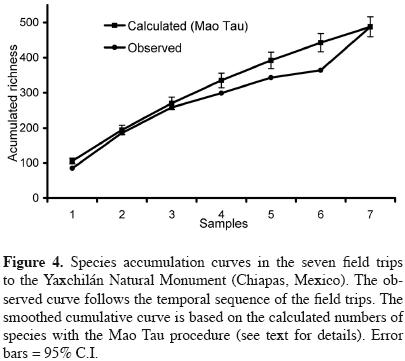
A smoothed species accumulation curve was constructed through randomization of the numbers of species collected on each trip with the Mao Tau procedure. This analysis used the data from the seven trips forming part of the systematic survey only, and was done with The Estimates software Ver. 8.0 (Colwell, 2006). The potential total size of the flora was obtained with the Chao 2 algorithm, which is based on species incidence (frequency) among samples. The seven collecting dates of the systematic survey were used as samples for this procedure. Incidence–based functions used to estimate total species richness usually require larger numbers of samples to guarantee unbiased results; therefore, the estimated total size of Yaxchilán flora should be viewed cautiously.
An initial evaluation of the conservation status of the flora of the YNM was obtained by identifying those species listed in the Mexican Official Norm of endangered species (SEMARNAT, 2002). Three categories are recognized in this Norm: threatened, endangered, and subjected to special protection.
Results
A total of 1,152 plant specimens were collected. Additionally, 29 species were sighted but it was not possible to obtain voucher specimens for them; their names are included in the checklist because their taxonomic identities were certain. The number of specimens determined to species level was 1,092. This gave a total of 547 species distributed in 337 genera and 102 families (appendix 1). Thus, the specimens/species ratio for the flora of YNM is slightly larger than 2. Most species (488) were collected during the seven expeditions of the longer systematic survey; the initial set of specimens collected in the 1–ha plot contributed 20 species, and the final collection trip added 17 more with specimens and a further 22 species that were observed but not collected (the lack of specimens for the latter is indicated on the checklist).
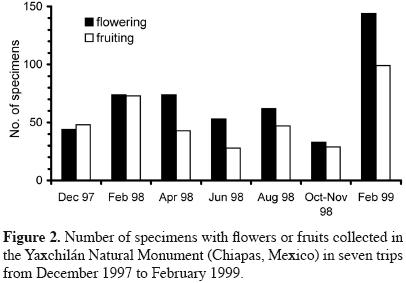
Flowering and/or fruiting specimens were found throughout the year; nonetheless, the yearly distribution of plants bearing reproductive structures was very irregular. Figure 2 shows the numbers of fertile specimens collected during each collecting expedition of the systematic survey: two peaks are evident, each in the month of February of two consecutive years. More remarkable, however, is the fact that the numbers of species in flower or fruit were different between this same month in the two years (132 specimens in 1998 vs. 219 specimens in 1999), despite the fact that collecting effort (number of collectors, duration of the trip) was almost identical. In contrast, October yielded the smallest number of fertile specimens with 57 species.
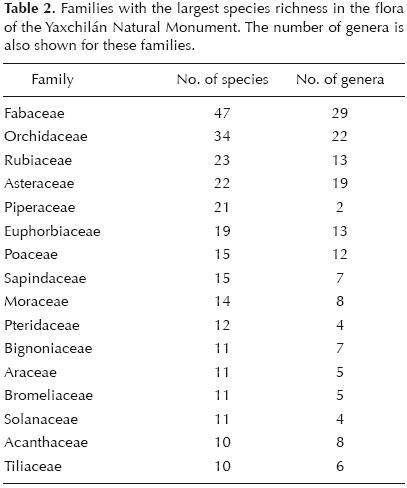
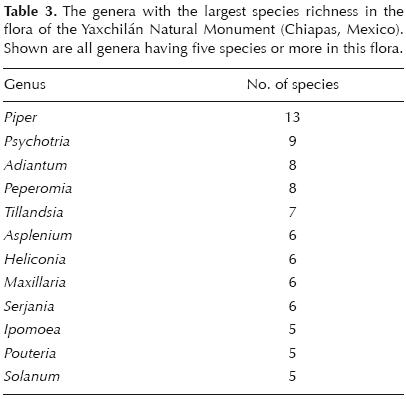
Fabaceae was the richest family, both by number of genera (28) and species (39) (table 2). Even so, this family only accounted for a relatively small percentage of the total recorded flora (7.6%). The family Orchidaceae was the second–richest family, both by genera (22) and species (34). While the Rubiaceae and the Asteraceae provided comparable species richness (23 and 22 species respectively), the Asteraceae had greater richness at the genus level (19) than the Rubiaceae (13). Other notable families contributing to species richness were Piperaceae (19, but only 2 genera), Euphorbiaceae (18 species, 13 genera), Sapindaceae (15 species, 7 genera), and Poaceae (15 species, 12 genera). On average, each family in the flora of the YNM is represented by 5.18 species and each genus by 1.56 species. Among the three most species–rich genera there were two belonging to the Piperaceae family (Piper, 11 species, and Peperomia, 8 species), and Psychotria (Rubiaceae) with 9 species. Table 3 lists all genera with five species or more in the flora of the YNM.
We classified all species collected in seven growth–form categories (figure 3). With 203 species, the category of trees was the best represented, but even this group accounted for a mere 37.1% of the total recorded flora. Ceiba pentandra is by far the most remarkable species due to its enormous heights (up to 50 m) and its abundance on the river banks, but other common canopy tree species are Ampelocera hottlei, Brosimum alicastrum, Manilkara zapota, Pouteria sapota and Terminalia amazonia. Beneath these trees, a large group of subcanopy tree species include Guarea glabra, Quararibea funebris, Sebastiania longicuspis, Stemmadenia donnell–smithii and Trichilia erythrocarpa. Finally, a well–represented group of short trees includes Rinorea hummelii, which is present in high density, but Chionanthus oblanceolatus, Garcinia macrophylla, Mouriri myrtilloides and Trichilia pallida are also common.
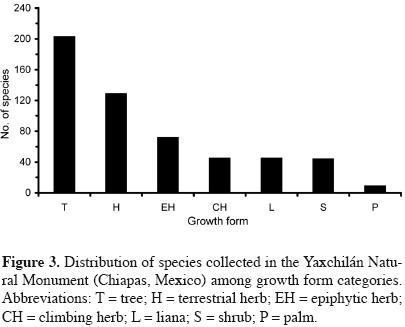
The group of terrestrial herbs was the second largest growth–form category, with 129 species (23.6%). These plants are poorly represented in the forest but common in places where vegetation has been disturbed. Common examples are several ferns (Adiantum spp., Asplenium spp., Ctenitis melanosticta, Hemionitis subcordata and Tectaria heracleifolia), as well as other forbs such as Aeschynomene americana, Costus pictus, Cyperus spp., Lasiacis divaricta, Oplismenus compositus, Porophyllum punctatum, Rene–almia mexicana and Tradescantia zanonia.
Epiphytic herbs were represented by an intermediate number of species (72, 13.2%; figure 3). Among them, species of the orchidaceae and Bromeliaceae were the most common, but there are also noteworthy ferns, Araceae and Piperaceae, particularly of the genera Aechmea, Anthurium, Epidendrum, Maxillaria, Oncidium, Peperomia, Polypodium, Scaphyglottis and Tillandsia.
The remaining groups of growth–forms combined, namely climbing herbs, shrubs, and lianas made a smaller and similar contribution to total richness. Examples of the herbs (45 species, 8.2%) include Dalechampia cissifolia, Matelea gentlei and Merremia umbellata; among the most conspicuous shrubs (45 species, 8.2%) are the following: Acalypha diversifolia, Critonia belizeana, Deherainia smaragdina, Justicia breviflora, Piper spp. and Psychotria limonesis. Common lianas (44 species, 8%) include Arrabidaea podopogon, Combretum fruticosum, Mascagnia vacciniifolia, Mikania leiostachya, Piptocarpha chontalensis and Tetracera volubilis.
The group of palms was represented by nine species only (1.7%), but they are worth mentioning because of their abundance. Particularly conspicuous palms belong to the Chamaedorea genus (C. elegans, C. ernesti–augusti, C. oblongata and C. tepejilote), although Attalea cohune, Cryosophila stauracantha and Geonoma interrupta are also common (A. cohune, a tall arborescent palm, is locally known as 'corozo', giving the name to the nearby community Frontera Corozal).
Many of the components of the flora of Yaxchilán show clear habitat preferences. The tall tropical rain forest located in deep soils was the vegetation type where the majority of specimens (896) were collected, and in which the largest number of species (459) occurred. This species subset includes, among other common species, the following: Alibertia edulis, Ampelocera hottlei, Bactris mexicana, Chionanthus oblanceolatus, Cryosophila stauracantha, Cymbopetalum penduliflorum, Dendropanax arboreus, Dialium guianense, Dieffenbachia oerstedii, Dracaena americana, Erythrina chiapasana, Faramea occidentalis, Garcinia macrophylla, Geonoma interrupta, Heliconia vaginalis, Licania platypus, Luehea seemannii, Mabea occidentalis, Mesadenella petenensis, Miconia impetiolaris, Mouriri myrtilloides, Nectandra martinicensis, N. salicifolia, Neea psychotrioides, Neomarica variegata, Pouteria durlandii, P. sapota, Pseudolmedia spuria, Psychotria chiapensis, P. limonensis, Pterocarpus rohrii, Rinorea hummelii, Simira lancifolia, Strychnos tabascana, Swartzia cubensis, Terminalia amazonia, Trichilia pallida, Tropidia polystachya, Vatairea lundellii and Vriesea heliconioides. In contrast, a smaller group of species tended to occur on the hilltops with shallower soils, where the environment is generally drier: Aspido–sperma megalocarpon, Chamaedorea elegans, Epidendrum ciliare, Manilkara zapota, Maxillaria aciantha, Mormolyca ringens, Ouratea lucens, Pimenta dioica, Randia aculeata, Trichilia minutiflora, Vitex gaumeri, Voyria parasitica and V. tenella.
Despite the relatively small area occupied by secondary vegetation in the YNM, 171 specimens representing 125 species were collected in this habitat. Many of these taxa live in fallows left by very limited agricultural activities in the recent past. Common species typical of the secondary vegetation include Acalypha diversifolia, Alchornea latifolia, Artocarpus altilis*, Attalea cohune, Bixa orellana, Bursera simaruba, Carica papaya, Cecropia peltata, Celtis iguanaea, Cnidoscolus multilobus, Costus pictus, Croton nitens, Dalechampia cissifolia, Delonix regia*, Desmoncus orthacanthos, Dioscorea bartlettii, Hamelia rovirosae, Heliocarpus americanus, H. mexicanus, Inga pavoniana, Iresine arbuscula, Mangifera indica*, Mikania leiostachya, Mucuna argyrophylla, Muntingia calabura, Piper yzabalanum, Plumeria rubra, Sapium lateriflorum, Schizolobium parahyba, Stemmadenia donnell–smithii, Terminalia catappa*, Tetracera volubilis, Thevetia ahouai, Trichospermum grewiifolium, T. mexicanum, Trophis racemosa and Zuelania guidonia. (This group includes several introduced species, marked with an asterisk on this list.) Similarly, those areas of highly–disturbed vegetation, namely the INAH camp and the small landing strip, contain a large group of species that thrive in these habitats, most notably Adelia barbinervis, Canna indica, Coix lacryma–jobi, Corchorus siliquosus, Desmodium scorpiurus, Heliotropium procumbens, Ipomoea quamoclit, Merremia umbellata, Muntingia calabura, Notylia aff. barkeri, Oplismenus compositus, Paspalum conjugatum, P. paniculatum, Piper fraguanum, Rhynchosia precatoria, Rivina humilis, Sinclairia deamii, Solanum americanum and Vigna vexillata.
The archeological zone, where the ancient buildings of the Mayan city are located, hosts an interesting combination of primary species as well as others typical of habitats with human disturbance. Among the most common ones, all likely to be encountered during a short visit to the zone, are Aechmea bracteata, Alchornea latifolia, Alseis yucatanensis, Aphelandra scabra, Asplenium cristatum, Brosimum alicastrum, Bursera simaruba, Castilla elastica, Cecropia peltata, Cedrela odorata, Ceiba pentandra, Chamissoa altissima, Coussapoa oligocephala, Dioscorea composita, Dorstenia contrajerva, Erythrina chiapasana, Hamelia rovirosae, Inga belizensis, Iresine arbuscula, Koanophyllon pittieri, Lasiacis divaricata, Leochilus scriptus, Maxillaria hedwigae, Nidema boothii, Peperomia nigropunctata, P. obtusifolia, Piper auritum, Protium copal, Pseudobombax ellipticum, Quararibea funebris, Rhipsalis baccifera, Ruellia pereducta, Spondias mombin, Stemmadenia donnell–smithii, Tillandsia schiedeana and Trichilia erythrocarpa.
The banks of the Usumacinta River provide habitats for plant species that are normally absent from forest vegetation with a closed canopy. These areas are particularly heterogeneous because some portions are regularly flooded during the rainy season, whereas others are only inundated infrequently, although all are subjected to disturbance by the river. Despite this heterogeneity, the narrow gallery forest contributed little to the flora of the YNM (four species, compared to the 19 species that were collected on the beaches that form during the low water period of the dry season along the Usumacinta River). Some conspicuous species of these environments are Amphitecna apiculata, Ceiba pentandra, Cyperus odoratus, Eragrostis hypnoides, Euphorbia hyssopifolia, Ficus glabrata, Guadua longifolia, Inga vera, Ludwigia leptocarpa, Mimosa pigra and Muntingia calabura.
While many species occurred in two vegetation types, only two species (Malvaviscus arboreus var. mexicanus and Cissus verticillata subsp. verticillata) occurred in three habitats (primary rain forest, secondary vegetation, and riparian habitats along the Usumacinta).
For Yaxchilán, as for any other protected area, the magnitude of biodiversity is an important question. Based on species collected during the systematic survey (figure 4), neither the observed nor the estimated cumulative species/ area curve (based on the Mau Tao procedure) showed a clear stabilization, suggesting that they would have continued to increase with additional collecting effort. The Chao 2 estimator, which is calculated based on information on species occurrence in the samples (the collecting trips), produced an estimated mean richness of 855.8 species (95% C.I. = 760.0 to 985.2). The mean value exceeds the observed richness by more than 56%.
According to the Mexican Norm of endangered taxa (SEMARNAT, 2002), two species occurring in the YNM are classified as requiring special protection (Pr): Tillandsia festucoides (Bromeliaceae), and Hamelia rovirosae (Rubiaceae). In turn, nine species are classified as threatened: two understorey palms (Chamaedorea ernesti–augusti and Cryosophila stauracantha), two Araceae (Dieffenbachia oerstedii and Monstera tuberculata), one Euphorbiaceae (Tetrorchidium rotundatum), one Clusiaceae (Calophyllum brasiliense), one Anacardiaceae (Astronium graveolens), one Acanthaceae (Bravaisia integerrima), and one Aspleniaceae (Asplenium serratum). Finally, one canopy tree species (Vatairea lundellii, Fabaceae) is considered to be in danger of becoming extinct. The number of taxa in this set (12 species) implies that only 2.2% of the flora of the YNM has received a protection status according to Mexican legislation.
Discussion
In agreement with its definition of Natural Monument, the protected area of Yaxchilán is very small (just over 2,600 ha) in comparison to other protected areas included in the Sinanp in Mexico, but also in comparison to many protected areas worldwide. Therefore, it is remarkable that this small protected area harbors a considerable plant diversity that encompasses almost 550 species of vascular plants. This figure is equivalent to 16.2% of the known flora (3,400 species) for the Selva Lacandona region (Martínez et al., 1994), and 7.2% of the more than 7,600 species enlisted by Breed–love (1986) for the entire State of Chiapas, one of the most diverse states in the country (Breedlove, 1973). The botanical information provided by this study agrees with the high levels of diversity recorded for other biological groups (235 bird species; Puebla–Olivares et al., 2002; 70 mammals species; Escobedo–Morales, 2005; 14 amphibian and 41 reptilian species; Ferreira–García and Canseco–Márquez, 2006).
The magnitude of the flora of YNM is equivalent to that recorded in Bonampak, one of the two other Natural Monuments forming part of Sinanp in Mexico. Meave–del Castillo (1990) reported 470 plant species for Bonampak, an area located less than 15 km from Yaxchilán, separated by the low elevation Cojolita range that divides the basins of the Lacanjá and Usumacinta rivers. The taxonomic information for Bonampak may be out of date, and a revision of the flora may be warranted to assess the degree of overlap and complementarity between these two well–preserved rain forest areas.
At present it is impossible to make a precise comparison of the size of the flora of the YNM with that of its enormous neighboring Montes Azules Biosphere Reserve, which covers an area larger than 330,000 ha (i.e. more than 127 times the size of the YNM); however, some data are available that allow an initial comparison. Castillo–Campos and Narave–Flores (1992) published a checklist including 984 species (distributed in 116 families) of vascular plants that were collected in eight localities spread over a large area of the Montes Azules Biosphere Reserve, all located along the Lacantún River and some of its major tributaries, or adjacent to lakes occurring in the reserve. Although these authors do not provide any indication of collecting effort, it is interesting that the known number of species for Yaxchilán is equivalent to 55.6% of the figure reported by them.
It must be acknowledged, however, that the YNM is by no means the richest tropical rain forest protected area in Mexico of equivalent size. In the Los Tuxtlas Biological Reserve (ca. 600 ha), located in Veracruz State, updated botanical information confirms the occurrence of 860 species of Magnoliophytes distributed in 506 genera and 119 families (Ibarra–Manríquez and Sinaca–Colín, 1997), plus 80 species of Pteridophytes sensu lato (Riba and Pérez–García, 1997), totaling 940 species of vascular plants. The difference in species richness between Yaxchilán and Los Tuxtlas likely also results from a much longer period of botanical exploration at the latter location (covering at least a couple of decades). Nonetheless, the difference may also be accounted for by the disparity in precipitation between the two regions: the Los Tuxtlas reserve receives an average precipitation of over 4,500 mm yearly, approximately 2.3 times that of the YNM. With a mean annual precipitation less than 2,000 mm, the climate of Yaxchilán is classified as wet seasonal (Walsh, 1996), which suggests that species with very high humidity requirements may be excluded from the forest, and in turn that many species present are more drought tolerant than most typical rain forest plants. These possibilities remain to be investigated.
It is also clear that the number of plants species in the YNM has not been fully tallied. The first indication of a larger flora for this site is the Chao 2 algorithm, which produced an estimated mean richness of 855.8 species, a figure similar to the known flora of Los Tuxtlas. Second, most of the field work was conducted in 1998 when a severe drought caused by a particularly strong El Niño event affected the southern part of Mexico, including the Yaxchilán region. In that year, a reduction in flowering intensity in Chajul was observed (S. Sinaca, com. pers.), a locality further south in Chiapas with a much larger annual precipitation than Yaxchilán. it is likely that this climatic event had a considerable negative impact; by causing a more intense water stress than usual during the period in which the flowering peak in this region takes place (February to May). In fact, the differences in number of flowering and fruiting species between February 1998 and February 1999 could be a further indication that 1998 was particularly dry. in regions marginal for rain forest development, as is the case of Yaxchilán, occasional very dry years may strongly limit the occurrence of species with very high water requirements (Jacobs, 1988); therefore, keeping adequate climatic records in the future may be very important to gain a better insight on the richness and floristic composition of the YNM.
Moreover, it cannot be denied that there was some bias in collecting plant specimens. A considerable effort was devoted to the collection of tree specimens; however, this bias may not have compensated for the very low densities of some tree species that are typical in rain forests, and, in fact, for many tree species only a single individual was encountered. However, the figure corresponding to trees in the YNM (203) is comparable to the 194 species reported by Ochoa–Gaona and Domínguez–Vázquez (2000) for Chajul, also in the Lacandon region. In Yaxchilán, terrestrial herbs richness ranked second, probably because they are easy to collect. In Chajul, however, shrubs (126 species) ranked second, greatly outnumbering the figure observed in the YNM; therefore, future botanical exploration in this reserve should pay more attention to this growth form. Also, more specimens were obtained both in areas of primary and disturbed vegetation in the vicinity of the archeology camp and much less time was allocated for the exploration of the most distant areas of the YNM, which are further away from the river. Additionally, limited access to the forest canopy (even though climbing gear was used to reach many canopy trees) may have concealed a considerable proportion of the epiphytic plants occurring in the area. A similar situation may be true for lianas, as derived from the contrast between the figure obtained for the YNM (45) and those reported by Ochoa–Gaona and Domínguez–Vázquez (2000) and Solórzano et al. (2002) for Chajul (72 and 128, respectively). In contrast, palms are very conspicuous elements of the flora and are likely very well represented in the checklist. These considerations highlight the need to continue the botanical exploration in Yaxchilán, paying particular attention to the canopy and the least accessible parts of the protected area.
An important criterion in assessing the protecting efficacy of the YNM for threatened plant species is the number of taxa included in the Mexican Norm. The small number of species (12) of the flora of the YNM that is listed in this environmental legal instrument may actually under–represent the risk situation in the flora of Yaxchilán; for example, there are many typical primary forest species that are practically absent from the large areas of the Lacandon Forest where vegetation has been altered or removed for agriculture and livestock, which are threatened by habitat destruction but not mentioned in the Official Norm. That is the case, among others, of Cojoba arborea (Mimosaceae), Dracaena americana (Agavaceae), Myroxylon balsamum (Fabaceae), Terminalia amazonia (Combretaceae), as well as several orchid species. In addition, there is at least one species (Swietenia macrophylla) that deserves special protection, given the need to maintain a diverse germplasm for an economically important tree species that for a long time was important in the economy of the tropical humid regions of Mexico (González–Pacheco, 1983; de Vos, 1991, 1994), and that is slowly gaining a new impulse through the establishment of plantations.
The evaluation of the potential for biological conservation in the YNM should not be constrained to its role in protecting different groups of species, but should also consider its capability to preserve particular habitats or whole ecological systems. González–Espinosa et al. (2005) provided a classification for the entire State of Chiapas in which 10 environmental groups were distinguished. Yaxchilán is located in the area corresponding to Environmental Group V, which is mostly distributed in the Lacandon Forest region, particularly within the Usumacinta River basin. Large areas of this region are presently deforested, especially in the Marqués de Comillas sector, and therefore the YNM provides an excellent opportunity to maintain at least a minimum representation of this environmental group.
The floristic survey reported here for the YNM confirms the large biological diversity that is being protected in this conservation unit. The present assessment, however, is insufficient, as the challenge is to maintain such large biodiversity in the future. By comparing land use/land cover changes within Mexican biological reserves and equivalent areas around them, Figueroa and Sánchez–Cordero (2008) found that the YNM has been an effective protection area since its creation, in contrast to many other areas of the Sinanp. In the Lacandon Forest region this is particularly important, as it has been demonstrated that the intensification of the slash–and–burn agriculture system, still practiced in this region, results in a continuous impoverishment of the local biological systems (Ochoa–Gaona et al., 2007). Every effort must be made to maintain the high effectiveness. In this context, it is critical that a legal management plan for the YNM is developed soon that will provide a legal framework to regulate human activities in Yaxchilán.
Acknowledgements
We thank the authorities of the Frontera de Corozal community for granting us permission to carry out this study in Yaxchilán; the assistance and hospitality of many people of this community is warmly acknowledged. The National Commission for Biodiversity in Mexico (Conabio) provided funds to carry out most of the field work in Yaxchilán (grant no. FB443/M099/97). We are grateful to the National Institute of Anthropology and History (INAH) for all the assistance provided, as well as for the permission to use their facilities at the archeological zone; we are particularly indebted to Mario Pérez Campa and Aidée Orea Magaña for their encouragement. Carlos Anaya, Liliana López, Efraín Tovar and Irene Sánchez–Gallen assisted in fieldwork. Continuous logistic support was provided by Manuel Pérez García, Marcos Cruz Díaz, Agustín López Arcos, Felipe Arcos Díaz, Gabriel Aguilar Méndez and Demetrio Álvarez Montejo. Collecting permits were provided by the Instituto Nacional de Ecología. Trudy Kavanagh kindly proof–read a previous version of this paper and improved our English. We express our gratitude to all plant specialists listed in table 1 for kindly providing and revising plant determinations; the taxonomic accuracy of our checklist depends very much on their generous collaboration.
Literature cited
Anonymous. 1995. Monumento Natural Yaxchilán. In: Gómez–Pompa A. and Dirzo R. Eds. Reservas de la Biosfera y Otras Áreas Naturales Protegidas de México, pp. 127–128, Instituto Nacional de Ecología (Secretaría de Medio Ambiente, Recursos Naturales y Pesca) and Comisión Nacional para el Conocimiento y Uso de la Biodiversidad, Mexico City. [ Links ]
Aliphat–Fernández M.M. 1996. Arqueología y paisajes del Alto Usumacinta. Arqueología Mexicana IV:24–29. [ Links ]
Borhidi A. 2006. Rubiáceas de México. Akadémia Kiado, Budapest. [ Links ]
Breedlove D.E. 1973. The phytogeography and vegetation of Chiapas (Mexico). In: Graham A. Ed. Vegetation and Vegetational History of Northern Latin America, pp. 149–165, Elsevier, New York. [ Links ]
Breedlove D.E. 1986. Listados Florísticos de México. IV. Flora de Chiapas. Universidad Nacional Autónoma de México, Mexico City. [ Links ]
Brummitt R.K. 1992. Vascular Plant Families and Genera. Royal Botanic Gardens, Kew. [ Links ]
Brummit R.K. and Powell C.E. (Eds.) 1992. Authors ofPlant Names. A List of Authors of Scientific Names of Plants, with Recommended Standard Form of Their Names, Including Abbreviations. Royal Botanic Gardens, Kew. [ Links ]
Castillo–Campos G. and Narave–Flores H. 1992. Contribución al conocimiento de la vegetación de la reserva de la biosfera Montes Azules, Selva Lacandona, Chiapas, México. In: Vázquez –Sánchez M.A. and Ramos–Olmos M.A. Eds. Reserva de la Biosfera Montes Azules, Selva Lacandona: Investigación para su Conservación. Centro de Estudios para la Conservación de los Recursos Naturales, A.C., San Cristóbal de las Casas, Chiapas. [ Links ]
Cardoso–D. M.D. 1979. El Clima de Chiapas y Tabasco. Instituto de Geografía, Universidad Nacional Autónoma de México. Mexico City. [ Links ]
Colwell R.K. 2006. EstimateS: Statistical estimation of species richness and shared species from samples. Version 8. <purl.oclc.org/estimates> [ Links ]
CONANP [Comisión Nacional de Áreas Protegidas]. 2008. Monumentos Naturales. <www.conanp.gob.mx/monumentos_naturales.html> Accessed on May 3rd, 2008. [ Links ]
Challenger A. 1998. Utilización y Conservación de los Ecosistemas Terrestres de México. Pasado, Presente y Futuro. Comisión Nacional para el Conocimiento y Uso de la Biodiversidad, Instituto de Biología (UNAM) and Agrupación Sierra Madre, S.C. Mexico City. [ Links ]
de la Maza J. 1998. El paisaje, la visión primigenia. In: Hernández–Obregón V.H. and Adé–Blanchard E. Eds. Selva Lacandona: Un Paraíso en Extinción, 2nd ed., pp. 29–47, Editorial México Desconocido, Mexico City. [ Links ]
de la Maza J. y de la Maza R. 1991. El "Monte Alto", esbozo de una región. In: Anonymous. Lacandonia: el Último Refugio, pp. 21–35, Universidad Nacional Autónoma de México, Mexico City. [ Links ]
de Vos J. 1991. Historia de la selva, crónica de una agresión. In: Anonymous. 1991. Lacandonia: el Último Refugio. pp. 37–51, Universidad Nacional Autónoma de México, Mexico City. [ Links ]
de Vos J. 1994. Oro Verde. La Conquista de la Selva Lacandona por los Madereros Tabasqueños, 1822–1949. Fondo de Cultura Económica, Mexico City. [ Links ]
de Vos J. 1996. Semblanza del río Usumacinta. Arqueología Mexicana IV:54–63. [ Links ]
Diechtl S. 1987. Cae una Estrella. Desarrollo y Destrucción de la Selva Lacandona. Secretaría de Educación Pública, Mexico City. [ Links ]
Escobedo–Morales L.A., León–Paniagua L, Arroyo–Cabrales J. and Polaco O.J., 2005. Diversidad y abundancia de los mamíferos de Yaxchilán, municipio de Ocosingo, Chiapas. In: Sánchez–Cordero V. and Medellín R.A. Eds. Contribuciones Mazto–zoológicas en homenaje a Bernardo Villa, pp. 283–298. Instituto de Ecología (UNAM) and Comisión Nacional para el Conocimiento y Uso de la Biodiversidad, México City. [ Links ]
Ferreira–García M.E. and Canseco–Márquez L. 2006. Estudio de la herpetofauna del Monumento Natural Yaxchilán, Chiapas, México. In: Ramírez–Bautista A., Canseco–Márquez L. and Mendoza–Quijano F. Eds. Inventarios Herpetofaunísticos de México: Avances en el Conocimiento de su Biodiversidad, pp. 293–310, Sociedad Herpetológica Mexicana, A.C., Mexico City. [ Links ]
Figueroa F. and Sánchez–Cordero V. 2008. Effectiveness of natural protected areas to prevent land use and land cover change in Mexico. Biodiversity and Conservation 17:3223–3240. [ Links ]
García–Gil J.G. and Lugo–Hupb J. 1992 Las formas del relieve y los tipos de vegetación de la Selva Lacandona. In: Vásquez–Sánchez, M.A. and Ramos M.A. Eds. Reserva de la Biosfera Montes Azules, Selva Lacandona: Investigación para su Conservación, pp. 34–39. Centro de Estudios para la Conservación de los Recursos Naturales, A.C., San Cristóbal de las Casas, Chiapas. [ Links ]
García–Moll R. 1996. Yaxchilán, Chiapas. Arqueología Mexicana IV:36–45. [ Links ]
García–Moll R. and Juárez–Cossío D. Eds. 1986. Yaxchilán: Antología de su Descubrimiento y Estudios. Instituto Nacional de Antropología e Historia, Mexico City. [ Links ]
Garza T. and Tommasi W. 1987. Atlas Cultural de México. Arqueología. Instituto Nacional de Antropología e Historia and Grupo Editorial Planeta, Mexico City. [ Links ]
Gómez–Pompa A. 1993. La silvicultura maya. In: Leff, E. and Carabias J. Coord. Cultura y Manejo Sustentable de los Recursos Naturales (Vol. II), pp. 367–384. Centro de Investigaciones Interdisciplinarias en Humanidades (UNAM) and Grupo Editorial Miguel Ángel Porrúa, Mexico City. [ Links ]
González–Espinosa M., Ramírez–Marcial N., Méndez–Dewar G., Galindo–Jaimes L. and Golicher D. 2005. Riqueza de especies de árboles en Chiapas: variación espacial y dimensiones ambientales asociadas al nivel regional. In: González–Espinosa M., Ramírez–Marcial N. and Ruiz–Montoya L. Eds. Diversidad Biológica en Chiapas, pp. 81–125, El Colegio de la Frontera Sur, COCyTECH, and Plaza y Valdés, Mexico City. [ Links ]
González–Pacheco C. 1983. Capital Extranjero en la Selva de Chiapas 1863–1982. Universidad Nacional Autónoma de México, Mexico City. [ Links ]
Hágsater E., Soto–Arenas M.A., Salazar–Chávez G.A., Jiménez–Machorro R., López–Rosas M.A. and Dressler R.L. 2005. Las Orquídeas de México, Instituto Chinoín, Mexico City. [ Links ]
Herbert–Pesquera L.L. 1995. Preservación de una Zona: Yaxchilán, Chiapas. B.A. Thesis, Escuela Nacional de Conservación, Restauración y Museografía, INAH–SEP–CONACULTA, Mexico City. [ Links ]
Ibarra–Manríquez G. y Sinaca–Colín S. 1997. Fanerógamas. In: González–Soriano E., Dirzo R. and Vogt R.C. Eds. Historia Natural de los Tuxtlas, pp. 162–174, Universidad Nacional Autónoma de México–CONABIO, Mexico City. [ Links ]
Ibarra–Manríquez G., Villaseñor J.L., Durán R. and Meave J. 2002. Biogeographical analysis of the tree flora of the Yucatan Peninsula. Journal of Biogeography 29:17–29. [ Links ]
IPNI [The International Plant Names] Index 2008. <www.ipni.org> Accessed in 2008. [ Links ]
Jacobs M. 1988. The Tropical Rain Forest: a First Encounter. Springer–Verlag, Berlin. [ Links ]
Lorea–Hernández F.G. 2002. La familia Lauraceae en el sur de México: diversidad, distribución y estado de conservación. Boletín de la Sociedad Botánica de México 71:59–70. [ Links ]
Martínez E., Ramos C.H. and Chiang F. 1994. Lista florística de la Lacandona, Chiapas. Boletín de la Sociedad Botánica de México 54:99–177. [ Links ]
Martínez E., Toledo V.M. and Ramos–Álvarez C.H. 1999. La vegetación de las Cañadas, Chiapas. Revista Chapingo, serie Ciencias Forestales y del Ambiente 5:15–26. [ Links ]
Meave J.A. 1995. La Selva Lacandona: una joya verde en peligro de desaparecer. Universidad de México L:11–15. [ Links ]
Meave J.A. 2008. Riqueza biológica y manejo del Monumento Natural Yaxchilán. Biodiversitas 78:12–15. [ Links ]
Meave J.A. and Luis–Martínez A.M. Eds. 1999. Caracterización biológica del Monumento Natural Yaxchilán como un elemento para el diseño de su plan rector de manejo. Final report (project FB443/M099/97) for the National Biodiversity Commission of Mexico (Conabio), Mexico City. <www.conabio.gob.mx/institucion/proyectos/resultados/InfM099.pdf> [ Links ]
Meave–del Castillo J. 1990. Estructura y Composición Florística de la Selva Alta Perennifolia en los Alrededores de Bonampak. Instituto Nacional de Antropología e Historia, Mexico City. [ Links ]
Medellín R.A. 1991. La fauna: diversidad de los vertebrados. In: Anonymous. Lacandonia: el Último Refugio, pp. 75–109, Universidad Nacional Autónoma de México, Mexico City. [ Links ]
Medellín R. 1996. La Selva Lacandona. Arqueología Mexicana IV:64–69. [ Links ]
Mickel J.T. and Smith A.R. 2004. The pteridophytes of Mexico. Memoirs of the New York Botanical Garden 88:1–1054 (328 plates). [ Links ]
Miranda F. 1961. Tres estudios botánicos en la Selva Lacandona, Chiapas, México. Boletín de la Sociedad Botánica de México 26:133–176. [ Links ]
Miranda F. and Hernández–X. E. 1963. Los tipos de vegetación de México y su clasificación. Boletín de la Sociedad Botánica de México 28:29–179. [ Links ]
Moran R.C. and Riba R. (Eds. in Pteridophytes) 1995. In: Davidse G., M. Sousa–S. and S. Knapp. Eds. Flora Mesoamericana Vol. 1 Psilotaceae a Salviniaceae. Instituto de Biología, Universidad Nacional Autónoma de México, Missouri Botanical Garden, The Natural History Museum (London), Mexico City. [ Links ]
Nations J.D. 1988. The lacandon Maya. In: Denslow J.S. and Padoch C. Eds. People of the Tropical Rain Forest, pp. 99–177. University of California Press, Berkeley. [ Links ]
Ochoa–Gaona S. and Domínguez–Vázquez G. 2000. Distribución y fenología de la flora leñosa de Chajul, Selva Lacandona, Chiapas, México. Brenesia 54:1–24. [ Links ]
Ochoa–Gaona S., Hernández–Vázquez F., de Jong B.H.J. and Gurri–García F. 2007. Pérdida de diversidad florística ante un gradiente de intensificación del sistema agrícola de roza–tumba–quema: un estudio de caso en la Selva Lacandona, Chiapas, México. Boletín de la Sociedad Botánica de México 81:65–80. [ Links ]
Puebla–Olivares F., Rodríguez–Ayala E., Hernández–Baños B.E. and Navarro–S. A.G. 2002. Status and conservation of the avifauna of the Yaxchilán Natural Monument, Chiapas, México. Ornitologia Neotropical 13:381–396. [ Links ]
Riba R. and Pérez–García B. 1997. Pteridofitas. In: González–Soriano E., Dirzo R. and Vogt R.C. Eds. Historia Natural de los Tuxtlas, pp. 175–181, Universidad Nacional Autónoma de México and Conabio, Mexico City. [ Links ]
Richards P.W. 1996. The Tropical Rain Forest: an Ecological Study. 2nd ed., Cambridge University Press, Cambridge. [ Links ]
Rzedowski J. 1978. Vegetación de México. Limusa, Mexico City. [ Links ]
Solórzano S., Ibarra–Manríquez G. and Oyama K. 2002 Liana diversity and reproductive attributes in two tropical forests in Mexico. Biodiversity and Conservation 11:197–212. [ Links ]
SPP [Secretaría de Programación y Presupuesto]. 1981. Atlas Nacional del Medio Físico. Dirección General de Geografía del Territorio Nacional, Mexico City. [ Links ]
SEDESOL [Secretaría de Desarrollo Social]. 1992. Decreto por el que se declara área natural protegida con el carácter de Monumento Natural, a la zona conocida con el nombre de Yaxchilán con una superficie de 2,261–25–23 hectáreas, ubicada en el Municipio de Ocosingo, Chis. Diario Oficial de la Federación, 24 de agosto de 1992. [ Links ]
SEMARNAT [Secretaría del Medio Ambiente y Recursos Naturales]. 2002. Norma Oficial Mexicana NOM–059–ECOL–2001, Protección ambiental–Especies nativas de México de flora y fauna silvestres–Categorías de riesgo y especificaciones para su inclusión, exclusión o cambio–Lista de especies en riesgo. Diario Oficial de la Federación, 2a Sección, 6 de marzo de 2002. [ Links ]
SEMARNAT. 2003. La deforestación en 24 regiones Proders. Secretaría del Medio Ambiente y Recursos Naturales, Mexico City, 239 p. [ Links ]
Siebe C., Martínez–Ramos M., Segura–Warnholtz J., Rodríguez–Velásquez J. and Sánchez–Beltrán S. 1996. Soil and vegetation patterns in the tropical rain forest at Chajul, southeast México. In: Simorangkir D. Ed. Proceedings of the International Congress on Soils of Tropical Forest Ecosystems, pp. 40–58, 3rd Conference on Forest Soils, Mulawarman University Press, Samarida. [ Links ]
Tropicos.org. Missouri Botanical Garden. <www.tropicos.org> Accessed in 2008. [ Links ]
Valle–Doménech J.A. 2000. Análisis de la estructura de una hectárea de selva alta perennifolia en el Monumento Natural Yaxchilán (Chiapas), México. B.Sc. Thesis (Biology). Facultad de Ciencias, Universidad Nacional Autónoma de México, Mexico City, 99 p. [ Links ]
Villaseñor J.L. 2001. Catálogo de Autores de Plantas Vasculares de México. Instituto de Biología, Universidad Nacional Autónoma de México, Mexico City. [ Links ]
Walsh R.PD. 1996. Climate. In: Richards P.W. The Tropical Rain Forest: an Ecological Study. 2nd ed., pp. 159–236, Cambridge University Press, Cambridge. [ Links ]
Wendt T. 1993. Composition, floristic affinities, and origins of the canopy tree flora of the Mexican Atlantic slope rain forests. In: Ramamoorthy T.P., Bye R., Lot A., and Fa J. Eds. Biological Diversity of Mexico. Origins and Distribution, pp. 595–680, Oxford University Press, New York. [ Links ]
Whitmore T.C. 1990. An Introduction to Tropical Rain Forests. Clarendon Press, Oxford. [ Links ]

