










Services on Demand
Journal

Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
 Cited by SciELO
Cited by SciELO Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkVeterinaria México
Print version ISSN 0301-5092
Vet. Méx vol.42 n.1 Ciudad de México Jan./Mar. 2011
Artículos científicos
Comportamiento maternal alrededor del parto y reconocimiento madre–cría en ovinos Pelibuey
Maternal behaviour around birth and mother–young recognition in Pelibuey sheep
Miriam Gloria Ramírez Martínez* Rosalba Soto González* Pascal Poindron Massot** Lorenzo Álvarez Ramírez*** Javier de Jesús Valencia Méndez‡ Francisco Rodolfo González Díaz* Angélica María Terrazas García*
* Laboratorio de Reproducción y Comportamiento Animal, Secretaría de Posgrado e Investigación, Facultad de Estudios Superiores Cuautitlán, Universidad Nacional Autónoma de México, Unidad de Investigación Multidisciplinaria, Laboratorio 02, km 2.5 Carretera Cuautitlán–Teoloyucan, San Sebastián Xhala, 54714, Cuautitlán Izcalli, Estado de México, México, Teléfono: (55) 56 23 19 99, extensión 39412.
** Laboratoire de Comportement, Neurobiologie et Adaptation, UMR 6175 INRA/CNRS/Université de Tours, Haras Nationaux, 37380 Nouzilly, France.
*** Centro de Enseñanza, Investigación y Extensión en Producción Animal en Altiplano, Facultad de Medicina Veterinaria y Zootecnia, Universidad Nacional Autónoma de México, 04510, México, D. F.
‡ Departamento de Reproducción, Facultad de Medicina Veterinaria y Zootecnia, Universidad Nacional Autónoma de México, 04510, México, D. F.
Correspondencia:
Dra. Angélica María Terrazas García,
Teléfono: (55) 56 23 19 99, extensión 39412,
Correo electrónico: garciate@servidor.unam.mx
Recibido el 25 de enero de 2010
Aceptado el 20 de septiembre de 2010.
Abstract
Maternal and social behavior in multiparous Pelibuey ewes was assessed under intensive production system. The social behavior was recorded several days before and during birth. Mother–young behavior during the first hour after birth was recorded. A selective maternal test was performed at 2 hours postpartum. Eight hours after birth, a double choice test was done in mothers to asses their ability to discriminate between the own or alien lamb. While a similar double choice test was performed in 12 hours–old lambs. The agitation index when the ewes were tested without companions was lower in parturient ewes than in145 days pregnant ewes and non pregnant ewes [parturient ewes: (—0.59 ± 0.9), 145 days pregnant ewes: (2.6 ± 0.7) and non pregnant ewes (4.2 ± 0.9) P < 0.05]. During the first hour after birth the latency to stand up, to begin to look for the udder and to suck was shorter in male than female lambs, also the male lambs sucked for longer time than females (P < 0.05). During this time mothers emitted more high–pitched bleats than low–pitched bleats (P < 0.05). Besides, the ewes got shorter latencies to lick the area of head and thorax of the lamb than its tail, but they licked more frequently the lamb's tail and thorax (P < 0.03). Two hours postpartum all mothers were selective, they showed clear preference to accept to the udder their own lamb and reject actively the alien ones (P < 0.05). At eight or 12 hours, mothers and lambs spent longer time watching and staying with their own familiar animal than with the alien ones (P < 0.05). It is concluded that Pelibuey ewes show an adequate maternal temperament, and the behavior of the newborn lamb could be affected because of its sex.
Key words: maternal behavior, indoor conditions, pelibuey breed, lambs.
Resumen
Se evaluó el comportamiento materno en ovejas multíparas de raza Pelibuey en estabulación, así como la conducta social, antes y durante el parto. Se evaluó el comportamiento madre–cría durante la primera hora posparto y se realizó una prueba de selectividad a dos horas posparto. A las ocho horas se realizó una prueba de elección entre su cordero y uno ajeno. A las 12 h, a los corderos se les hizo una prueba de elección entre la madre propia y una ajena. El índice de agitación sin compañeras fue menor al parto que en ovejas a 145 días de gestación y en ovejas vacías [al parto: (—0.59 ± 0.9), con 145 días (2.6 ± 0.7) y a vacías (4.2 ± 0.9) P < 0.05]. En la primera hora posparto, los corderos machos se incorporaron, iniciaron la búsqueda de la ubre y se amantaron más rápido y por más tiempo que las hembras (P < 0.05). Las madres emitieron más balidos bajos que altos (P < 0.05); limpiaron más rápido a la cría en la cabeza y el tórax que en la cola (P < 0.05), pero limpiaron más veces la cola que el tórax (P < 0.03). A dos horas posparto, las madres fueron selectivas y rechazaron a la cría ajena (P < 0.05). A las ocho y 12 horas, tanto madres como corderos se miraron y permanecieron más tiempo con su co–familiar que con el ajeno (P < 0.05). Se concluyó que las ovejas de la raza Pelibuey muestran buena aptitud maternal, y que la conducta de la cría varía según su sexo.
Palabras clave: conducta materna, estabulación, pelibuey, corderos.
Introducción
La conducta materna es el proceso que resulta de la combinación de factores neuronales, humorales y sensoriales, cuyo fin lleva al individuo a nutrir y cuidar a su progenie mediante la expresión de diversos patrones conductuales dirigidos a incrementar la viabilidad de la cría.1 En mamíferos, la supervivencia de la progenie es una etapa crítica de la reproducción, ya que determina el éxito reproductivo de los padres;2,3 asimismo, la expresión adecuada de la conducta materna adquiere importancia porque el recién nacido depende totalmente de la madre para su alimentación.3–5 En las hembras de cualquier especie existen varios elementos conductuales que caracterizan la conducta materna, en el caso de la oveja, la reducción de la conducta gregaria y el aislamiento del rebaño antes del parto forman parte de este perfil.1,6,7 Este aislamiento del grupo facilita el reconocimiento y el rápido establecimiento del vínculo selectivo madre–cría, lo cual representa el elemento propio de la conducta materna en la oveja.8 La conducta de aislamiento varía entre razas9,10 y representa una alteración transitoria al carácter gregario de la especie, huyen en grupo, mantienen vigilancia colectiva ante situaciones de peligro y forman grupos cerrados, aunque conservan su espacio individual.11
Las especies precoces como los ungulados tienen crías bien desarrolladas desde el punto de vista motriz y sensorial, con alta autonomía térmica. Las crías de estas especies son capaces de levantarse rápido y seguir a su madre después de nacidas, lo cual está asociado con un proceso de reconocimiento rápido entre ambos a través de diferentes vías sensoriales.12 En el ovino, la madre aprende a reconocer a la cría por su olor dentro de las primeras dos a cuatro horas posteriores al nacimiento,13,14 lo que resulta en el cuidado y amamantamiento exclusivo de su cría. Una vez establecido este vínculo selectivo la madre rechaza, e incluso muestra conductas agresivas, hacia cualquier cría ajena que intente amamantarse.8,15 La interacción madre–cría favorece el aprendizaje del olor del cordero.8,16 En la oveja, el reconocimiento a distancia intervienen el oído y la vista, aunque la participación de estos sentidos parece necesitar más tiempo.13,17,18 Los corderos también son capaces de mostrar preferencias hacia su madre desde las 12 horas de edad, aun cuando en algunos momentos intenten amamantarse de otras ovejas.19 Dicha preferencia depende de la capacidad de discriminación por parte de la cría entre un conjunto de señales de aceptación (mostradas por la madre propia) y señales de rechazo (mostradas por la madre ajena).20 La capacidad de reconocimiento del cordero se considera completa a las 48 horas después del nacimiento, cuando ya cuentan con la habilidad de discriminación de los rasgos individuales de su madre desde una distancia de varios metros.21
En México existen dos razas de pelo predominantes, la Pelibuey y la Black Belly, ambas fueron traídas desde el oeste del continente africano por los españoles durante la Colonia. Sin embargo, en un estudio reciente se determinó que las razas mexicanas (criollas, borrego Chiapas y Pelibuey) presentan genotipo B, que los ubica con un origen europeo, mientras que la Black Belly parece ubicarse en genotipo A.22 Asimismo, por su distribución en el mundo, la raza Pelibuey es considerada como una raza local, mientras que la Black Belly es más de tipo transfronterizo.23 La FAO ha informado que las razas locales cuentan con características superiores de adaptación al medio, y que sus recursos zoogenéticos contribuyen a la biodiversidad ganadera. Además, tienen un gran valor socioeconómico porque ayudan a disminuir la pobreza y garantizar la sustentabilidad ambiental.24
En los últimos años se ha incrementado el interés de los ovinocultores nacionales por la cría y producción de la raza Pelibuey, en forma pura o en cruzamientos principalmente con las razas sintéticas Dorper y Kathadin. Entre las características potencialmente más valiosas de la raza Pelibuey se encuentran las genéticas y productivas, como su alta prolificidad, baja estacionalidad, precocidad y rusticidad,23,25 por lo que en México la crianza sistemática de esta raza representa una alternativa de producción por su buena adaptación al medio ambiente.23–25
En México no existe un programa de selección de razas consistente, sobre todo de aquellas que pueden mostrar un mejor temperamento materno. El hecho de conocer razas que cuenten con estas características podrá servir para utilizarlas como criadoras de animales terminales que se van al consumidor. Conocer las características conductuales relacionadas con la maternidad en razas que tienen un alto potencial de adaptación serviría para, en un futuro, sugerir el uso de las mismas en la producción ovina nacional.
El presente trabajo tuvo como objetivo evaluar la aptitud materna de las ovejas Pelibuey, midiendo algunos elementos de la conducta materna y de la cría antes y después del parto, que han sido bien descritos en otras razas con buena aptitud maternal como la Scottish Blackface.26
Material y métodos
Ética
El diseño y protocolo experimental descrito en este trabajo fue aprobado por el subcomité local para el cuidado y uso de los animales para experimentación.
Animales
El presente trabajo se realizó con dos rebaños de ovejas de la raza Pelibuey, mantenidos en condiciones similares de manejo. La primera parte experimental (respuesta a la separación social) se realizó en la Facultad de Estudios Superiores Cuautitlán, de la Universidad Nacional Autónoma de México (UNAM), con un grupo de 27 ovejas multíparas. La segunda parte (relación madre–cría al parto) se realizó con un rebaño de 21 ovejas multíparas de la misma raza, pertenecientes al Centro de Enseñanza, Práctica e Investigación en Producción y Salud Animal, Facultad de Medicina Veterinaria y Zootecnia de la UNAM, localizado en Topilejo, Ciudad de México.
En ambos casos los animales permanecieron en estabulación total y fueron alimentados de acuerdo con sus requerimientos,27 con heno de alfalfa y avena, silo de maíz, alimento concentrado y agua a libre acceso.
La época de gestación se controló de manera completa. Se aplicaron por 14 días esponjas vaginales impregnadas con 30 mg de acetato de fuorogestona* y 350 UI de gonadotropina coriónica equina (eCG)* al retirar la esponja. El apareamiento fue controlado en todas las hembras para poder estimar la fecha prevista de parto (150 días de gestación). La sincronización de la reproducción fue programada de tal manera que se pudieran probar en forma simultánea, una oveja de cada grupo, para la prueba de conducta social.
Proceso experimental
Evaluación de la disminución de la conducta social alrededor del parto
Para esta fase experimental se utilizaron tres grupos independientes de hembras, según uno de los tres estados fisiológicos siguientes:
• Grupo testigo de hembras vacías (no gestantes ni lactando, n = 9).
• Grupo de hembras a 145 días de gestación (77.4 ± 5.4 horas antes del parto, n = 11).
• Grupo de hembras al parto (44.3 ± 0.01 minutos antes del parto, n = 7).
A todos los animales se les realizó una prueba de separación social, similar a la descrita por Poindron et al.6
Se colocó a cada oveja en un corral de 2 x 2 m, dicho corral se encontraba, a su vez, dentro de otro de 4 x 6 m en el que se introducían otras ovejas. La hembra fue probada durante cinco minutos en presencia de 10 ovejas vacías de la misma raza (pertenecientes al mismo rebaño) y otros 5 minutos fue probada en aislamiento total, retirando a las 10 ovejas a otro corral fuera de la vista del sujeto estudiado. Ambas fases de la prueba ocurrieron de manera continua, siempre en el mismo orden.
Con la ayuda de cronómetros y hojas formateadas, se registraron las siguientes conductas:
Frecuencia de balidos altos: Número de vocalizaciones emitidas por el animal con el hocico abierto.
Frecuencia de balidos bajos: Número de vocalizaciones emitidas con el hocico cerrado.
Frecuencia de cambios de lugar: Dentro del corral de prueba se trazaron en el suelo dos líneas que lo dividieron internamente en cuatro cuadrantes, de un metro cuadrado cada uno. Se contabilizó el número de movimientos realizados de un cuadrante a cualquiera de los otros tres, considerando que la hembra cruzó cada cuadrante con la cabeza y los miembros delanteros.
Frecuencia de olfateos a objetos: Número de veces que la hembra olfateó objetos (acercaba su nariz con intención de oler a menos de cinco cm de distancia, ya sea al suelo, a paneles o material que hubiera en el corral de prueba).
Frecuencia de olfateos a conespecíficos: Número de veces que acercaba su nariz para oler a sus compañeras.
Frecuencia de intentos de escape del corral: Número de veces que la hembra intentó saltar, o logró escapar fuera del corral de prueba.
Frecuencia de eliminaciones: Número de veces que la hembra orinó o defecó durante la prueba.
También se calculó un índice de agitación en cada una de las partes de la prueba, el índice se calculó sumando las variables estandarizadas de balidos altos + cambios de lugar + eliminaciones + intentos de escape. La estandarización, con media de cero y desviación estándar de ±1 permitió que todas las variables tuvieran el mismo peso en el índice. Las variables que no pudieron entrar en el índice se analizaron de manera individual para explorar un efecto de la situación social del animal.
Evaluación de la conducta de la madre y la cría durante la primera hora posparto
Condiciones de observación y variables conductuales
Se registró la información de un total de 17 ovejas, de las cuales hubo 12 partos dobles (17 machos y siete hembras) y cinco sencillos (cinco machos y cero hembras).
Cuando en la madre asomaban los miembros anteriores del producto a través de la vulva, o en su caso alguna parte del cuerpo del cordero, era colocada en un corral de 2 x 2 m con paneles abiertos, este corral fue colocado en el lugar elegido por la madre para el parto. Una vez que la oveja había expulsado la primera cría, iniciaba una filmación, que se prolongaba hasta una hora después de haber tenido a la primera cría. Se registraron las conductas de todas las crías nacidas durante la primera hora. La oveja y su(s) cría(s) permanecían ahí hasta cumplir las dos horas posparto. Al finalizar la filmación, las crías eran identificadas con aretes.
Las conductas registradas en las madres durante la filmación fueron:
Latencia y frecuencia de olfateos a la cría: La primera vez que la madre olfateó o acercó su nariz hacia el cordero a una distancia menor de cinco cm, con la intención de olerlo, y se contabilizó el número de veces que lo hizo en la hora de aceptación.
Frecuencia de balidos altos y bajos: El número de veces que la hembra emitió vocalizaciones con el hocico abierto (balidos altos) o cerrado (balidos bajos).
Latencia, frecuencia y duración de limpieza de la cría: Tiempo de inicio, número de veces y duración de la limpieza. La limpieza o acicalamiento se definió como el momento en que la madre frota alguna parte del cuerpo del cordero con la lengua. Se dividió el cuerpo del cordero en secciones para identificar la preferencia de limpieza, sólo se pudo medir la latencia y la frecuencia de esa conducta.
Frecuencia de agresiones: Número de veces que la hembra golpeaba o amenazaba a la cría, especialmente con la cabeza.
Las conductas evaluadas en las crías fueron:
Latencia de intento por levantarse: Tiempo en que por primera vez el cordero intentó levantarse.
Latencia de ponerse en pie: Tiempo en que la cría se puso de pie por primera vez.
Latencia de búsqueda de la ubre: Tiempo en que el cordero inició por vez primera la búsqueda de la ubre.
Latencia, frecuencia y duración cerca de la ubre: Cuando el cordero estuvo cerca de la región inguinal de su madre por primera vez, el número de veces que estuvo cerca de esa zona y la duración total que permaneció ahí.
Latencia, frecuencia y duración del amamantamiento: Cuando el cordero succionó por primera vez uno de los pezones de la madre, el número de veces que lo hizo durante la hora de observación, y la duración total de succión de los pezones de la madre.
La información registrada durante este periodo se analizó con la ayuda de un sistema para análisis de comportamientos.**
Selectividad a las dos horas posparto
Descripción de la prueba
La selectividad maternal en la oveja se define como la capacidad de la madre para aceptar únicamente a su cordero en la ubre, y rechazar a cualquier cordero ajeno que intente amamantarse de ella. Dicha capacidad está controlada por el reconocimiento de señales olfatorias de la cría.
Esta prueba se aplicó a las dos horas posparto en el mismo corral de 2 m x 2 m en que la oveja parió, y consistió en medir la conducta de la madre (n = 19) hacia su cordero y hacia una cría ajena, durante cinco minutos con cada una. El registro de las conductas se realizó con la ayuda de dos cronómetros y hojas formateadas.
Conductas registradas
Frecuencia de emisión de balidos bajos de la oveja.
Frecuencia de emisión de balidos altos de la oveja.
Frecuencia de aceptaciones en la ubre: Número de veces que la madre permitió a la cría el acercamiento a la ubre por un periodo mayor a cinco segundos continuos.
Duración de amamantamiento: Tiempo total que la madre permitió el amamantamiento.
Frecuencia de rechazos a la ubre: Número de veces que la madre evitó que el cordero se acercara a la ubre.
Frecuencia de agresiones: Número de veces que la madre dirigió golpes, amenazas o mordidas al cordero durante la prueba.
Reconocimiento no olfatorio a las ocho horas posparto
A las ocho horas posparto, a 17 ovejas se les realizó prueba de reconocimiento a distancia de doble elección sin la ayuda de señales olfatorias, similar a la descrita por Terrazas et al.17 La prueba se realizó en un corral armable en forma de triángulo con dimensión de 10 x 10 x 10 m (Figura 1), construido con paneles cerrados de un metro de altura para impedir distractores a los animales. En las esquinas de la base del triángulo se armaron dos corrales de 1 m2 con paneles abiertos, en los cuales se colocaron los corderos (propio y extraño).

A un metro de distancia alrededor de estos corrales se colocó una barrera armada con paneles abiertos para impedir la percepción de señales olfatorias de los corderos hacia la madre;28 sin embargo, esta barrera permitía un contacto visual y auditivo entre la madre y las crías. Una zona de un metro alrededor de esa barrera fue definida como zona de contacto (Figura 1). En la tercera esquina del corral se armó otro corral de 2 x 1 m (corral de espera) en donde se colocó a la madre antes de iniciar la prueba. Esta prueba duró cinco minutos.
Al momento de la prueba, se introdujo a la oveja en el corral de espera, en seguida uno de sus corderos y uno extraño de edades similares fueron colocados individualmente en los corrales a contraesquina del corral de prueba.
La posición del cordero propio (derecha o izquierda) se cambiaba en cada prueba. En el caso de las ovejas con mellizos, el cordero que no era probado en ese momento era apartado del sitio de prueba a una distancia no menor de 10 m y fuera de la vista de la madre para evitar distracciones. Una vez liberada la oveja del corral de espera, se inició el registro de las conductas con la ayuda de dos observadores, cronómetros, un contador y hojas formateadas.
Las conductas registradas fueron:
Duración de permanencia cerca de cada cordero (en segundos): Tiempo durante el cual la madre permaneció en la zona de contacto cerca de cada cordero.
Tiempo de observación a cada cría (en segundos): Tiempo total en que la madre miró hacia la dirección en que se encontraba cada uno de los corderos, a cualquier posición de la oveja en el corral triangular. Frecuencia de balidos altos: Número de veces que la madre emitió balidos altos.
Latericia de salida del corral de espera (en segundos): Tiempo que tardó la madre en salir del corral de espera una vez que se abrió la puerta a éste. Latericia de alcance de la primera cría (en segundos): Tiempo que tardó la madre en llegar a cualquiera de las zonas de contacto de los corderos por primera vez.
Frecuencia de visitas a cada cría: Número de veces que la madre visitó a cada una de las zonas de contacto de los corderos, sin importar el tiempo que permaneció en esa zona.
Evaluación de la capacidad del cordero para reconocer a su madre a las 12 horas de edad
Con el objetivo de investigar la capacidad de los corderos para diferenciar a su madre de una extraña a las 12 horas de nacidos, los corderos fueron sometidos a una prueba de elección doble para evaluar la preferencia por su madre, similar a la descrita por Terrazas et al,20 para esta etapa se utilizaron todos los corderos nacidos del grupo de ovejas de la segunda parte experimental (n = 32: 21 machos y 11 hembras, ocho de parto sencillo y 24 de parto doble). Desde el nacimiento las crías permanecieron sólo con su madre, previa identificación y manejo sanitario general, a las cuatro horas de edad fueron liberados junto con las otras ovejas para que pudieran tener interacción con otras madres y corderos.
Se utilizó un corral triangular con paneles cerrados, de 6 m x 6 m (Figura 2). En la base se colocaron dos corrales de 1 m x 2 m, uno para la madre ajena y el otro para la madre propia; estos corrales se armaron con paneles abiertos con la finalidad de que existiera contacto visual y olfatorio entre el cordero y las madres. En la parte opuesta se armó un corral de 1 m x 1 m para colocar al cordero de prueba.

Al momento de realizar la prueba se llevó a las dos madres y sus crías al corral de prueba, cada madre fue colocada en su respectivo corral (dispuestos en la base del triángulo) mientras tanto el cordero se colocó en el corral de espera y el cordero de la madre ajena permaneció lejos de la vista de su madre para evitar distracción. A diferencia de las madres, los corderos podían olfatear a las ovejas además de verlas y oírlas, pero no podían amamantarse.
La prueba duró cinco minutos, y las conductas se registraron en hojas formateadas iguales a las empleadas con las madres.
Conductas registradas:
Latencia de salida.
Latencia de alcance de la primera madre elegida.
Duración de permanencia con cada una de las madres.
Duración de observación a cada una de las madres.
Frecuencia de balidos altos.
Análisis estadístico
A todos los datos se les hizo una comprobación de su distribución, para ello se utilizó una prueba de Kolmogorov–Smirnov, la probabilidad de Lilliefors arrojó que la mayoría de los datos eran significativos (P ≤ 0.05), lo que indica que no contaban con una distribución normal, por tanto, para analizarlos se utilizaron pruebas de estadística no paramétrica.29
De esta manera, para evaluar la respuesta a la separación social, así como para comparar entre los tres grupos probados (vacías, gestantes y parturientas), se utilizaron las pruebas de Kruskal Wallis y U de Mann Whitney. Para comparar la conducta con y sin compañeras dentro de cada grupo, se utilizó la prueba de Wilcoxon de rangos.
En la observación de la conducta al parto, tanto en la madre como en la cría se realizó análisis descriptivo. Además, se investigó si existían preferencias por parte de la madre en lamer alguna zona en particular del cuerpo de la cría, se consideraron tres regiones en el cuerpo del cordero: tórax, cabeza y cola, y los datos se compararon con pruebas Friedman y de Wilcoxon. Las vocalizaciones de los corderos fueron analizadas con una prueba de Wilcoxon de rangos. La proporción de corderos machos vs. hembras, se comparó utilizando una prueba de Ji cuadrada de Pearson. Para comparar las conductas entre corderos machos y hembras se utilizó una prueba U de Mann Whitney.
Los datos se presentan en términos de medianas y rangos intercuartiles. La información fue procesada con la ayuda del programa estadístico SYSTAT 10.
Resultados
Disminución de la conducta social alrededor del parto
Prueba con compañeras
Durante esta etapa de la prueba, la frecuencia de emisión de balidos altos difirió entre grupos (P = 0.05), con una tendencia de las ovejas del grupo al parto a emitir un mayor número de balidos altos que las ovejas gestantes (P = 0.06, Cuadro 1). No se encontraron diferencias significativas entre los demás grupos.
La frecuencia de balidos bajos también difirió entre grupos (P < 0.001). El grupo de las ovejas parturientas emitió mayor número de balidos bajos que las ovejas de los grupos vacías y gestantes (P < 0.005, Cuadro 1), estos dos últimos grupos no difirieron entre sí.
El olfateo a conespecíficos difirió entre los tres estados fisiológicos (P = 0.02); las ovejas vacías tuvieron una frecuencia de olfateos a otras ovejas mayor que las ovejas parturientas (P = 0.02, Cuadro 1).
Prueba sin compañeras
Las ovejas parturientas emitieron más balidos bajos que las ovejas gestantes y vacías (P < 0.04, Cuadro 1). No se encontraron diferencias entre las gestantes y las vacías (P > 0.05).
La frecuencia de cambios de lugar difirió entre grupos (P = 0.007). Las ovejas parturientas cambiaron de lugar con menor frecuencia que las vacías y gestantes (P < 0.01, Cuadro 1). No se encontraron diferencias entre las ovejas del grupo gestantes y las del grupo vacías (P > 0.05). Éstas tuvieron mayores intentos de escape que las ovejas gestantes y parturientas (P < 0.01, Cuadro 1); no se registraron diferencias entre las gestantes y las parturientas (P > 0.05).
El índice de agitación con compañeras no fue afectado por el grupo (P > 0.05); sin embargo, cuando se evaluó el índice sin compañeras se encontró que la respuesta de los grupos fue diferente (P = 0.008, Figura 3).

De esta manera, se encontró que las ovejas con 145 días de gestación tuvieron mayor índice de agitación, cuando estaban sin compañeras, que las ovejas parturientas (P = 0.02, Figura 3); asimismo, el índice de agitación sin compañeras fue notablemente más alto en las ovejas vacías que en las parturientas (P = 0.004, Figura 3). Mientras que las ovejas probadas a 145 días de gestación no difirieron con las ovejas vacías sin compañeras (P > 0.05).
Comparación con compañeras vs sin compañeras
Las ovejas de los tres grupos emitieron mayor frecuencia de balidos altos y presentaron cambios de lugar cuando fueron aisladas de sus conespecíficos (P < 0.02, Cuadro 1). Las frecuencias de eliminaciones e intentos de escape del corral fueron mayores, al encontrarse sin compañeras, en las ovejas gestantes y vacías (P < 0.02, Cuadro 1). Ambas conductas no difirieron entre las dos partes de la prueba en el grupo de ovejas parturientas (P > 0.05).
Sólo las ovejas parturientas emitieron balidos bajos, pero la frecuencia de emisión no difirió entre las dos partes de la prueba (P > 0.05, Cuadro 1).
Finalmente, el índice de agitación para los tres grupos fue mayor en la parte de la prueba sin compañeras (P < 0.02, Figura 3).
Conducta de la madre y la cría durante la primera hora posparto
Conducta de la madre
Se encontró que la latencia de lamer en los corderos dependía de la zona del cuerpo de la cría (P = 0.014, Figura 4). Las madres iniciaron más rápido el lamido o la limpieza de las crías en la región de la cabeza y del tórax que en la región de la cola (P ≤ 0.01). No se encontraron diferencias entre la latencia de lamer la región de la cabeza y la del tórax (P > 0.05).

La frecuencia de lamido a las distintas regiones del cuerpo sólo difirió entre el lamido de la región de la cola y la del tórax, la madres limpiaron con mayor frecuencia la región de la cola que la del tórax (P = 0.03, Cuadro 2). Respecto de la conducta vocal de las madres, ellas emitieron más balidos bajos que balidos altos (P = 0.006, Cuadro 2).
Conducta de la cría
La proporción de corderos nacidos macho fue mayor que la de hembras (22/29 vs 7/29, respectivamente P < 0.001). La latencia para ponerse de pie después del nacimiento, de buscar la ubre, de llegar cerca de la ubre, así como la latencia del amamantamiento, fueron significativamente menores para los corderos macho que para las hembras (P < 0.05, Cuadro 1). Los machos tuvieron mayor frecuencia y duración para iniciar el amamantamiento que las hembras (P < 0.05). De hecho, la tendencia indicó que los machos tenían mayor frecuencia y tiempo de amamantamiento que las hembras (P = 0.06, Cuadro 3).
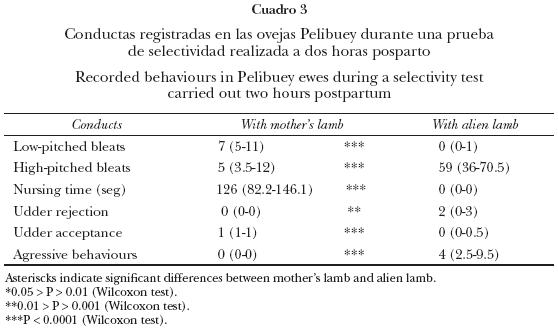
Selectividad materna a las dos horas posparto
La frecuencia de balidos bajos fue mayor durante la prueba con la cría propia que con la ajena (P < 0.001, Cuadro 4), mientras que la frecuencia de balidos altos fue mayor cuando se probó con la cría ajena que con la cría propia (P < 0.05). Asimismo, el tiempo de amamantamiento y la frecuencia de aceptaciones de la ubre fueron mayores para la cría propia que para la ajena (P < 0.0001). La frecuencia de rechazos y conductas agresivas fueron mayores para la cría ajena que para la propia (P ≤ 0.005; Cuadro 4).
Reconocimiento no olfatorio a las ocho horas posparto
La madre tuvo una latencia de salida del corral de espera con una mediana de tres segundos, mientras que la latencia de alcance a las crías fue de nueve segundos. Las madres permanecieron más tiempo cerca de la cría propia que de la ajena [126 (80–188) vs 4 (0–21) seg, P = 0.001]. También miraron por más tiempo a su cría que a la ajena [73.8 (53–91.2) vs 34 (22–40) seg, P = 0.001]. Las madres realizaron más visitas a la cría propia que a la ajena [6 (4–6) vs 1 (0–2), P = 0.001].
Capacidad del cordero para reconocer a su madre a las 12 horas de edad
En esta prueba también se midió el efecto del sexo del cordero y el tipo de parto del que provenía, y no se encontraron diferencias significativas en todas las conductas registradas entre hembras y machos, ni entre partos dobles y sencillos (P < 0.05). El cordero tuvo latencia de salida del corral de espera con mediana de 17.5 seg, mientras que la latencia de alcance en las zonas de contacto con las madres fue de 44 seg. Los corderos permanecieron más tiempo cerca de la madre propia que de la madre ajena [109 (30–200) vs 49.5 (7.5–109) seg, P = 0.015]. Asimismo, miraron más tiempo hacia la madre propia que a la ajena [61.4 (30.5–85.2) vs 31 (13.5–69) seg, P = 0.03]. La frecuencia de visitas a la madre propia tendió a ser mayor que la de la madre ajena [2 (1–3) vs 1 (0.5–3), P = 0.07]. La frecuencia media de balidos emitidos por los corderos durante la prueba fue de 23 (13.5–38).
Discusión
El presente trabajo generó información acerca del comportamiento materno y el reconocimiento mutuo, por primera vez realizado en ovejas y sus crías de la raza Pelibuey, en condiciones estabuladas.
Se observó que la condición fisiológica de la oveja influyó en el cambio de su conducta social, especialmente relacionada con el gregarismo, pues las ovejas vacías respondieron muy fuertemente a la separación social, mientras que la respuesta disminuyó al momento del parto. Estos estudios son similares a los registrados en ovejas de la raza Rambouillet, en las que se encontró que, a medida que se acercaba el parto, la respuesta a la separación social también disminuía.6 La disminución de la conducta gregaria también se ha encontrado en otras especies de rumiantes, en particular en cabras bajo condiciones intensivas. Veinticuatro horas antes del parto, las hembras se agitaron menos en una prueba de separación social similar a la utilizada en este experimento.30
En este estudio no se midió la tendencia de las hembras Pelibuey a buscar aislamiento, porque el trabajo se hizo en condiciones estabuladas. Sin embargo, podría ser tema de estudio, ya que muchos de los rebaños de esta raza son criados bajo condiciones extensivas,31,32 donde incluso el animal podría buscar sitios apropiados para el aislamiento, como sucede en las ovejas Merino en pastoreo.33
Durante la prueba con compañeras, las ovejas parturientas emitieron balidos altos y bajos con mayor frecuencia que las ovejas vacías y con 145 días de gestación. Mientras que en la prueba sin compañeras la emisión de balidos bajos también fue alta, resultado similar a las observaciones hechas por Sèbe et al.,34 quienes registraron una alta actividad vocal durante las primeras horas posparto en ovejas Ile de France. Los presentes resultados en la oveja Pelibuey demuestran una clara disminución de su actividad social mientras se encuentran en el parto, asociada con la necesidad de atención a la camada, lo cual es similar a lo registrado en razas cuya conducta maternal es notoria, como la Scottish Blackface.35,36
Las madres comenzaron la limpieza de las crías durante la primera hora posparto, se ha corroborado que con ello se estimula en varias vías a las crías para incorporarse y amamantarse lo más rápido posible, tanto en ovejas como en cabras.37,38 El hecho de responder rápidamente, como lo hicieron las ovejas Pelibuey, es un buen indicio de motivación materna, similar a la que muestran ovejas de otras razas con buena habilidad materna, como la Scottish Blackface.26,35 En el presente trabajo la conducta que ocupó mayor tiempo durante la primera hora posparto fue la de limpieza de la cría que, como se ha informado, es necesaria para desencadenar la actividad asociada con el amamantamiento. Asimismo, la emisión de balidos bajos fue otra de las conductas que se presentó con mayor frecuencia. Los balidos de baja frecuencia son eventos conductuales que en ovejas se presentan exclusivamente durante los periodos maternales y tienen la función principal de estimular a la cría para incorporarse al menos durante la primera hora de nacidos.21,34,39
La latencia de amamantamiento desde la expulsión de la cría fue relativamente corta, tal y como sucede en las razas de buen temperamento materno.35 Al respecto, la primera vez que la cría se amamantó fue de alrededor de 30 minutos, y todos los corderos nacidos en este estudio se amamantaron dentro de la primera hora posparto. Es de vital importancia que los corderos se amamanten lo más pronto posible, ya que ello incrementa sus posibilidades de sobrevivencia.40 Los resultados del comportamiento materno de la oveja Pelibuey durante la primera hora posparto son similares a lo previamente informado en ovejas26,35,41–43 y en cabras.41,44
En este estudio, en la primera hora posparto resultó relevante que las madres lamieran más rápidamente la región de la cabeza y el tórax, y que limpiaran con mayor frecuencia la zona de la cabeza y la de la cola, en comparación con el tórax. Como otros estudios lo sugieren, la comunicación táctil representa un factor importante en el afianzamiento del vínculo entre la madre y su cría y es el estímulo principal para que el cordero inicie conductas propias al momento del nacimiento.37–39 Los resultados del presente estudio indican que la madre tiene preferencia por limpiar más algunas zonas específicas del cordero, como ya lo sugerían otras investigaciones.45 Los factores que inducen dicha atracción no han sido completamente explicados; investigaciones realizadas en útero demuestran que el feto responde de manera temprana a estímulos en la cabeza, que desencadenan una serie de movimientos, los cuales se sincronizan hacia las extremidades.46
Sin embargo, la relación entre la preferencia para estimular con mayor frecuencia dichas regiones del cordero por parte de la madre, y el hecho de desencadenar en el cordero movimientos asociados con la incorporación después del nacimiento quedan aún por investigarse. Por ejemplo, en otros estudios se ha observado que cuando se altera el color de las zonas de la cabeza y la cola del cordero, se afecta también el reconocimiento a través de la vista por parte de la madre;47 por lo que es posible que existan ciertas señales en esas regiones que son atractivas para la madre y, por tanto, las prefiera más. Esta situación parecería similar a lo encontrado en roedores, que paren crías altriciales, en los que se ha observado que la madre prefiere también limpiar zonas como la cabeza y la región anogenital de las crías, incluso se ha demostrado que las madres pueden identificar a las crías en ausencia de otras señales sensoriales.48
Respecto del comportamiento de la cría durante la primera hora de nacida se encontró que el sexo también es determinante, ya que los corderos macho tuvieron mayor actividad que las hembras, por lo que se amamantaron más rápido. Resultados similares se han encontrado en cabras Criollas durante la primera hora de nacidas.49 Asimismo, en cabras primíparas se han encontrado tendencias de mayor actividad en crías macho que en crías hembra.50
En contraste, en trabajos realizados en ovejas Suffolk, los corderos macho tuvieron menor desempeño que las hembras durante las primeras horas de nacidos, mientras que no se observó efecto del sexo de la cría en ese mismo estudio en la raza Scottish Blackface; además, se informó que el orden de nacimiento en partos múltiples afecta el desempeño conductual de los corderos.51 Las inconsistencias de los pocos trabajos hasta ahora realizados en ovejas y cabras sobre el efecto del sexo de la cría sobre su desempeño conductual, indican la necesidad de realizar más estudios para aclarar dichos hallazgos.
En relación con la presencia del comportamiento selectivo evaluado en este estudio en ovejas Pelibuey a las dos horas posparto, se encontró que todas las ovejas probadas tuvieron una clara respuesta de rechazo a las crías ajenas y de aceptación a las crías propias. Este resultado es consistente con las observaciones informadas por primera vez por Smith,14 y confirmado por otros autores en ovejas,41 de tal manera que cuando las ovejas presentan adecuadamente esta respuesta selectiva hacia los corderos se considera un desarrollo maternal normal.
Las ovejas también fueron probadas en su capacidad para reconocer a su cría a distancia o en ausencia de las señales olfatorias a ocho horas posparto. En dicha prueba las madres fueron más atraídas por la cría propia que por la ajena, lo cual indica su capacidad de discriminación. Las ovejas Pelibuey son capaces de conocer sus crías a las ocho horas, por lo que se concluye que su desempeño parece tan adecuado como el que presentan otras razas.13,28
En el caso de los corderos Pelibuey, en el presente estudio, se observó que éstos ya son capaces de discriminar a su madre de una ajena a las 12 horas de edad, en este caso haciendo uso de todas las señales sensoriales provistas por la madre. Este resultado es similar al de los corderos de la cruza Merino X Border–Leicester, en contraste con los corderos Merino, que fueron más lentos para reconocer a sus madres.19 En otro estudio realizado en corderos de la raza Columbia se encontró que también a las 12 horas de edad ya son capaces de discriminar a su madre de una ajena a través de la percepción de señales sensoriales combinadas como la actitud calmada de la madre y la emisión de balidos bajos durante la prueba.20
Se concluyó que la oveja Pelibuey en condiciones estabuladas puede mostrar comportamiento pre y posparto adecuado que se compara con razas de aptitud maternal alta bien estudiadas, como la Scottish Blackface, lo que podría constituir una ventaja más para la producción de esta raza en México.
Agradecimientos
Los autores agradecen al Consejo Nacional de Ciencia y Tecnología por el financiamiento parcial de este trabajo con el proyecto clave: 4881–N y por la beca de maestría otorgada a Miriam Ramírez, así como el financiamiento otorgado por el Fondo Internacional para la Ciencia B3872–1UNAM–FIS y por el Programa de Apoyo a Proyectos de Investigación e Innovación Tecnológica–UNAM–IN207508.
Se extiende un cordial agradecimiento a Norma Serafín López por la asistencia en el análisis de videos con el programa Observer®, a Antonio Roldán por su ayuda en el manejo reproductivo de los animales, a Emmanuel Lara, Hitandewy Sánchez y Ángel Fierros por su ayuda durante el proceso experimental.
Referencias
1. GONZALEZ–MARISCAL G, POINDRON P. Parental care in mammals: inmediate internal and sensory factors of control. Horm Behav 2002; 1:215–298. [ Links ]
2. CARLSON SG, LARSSON K, SCHALLER J. Early mother–child contact and nursing. Reprod Nutr Develop 1980; 20:881–899. [ Links ]
3. POINDRON P. El control fisiológico de la conducta materna al momento del parto en ovinos y caprinos. En: VELÁZQUEZ MJ, editor. Biología de la reproducción II. México, D F: Universidad Autónoma Metropolitana–Programa Universitario de Investigación en Salud; 2001:301–323. [ Links ]
4. NOWAK R, PORTER RH, LEVY F, ORGEUR P, SCHAAL B. Role of mother–young interactions in the survival of offspring in domestic mammals. Rev Reprod 2000; 5:153–163. [ Links ]
5. NOWAK R, POINDRON P. From birth to colostrum: early steps leading to lamb survival. Reprod Nutr Dev 2006; 46. [ Links ]
6. POINDRON P, SOTO R, ROMEYER A. Decrease of response to social separation in preparturient ewes. Behav Processes 1997; 40:45–51. [ Links ]
7. POINDRON P, LÉVY F, KREHBIEL D. Genital, olfactory, and endocrine interactions in the development of maternal behaviour in the parturient ewe. Psychoneu–roendocrinology 1988; 13:99–125. [ Links ]
8. LÉVY F, KENDRICK K, KEVERNE EB, PORTER RH, ROMEYER A. Physiological, sensory and experiential factors of parental care in sheep. Adv Study Behav 1996; 25:385–473. [ Links ]
9. STEVENS D, ALEXANDER G, LYNCH JJ. Do Merino ewes seek isolation or sheltering at lambing? Appl Anim Ethology 1981; 7:149–166. [ Links ]
10. ALEXANDER G, STEVENS D, Bradley LR. Distribution of feld birth–sites of lambing ewes. Aust J Exp Agric 1990; 30:759–767. [ Links ]
11. DUMONT B, BOISSY A. Grazing behaviour of sheep in a situation of confict between feeding and social motivations. Behav Processes 2000; 49:131–138. [ Links ]
12. POINDRON P. Mechanisms of activation of maternal behaviour in mammals. Reprod Nutr Dev 2005; 45:341–351. [ Links ]
13. KELLER M, MEURISSE M, POINDRON P, NOWAK R, FERREIRA G, SHAYIT M et al. Maternal experience influences the establishment of visual/auditory, but not olfactory recognition of the newborn lamb by ewes at parturition. Dev Psychobiol 2003; 43:167–176. [ Links ]
14. SMITH F, VAN–TOLLER VC, BOYES T. The "critical period" in the attachment of lambs and ewes. Anim Behav 1966; 14:120–125. [ Links ]
15. POINDRON P, NOWAK R, LÉVY F, PORTER RH, SCHAAL B. Development of exclusive mother–young bonding in sheep and goats. Oxf Rev Reprod Biol 1993; 15:311–364. [ Links ]
16. POINDRON P, LE NEINDRE P. Endocrine and sensory regulation of maternal behavior in the ewe. Adv Study Behav 1980; 11:75–119. [ Links ]
17. TERRAZAS A, FERREIRA G, LÉVY F, NOWAK R, SERAFIN N, ORGEUR P et al. Do ewes recognize their lambs within the first day postpartum without the help of olfactory cues? Behav Processes 1999; 47:19–29. [ Links ]
18. FERREIRA G, TERRAZAS A, POINDRON P, NOWAK R, ORGEUR P, LÉVY F. Learning of olfactory cues is not necessary for early lamb recognition by the mother. Physiol Behav 2000; 69:405–412. [ Links ]
19. NOWAK R, POINDRON P, LE NEINDRE P, PUTU IG. Ability of 12–hour–old Merino and crossbred lambs to recognize their mothers. Appl Anim Behav Sci 1987; 17:263–271. [ Links ]
20. TERRAZAS A, NOWAK R, SERAFIN N, FERREIRA G, LÉVY F, POINDRON P. Twenty–Four–Hour–Old lambs rely more on maternal behavior than on the learning of individual characteristics to discriminate between their own and alien mother. Dev Psychobiol 2002; 40:408–418. [ Links ]
21. NOWAK R. Mother and sibling discrimination at a distance by three– to seven–day–old lambs. Dev Psychobiol 1990; 23:285–295. [ Links ]
22. ULLOA–ARVIZUA R, GAYOSSO–VÁZQUEZA A, ALONSO MORALESA J. Origen genético del ovino criollo mexicano (Ovis aries) por el análisis del gen del Citocromo C Oxidasa subunidad I. Téc Pecu 2009; 47:323–328. [ Links ]
23. GONZALEZ–REYNA A, VALENCIA J, FOOTE WC, MURPHY BD. Hair sheep in Mexico: reproduction in the Pelibuey sheep. Anim Breed Abstr 1991; 59:509–524. [ Links ]
24. FAO. Recursos zoogenéticos, la primera evaluación mundial. Roma: Comisión de los recursos genéticos para la alimentación y la agricultura, 2007. [ Links ]
25. PORRAS AA, ZARCO LB, VALENCIA JM. Estacionalidad reproductiva en ovejas. Cienc Vet 2003; 9:1–34. [ Links ]
26. DWYER CM, LAWRENCE AB. Ewe–ewe and ewe–lamb behaviour in a hill and a lowland breed of sheep: a study using embryo transfer. Appl Anim Behav Sci 1999; 61:319–334. [ Links ]
27. NRC. Nutrient Requirements of Small Ruminants, Sheep, Goats, Cervids, and New World Camelids. Washington, D C: The National Academies Press, 2007. [ Links ]
28. ALEXANDER G. Odour, and the recognition of lambs by Merino ewes. Appl Anim Ethology 1978; 4:153–158. [ Links ]
29. SIEGEL S, CASTELLAN NJ. Non parametric statistics for the behavioral sciences. 2nd ed.. Mexico DF: McGraw–Hill Interamericana, 1988. [ Links ]
30. POINDRON P, TERRAZAS A, NAVARRO–MONTES DE OCA ML, SERAFIN N, HERNANDEZ H. Sensory and physiological determinants of maternal behavior in the goat (Capra hircus). Horm Behav 2007; 52:99–105. [ Links ]
31. MASON IL. La raza Tabasco en México. Estudio de la Organización de las Naciones Unidas para la Agricultura y la Alimentación. FAO, 1980. [ Links ]
32. CRUZ LC. Generalidades de ovinos de pelo: Origen, distribución, razas, características. Experiencia en la producción de ovinos de pelo en el CEIEGT. México D F: Universidad Nacional Autónoma de México; 1995: 8–16. [ Links ]
33. LE'CRIVAIN E, JANEAU G. Comportement d'isolement et de recherche d'abri de brebis agnelant en plein air dans un syste' me d'e'levage a' caracte' re extensif. Biol Behav 1987; 12:127–148. [ Links ]
34. SÈBE FRN, POINDRON P, AUBIN T. Establishment of vocal communication and discrimination between ewes and their lamb in the first two days after parturition. Dev Psychobiol 2007; 49:375–386. [ Links ]
35. DWYER CM, LAWRENCE AB. Variability in the expression of maternal behaviour in primiparous sheep: Effects of genotype and litter size.. Appl Anim Behav Sci 1998; 58:311–330. [ Links ]
36. DWYER CM. Genetic and physiological determinants of maternal behavior and lamb survival. J Anim Sci 2008; 86:246–258. [ Links ]
37. COLLIAS NE. The analysis of socialization in sheep and goats. Ecology 1956; 37:228–239. [ Links ]
38. LICKLITER RE. Behavior associated with parturition in the domestic goat. Appl Anim Behav Sci 1985; 13:335–345. [ Links ]
39. VINCE MA. Newborn lambs and their dams: the interaction that leads to sucking. Adv Study Behav 1993; 22:239–268. [ Links ]
40. NOWAK R, POINDRON P. From birth to colostrum: early steps leading to lamb survival. Reprod Nutr Dev 2006; 46:431–446. [ Links ]
41. POINDRON P, LÉVY F, KELLER M. Maternal responsiveness and maternal selectivity in domestic sheep and goats: the two facets of maternal attachment. Dev Psychobiol 2007; 49:54–70. [ Links ]
42. ALEXANDER G, WILLIAMS DT. Teat–seeking activity in lambs during the first hours of life. Anim Behav 1966; 14:166–176. [ Links ]
43. NOWAK R, MURPHY TM, LINDSAY DR, ALSTER P, ANDERSSON R, UVNAS–MOBERG K. Development of a preferential relationship with the mother by the newborn lamb: importance of the sucking activity. Physiol Behav 1997; 62:681–688. [ Links ]
44. ADDAE PC, AWOTWI EK, OPPONG–ANANE K, ODDOYE EOK. Behavioural interactions between West African dwarf nanny goats and their single–born kids during the first 48 hours post–partum. Appl Anim Behav Sci 2000; 67:77–88. [ Links ]
45. DWYER CM, LAWRENCE AB. Maternal behaviour in domestic sheep (Ovis aries): constancy and change with maternal experience. Behav Processes 2000; 137:1391–1413. [ Links ]
46. LYNCH JJ, HINCH GN, ADAMS BD. The Behavior of Sheep. Biological principles and implications for production. Oxon UK: C A B International,1992. [ Links ]
47. SHILLITO–WALSER EE. A comparison of the role of vision and hearing in ewes finding their own lambs. Appl Anim Ethology 1978; 4:71–79. [ Links ]
48. LÉVY F, KELLER M, POINDRON P. Olfactory regulation of maternal behavior in mammals. Horm Behav 2004; 46:284–302. [ Links ]
49. SERAFIN N, TERRAZAS A, HERNANDEZ H, PAREDES A, POINDRON P. Maternal behavior of intact and anosmic parturient goats. Poster presented at the International Ethological Conference; 2003 August 20 –27; Florianapolis, Brazil, 2003: 32. [ Links ]
50.MARTINEZ M, OTAL J, RAMIREZ A, HEVIA ML, QUILES A. Variability in the behavior of kids born of primiparous goats during the first hour after parturition: effect of the type of parturition, sex, duration of birth, and maternal behavior. J Anim Sci 2009; 87:1772–1777. [ Links ]
51.DWYER CM. Behavioural development in the neonatal lamb: Effect of maternal and birth–related factors. Theriogenology 2003; 59:1027–1050. [ Links ]
*Intervet/Schering–Plough Animal Health, México.
**The Observer®, Noldus video–pro, versión 4.0, Holanda.

