











 nueva página del texto (beta)
nueva página del texto (beta) Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
Permalink
INTRODUCCIÓN
El filo Mollusca es uno de los componentes faunísticos bentónicos conocido en el golfo de California, con investigaciones que abarcan aspectos de sistemática, biología y ecología (Keen, 1971; Houston, 1980; Domínguez-Orozco & Tripp-Quezada, 1997; Holguín-Quiñones et al., 2000; Tripp-Quezada, 2008). En el archipiélago Espíritu Santo solamente dos estudios se han referido a los moluscos. El primero fue realizado por González-Medina et al. (2006), quienes evaluaron la biodiversidad de los moluscos y equinodermos de sustratos rocosos, mientras que el segundo (Tripp-Quezada et al., 2009) tuvo como objetivo utilizar a los moluscos de fondos blandos como indicadores de calidad ambiental, y para detectar zonas con asentamientos de moluscos de interés comercial.
Los moluscos, han sido considerados como grupo focal para realizar estudios de biodiversidad en el ambiente marino, ya que constituyen un buen indicador de la diversidad biológica total de los ecosistemas (Alcolado & Espinoza, 1996), tienen un amplio espectro trófico y presentan una elevada radiación evolutiva (Espinosa, 1992). Estas características, hacen muy atractiva la sugerencia de utilizar a los moluscos para determinar las variaciones en las comunidades ecológicas en una región determinada (Capetillo-Piñar et al., 2015). También pueden ser utilizados para evaluar el estado de salud y estrategias de manejo en las reservas naturales y áreas marinas de interés para la conservación, ya que son un excelente indicador de la riqueza de especies en los hábitats marinos tropicales y subtropicales (Espinosa & Ortega, 2001).
El archipiélago Espíritu Santo comprende varias islas e islotes en un área total de 102.076 km2 y se localiza en el límite oriental de la bahía de La Paz, que es uno de los cuerpos de agua de mayor extensión en el golfo de California (Arizpe, 1987; Casas-Valdez et al., 1997).
El archipiélago es parte del Área de Protección de Flora y Fauna “Islas del Golfo de California” (Diario Oficial de la Federación, 7 de junio de 2000) y actualmente cuenta con un plan de manejo (SEMARNAT/CONANP, 2001). Se ha observado un creciente interés turístico y ecológico por el archipiélago debido a su diversidad biológica y belleza paisajística, lo que ha propiciado el desarrollo de actividades de pesca ribereña, pesca deportiva y ecoturismo (Ezcurra, 2002). Sin embargo, se precisan estudios ecológicos, los cuales son escasos, que permitan aumentar el conocimiento de las variaciones en las comunidades bentónicas de fondos blandos, los que tributarían información complementaria para robustecer el plan de manejo que existe en la zona. Por tal motivo el objetivo del presente estudio fue determinar la variación espacial de los ensambles de los moluscos de fondos blandos someros del archipiélago, y describir la relación entre estos y el tipo de sustrato en su estructuración.
MATERIALES Y MÉTODOS
Área de estudio: El archipiélago Espíritu Santo se localiza al oriente de la bahía de La Paz, en el golfo de California (Fig. 1). Es un Complejo insular formado por dos islas mayores: La Partida, al norte, y Espíritu Santo, al sur, así como otras menores llamadas La Ballena, Los Islotes, El Gallo y La Gallina. El litoral oriental de las islas mayores es rocoso y homogéneo, presentando en el sur una extensa playa llamada La Bonanza. En contraste, su litoral occidental presenta pequeñas bahías, ensenadas, caletas, playas arenosas y lagunas bordeadas por manglares (Bourillón-Moreno et al., 1988).
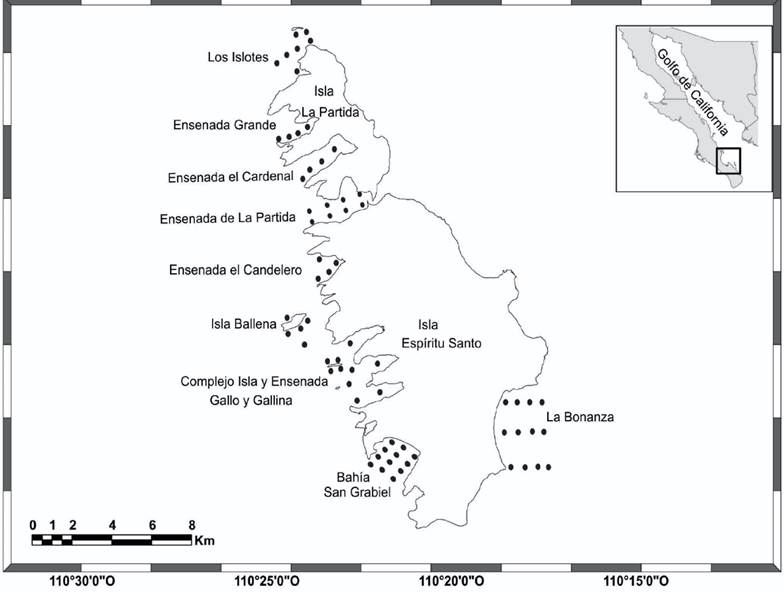
Figura 1 Zona de estudio y ubicación de las localidades y sitios de muestreo en el archipiélago Espíritu Santo, Baja California Sur, México.
Muestreo: Se realizó una campaña de muestreo en junio de 2013. Se establecieron en la zona infralitoral 66 sitios de muestreo, hasta una profundidad de 10 m, los que se distribuyeron cubriendo todos los tipos de fondos blandos observados en 9 localidades: 1- Playa La Bonanza (12 sitios), 2- bahía San Gabriel (12 sitios), 3- Complejo de Islotes y Ensenadas Gallo y Gallina (10 sitios) 4- Isla Ballena (5 sitios), 5- Ensenada Grande (4 sitios), 6- Ensenada El Candelero (4 sitios), 7- Ensenada La Partida (8 sitios), 8- Ensenada El Cardonal (4 sitios) y 9- Los Islotes (7 sitios). Los sitios de muestreos fueron considerados como replicas dentro de cada localidad. Las muestras se obtuvieron mediante buceo autónomo utilizando un marco de metálico de 1 m2 el cual se fijó al fondo en cada sitio de muestreo. Se recolectó el sedimento hasta 15 cm de profundidad y se tamizó in situ mediante una criba de madera de 40 x 40 cm con malla metálica de 3 mm de luz, siguiendo la metodología propuesta por Holme (1971). Se seleccionaron los organismos vivos, los cuales fueron guardados en bolsas de plástico para su traslado al laboratorio y su posterior identificación taxonómica con base en los trabajos de Keen (1971) y Abbott (1974). Los nombres científicos se actualizaron de acuerdo con Skoglund (1991, 1992), Coan et al. (2000), Coan et al. (2012) y la página web World Register of Marine Species (https://www.marinespecies.org). Los organismos recolectados fueron depositados en la colección de invertebrados marinos del Centro Interdisciplinario de Ciencias Marinas, IPN.
Para conocer el tipo de sustrato por localidad, se tomó una muestra de sedimento con un nucleador de 20 cm de largo y 7 cm de diámetro en cada sitio de muestreo de cada localidad, para un total de 66 muestras. El tamaño de grano se interpretó de acuerdo con la escala de Wentworth (Folk, 1980). El tipo de sustrato prevaleciente en las localidades se estimó como el porcentaje de sitios de muestreos donde aparecía un sustrato determinado con respecto al total de sitios de esa localidad. Se consideraron cinco tipos de sustrato: arena de grano medio, arena de grano fino, arena de grano muy fino, arena de grano grueso y arena de grano muy grueso.
Análisis de los datos: El esfuerzo de muestreo se evaluó mediante curvas de acumulación de especies basadas en el número de especies de cada localidad. Se utilizaron los estimadores no paramétricos Chao 2, Jackknife 1 y Jackknife 2 para determinar la riqueza esperada (Moreno, 2001).
Con el objeto de comparar las localidades con base en la estructura de los ensambles de moluscos, se analizaron los indicadores estructurales de abundancia (número de individuos), riqueza específica (S) estimada como el número de especies presentes en una muestra, la diversidad de acuerdo con el índice de Shannon-Wiener (H´) utilizando el logaritmo base 2, y la equidad (J) con el índice de Pielou (Moreno, 2001). Las comparaciones entre las localidades se realizaron mediante la prueba estadística Kruskal-Wallis (1952), ya que no se encontró normalidad ni homocedasticidad en las muestras. Cuando se detectaron diferencias estadísticas significativas (para un a= 0.05) se realizaron pruebas a posteriori de Dunn (z) con ajuste del grado de significación (p) por Bonferroni (Dunn, 1961), para detectar las localidades que eran diferentes. Los análisis se realizaron mediante el paquete estadístico SPSS V23.
La distribución de las especies en el archipiélago se determinó con base al porcentaje de localidades en el cual una especie estuvo presente. La distribución de las especies se clasificó de forma similar a López-Rojas et al. (2017), en: a) amplia, la especie se encontró en al menos el 70% de las localidades, b) frecuente, la especie se presentó entre el 50 y 69% de las localidades, c) limitada, la especie estuvo presente entre el 30 y 49% de las localidades y d) restringida, la especie se registró entre el 10 y 29% de las localidades. De esta forma se pudo medir la rareza de especies, considerando especies únicas, aquellas cuya ocurrencia fue en una sola localidad y duplicadas, las registradas en dos localidades (Kunin & Gaston, 1993).
Se construyó una matriz de similitud de Bray-Curtis, previa transformación de los datos de abundancia por especie con raíz cuarta (4√) (Field et al., 1982), a partir de la cual se realizó un análisis de clasificación. Los dendrogramas se construyeron con el método de agrupamiento de pares con la media aritmética no ponderada (UPGMA). La identificación de los grupos se realizó con la prueba de perfiles de similitud (SIMPROF) basada en promedio con 1000 permutaciones y 999 simulaciones con un a= 0.05 (Clarke & Gorley, 2015). Al encontrar diferencias significativas se identificaron los taxa que más contribuyeron a esas diferencias mediante la técnica de análisis SIMPER (Clarke, 1993, Clarke & Warwick, 2001).
Para conocer el efecto del tipo de sustrato sobre la estructura de los ensambles de moluscos, se realizó un análisis de varianza permutacional PERMANOVA (Anderson et al., 2008), sobre la base de la matriz de similitud de Bray Curtis de abundancia de especies. Se tomaron como variables los tipos de sustratos de los sitios de muestreo de cada localidad. Como factores fijos se utilizaron las Islas (dos niveles) y localidades (nueve niveles) y estas últimas anidadas dentro de Islas. Los tipos de sustratos (5 niveles) fueron aleatorios y anidados dentro de localidades e Islas. Los valores Pseudo-F se obtuvieron a partir de 999 (a = 0.05) permutaciones aleatorias.
Los análisis del esfuerzo de muestreo, dendrogramas con SIMPROF, SIMPER y PERMANOVA se realizaron usando el programa PRIMER v7 & PERMANOVA add on (Clarke & Gorley, 2015).
RESULTADOS
El análisis de ocurrencia del tipo de sustrato por localidad demostró que para el archipiélago, la arena de grano medio (54.5%) fue la de mayor distribución, seguido por la arena fina (22.7%), arena muy fina (15.9%), arena gruesa (6.8%) y arena muy gruesa (4.5%). La arena de grano medio fue el componente mayoritario (75 y 80% de los sitios) en seis localidades (Complejo de Islotes y Ensenadas Gallo y Gallina, Isla Ballena, Ensenada Candelero, Ensenada Cardonal, Ensenada Grande y Los Islotes). Las localidades restantes presentaron un sustrato conformado por una mezcla de arena de grano medio (27.5%), arena gruesa (52.5%) y arena fina (20%) en playa La Bonanza; arena muy gruesa (33.3%) y arena muy fina (66.6%) en bahía San Gabriel, y arena fina (57%) y arena muy fina (42.8%) en Ensenada La Partida.
Las curvas de acumulación de especies mostraron una tendencia a la asíntota al analizar las nueve localidades en su conjunto (Fig. 2), indicando una buena representación del esfuerzo de muestreo. De acuerdo con los estimadores Jackknife 1 y Jackknife 2, el valor máximo de especies esperadas estuvo entre 114 y 113 respectivamente, mientras que para Chao 2 fue de 106, lo que corresponde a una eficiencia de muestreo de 89.9%.
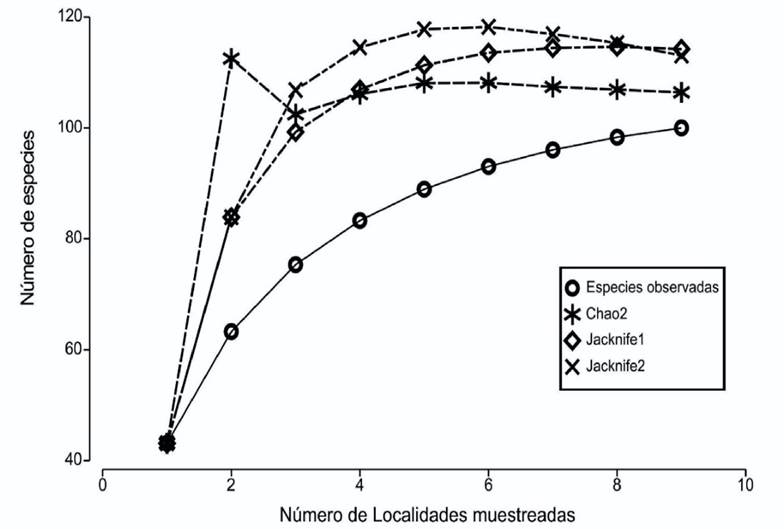
Figura 2 Curvas de acumulación de especies observadas y esperadas con índices no paramétricos Chao 2, Jacknife 1 y Jacknife 2 para el archipiélago Espíritu Santo.
Se registraron un total de 102 especies de moluscos (Anexo) incluidas en tres clases, 17 órdenes, 39 familias, y 70 géneros. Los bivalvos estuvieron representados por 49 especies, 9 órdenes, 17 familias, y 39 géneros. Los gastrópodos por 52 especies, 7 órdenes, 21 familias, y 30 géneros y los escafópodos por 1 especie.
La abundancia media de moluscos para el archipiélago fue de 2638.1 ± 1424.2 indv. m-2. La clase Bivalvia fue la que presentó la mayor abundancia (1855.5± 876 indv. m-2). Las familias mejor representadas fueron Lucinida y Veneridae con 7 (17.9%) y 6 (15.3%) géneros respectivamente, seguida por Arcidae con cinco géneros (12%) y Tellinidae con tres géneros (7.6%). El orden Venerida fue el mejor representado con tres familias (17.6%). La clase Gastropoda tuvo una abundancia de 753.5 ± 176 indv. m-2. Las familias mejor representadas fueron Calyptraeidae con tres géneros (10%), seguida de Olividae con dos géneros (6.6%).
Cuarenta especies estuvieron representadas por más de 50 individuos, de las cuales los bivalvos Eurytellina eburnea (Hanley, 1844), Cavilinga prolongata (Carpenter, 1857) y Ameritella coani (Keen, 1971), seguidas por los gastrópodos Siphonaria maura GB Sowerby I, 1835, Olivella dama (W. Wood, 1828) y O. gracilis (Broderip & GB Sowerby I, 1829), presentaron las mayores abundancias (Fig. 3).

Las variaciones observadas en la abundancia entre las localidades fueron estadísticamente significativas [KW-H(8;23 743) = 55.91; p = 0.002]. Estas diferencias se debieron al bajo valor registrado en Isla Ballena con respecto al resto de las localidades (Tabla 1). También a la alta abundancia de moluscos en Los Islotes vs. ensenada La Partida [z = 2.34, p = 0.04], Candelero [z = 3.02, p = 0.03] y el Complejo de islotes y ensenadas Gallo y Gallina [z = 3.45, p = 0.02], así como los valores bajos de las ensenadas Candelero y La Partida vs. La Bonanza [z = 1.60, p = 0.02 y z = 2.01, p = 0.01 respectivamente] (Fig. 4a). Los Islotes presentó la mayor abundancia (5269 indv. m-2), seguida por La Bonanza (3600 indv. m-2) y bahía San Gabriel (3345 indv. m-2) (Tabla 1).
Tabla 1 Indicadores estructurales de los ensambles de moluscos de fondos blandos de cada localidad del archipiélago Espíritu Santo. L. Bo.: La Bonanza, B.SG: bahía San Gabriel, C.IGG: Complejo de islotes y ensenadas Gallo y Gallina, I.Ba.: Isla Ballena, E.Cand.: Ensenada Candelero, E. Part.: Ensenada La Partida, E. Card.: Ensenada Cardonal, E. Grand.: Ensenada Grande, L. Islot.: Los Islotes. N.: abundancia total (No. indv m-2), S: riqueza de especies, H´: diversidad de Shannon (bits indv.-1), J´: equidad.
| Indicadores estructurales | L. Bo. | B. SG. | C. IGG | I. Ba. | E. Cand. | E. Part. | E. Card. | E. Grand | L. Islot. |
| N | 3600 | 3345 | 1800 | 127 | 2353 | 2020 | 3066 | 2163 | 5269 |
| S | 56 | 37 | 54 | 20 | 41 | 60 | 39 | 31 | 55 |
| H´ | 3.80 | 3.23 | 3.76 | 3.64 | 2.64 | 3.84 | 3.11 | 2.56 | 2.24 |
| J´ | 0.65 | 0.62 | 0.64 | 0.85 | 0.49 | 0.65 | 0.59 | 0.51 | 0.40 |
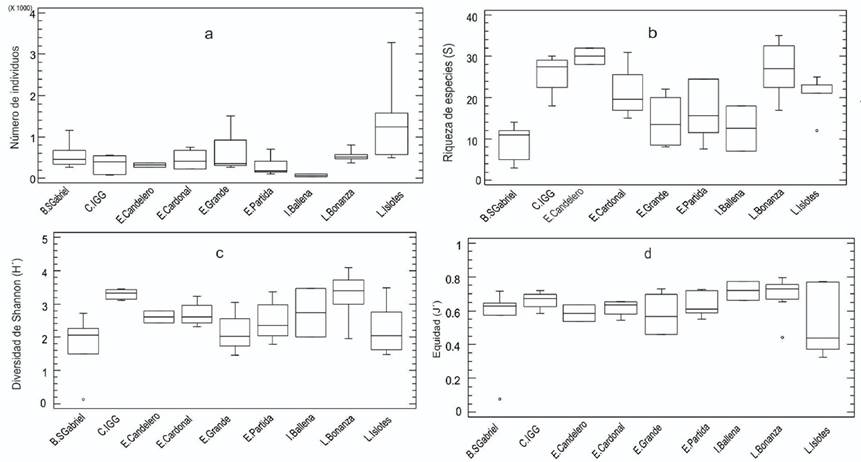
Figura 4 Variaciones de la abundancia (a), riqueza (S), diversidad (H´) y equidad (J´) de los ensambles de especies de moluscos bentónicos de fondos blandos en las localidades estudiadas del archipiélago Espíritu Santo.
En La Bonanza e isla Ballena la especie más abundante fue el gastrópodo S. maura (1271 indv. m-2. y 50 indv. m-2 respectivamente). Para bahía San Gabriel lo fue el bivalvo C. prolongata (1023 indv. m-2) y en el Complejo de islotes y ensenadas Gallo y Gallina los gastrópodos Neoterebra allyni (Bratcher & RD Burch, 1970) (392 indv. m-2) y O. dama (349 indv. m-2). Para las ensenadas Candelero y Grande la especie de mayor abundancia fue E. eburnea (1152 indv. m-2 y 1099 indv. m-2 respectivamente), mientras que para ensenada La Partida E. eburnea (600 indv. m-2) y el gastrópodo O. dama (288 indv. m-2) y en Los Islotes los bivalvos E. eburnea (2330 indv. m-2) y C. prolongata (2006 indv. m-2). En la ensenada Cardonal las especies más abundantes fueron los bivalvos C. prolongata (1027 indv. m-2), E. eburnea (595 indv. m-2) y el gastrópodo O. gracilis (502 indv. m-2).
El valor promedio de riqueza de especies para todo el archipiélago, teniendo en cuenta las nueve localidades, fue de 43.7 ± 13.5 especies, con un máximo de 60 especies en ensenada La Partida y mínimo de 20 en Isla Ballena. La equidad presentó un valor promedio de 0.6 ± 0.13, con un valor mínimo de 0.40 en Los Islotes y máximo (0.85) en Isla Ballena, mientras que la diversidad tuvo un valor promedio de 3.19 ± 0.59 bits indv.-1, con un máximo de 3.84 bits indv.-1 en ensenada La Partida y bajo (2.24 bits indv.-1) en Los Islotes.
Los indicadores estructurales de riqueza de especies, diversidad y equidad de cada localidad se muestran en la Fig. 4. Se encontraron diferencias significativas entre las localidades para la riqueza de especies [K-W(8,66) = 20.28; p = 0.002] y diversidad [K-W(8,66) = 15.19; p = 0.047], pero no para la equidad [K-W(8,66) = 12.28; p = 0.10]. Para la riqueza de especies, bahía San Gabriel fue diferente significativamente [z = 2.82, p = 0.003] a La Bonanza, ensenadas Cardonal [z = 2.34, p = 0.02] y Candelero [z = 1.80, p = 0.042], Los Islotes [z = 1.71, p = 0.032] y al Complejo de islotes y ensenadas Gallo y Gallina [z = 1.82, p = 0.04] (Fig. 4b).
Las diferencias encontradas en la diversidad fueron debidas a La Bonanza vs. bahía San Gabriel, Ensenada Grande y Los Islotes [z = 21.16, p = 0.001, z = 7.51, p = 0.01 y z = 15.60, p = 0.02, respectivamente] y al del Complejo de islotes y ensenadas Gallo y Gallina con respecto a bahía San Gabriel [z = 22.16, p = 0 005], Ensenada Grande [z = 19.70, p = 0.02] y Los Islotes [z = 16.60, p = 0.045] (Fig. 4c).
Para el archipiélago 16 especies (15.5%) presentaron una amplia distribución, de las cuales el bivalvo Brachidontes adamsianus (Dunker, 1857) se localizó en todas las localidades. El 20.5% (21 especies) fueron considerados taxa frecuentes, mientras que 30 (29.4%) se consideraron limitadas. Sin embargo, 36 especies (35.2%) fueron calificadas como restringidas (raras) de las cuales 16 (15.6%) fueron especies únicas, y 20 duplicadas (19.6%). Los bivalvos estuvieron representados por cinco especies únicas (4.9%) y ocho (7.8%) duplicadas, mientras que los gastrópodos por 11 únicas (10.7%), y 12 (11.7%) duplicadas. Ensenada La Partida fue la de mayor rareza de especies con 45 (44.1%), seguida por bahía San Gabriel con 32 (31.3%), mientras que las ensenadas Cardonal y Grande con 24 (23.5%) y 22 especies (21.5%), respectivamente.
De acuerdo con la estadística de Bray-Curtis, la similitud entre las localidades sobre la base de la composición y abundancia de las especies de moluscos se puede observar en la Fig. 5. El análisis SIMPROF distinguió un grupo (grupo A), conformado por seis localidades, con un nivel de similitud de 42%, (SIMPROF: π = 4.9; p = 0.001). Adicionalmente, tres localidades (La Bonanza, Complejo de islotes y ensenadas El Gallo y La Gallina e Isla Ballena) quedaron aisladas con valores bajos de similitud. Las localidades que conformaron el grupo A se dividieron en dos subgrupos: uno formado por ensenada La Partida y bahía San Gabriel con 47% de similitud, y otro por las ensenadas Candelero y Cardonal (65%) y ensenada Grande y La Lobera con 67.7% de similitud. Las localidades que quedaron aisladas presentaron bajos niveles de similitud y fueron estadísticamente diferentes entre sí y con respecto al grupo formado por La Bonanza (28.5%, SIMPROF: π = 5.9; p = 0.001), Complejo de Islotes y ensenadas Gallo y Gallina (24%, SIMPROF: π = 6.7; p = 0.001) e Isla Ballena (13%, SIMPROF: π = 8.4; p = 0.001).
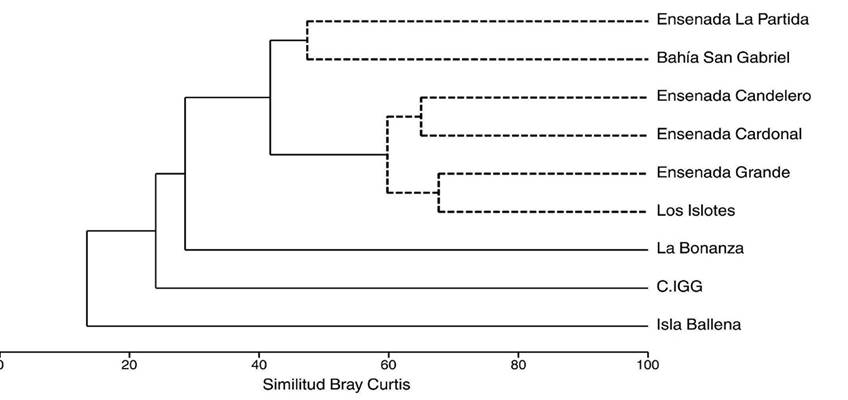
Figura 5 Dendrograma del análisis de similitud de los ensambles de moluscos entre las localidades del archipiélago Espíritu Santo. Las líneas segmentadas en el dendrograma muestran los grupos identificados por la prueba SIMPROF. C. IGG: complejo de islotes y ensenadas Gallo y Gallina.
Los resultados del análisis SIMPER para determinar la contribución de las especies en las agrupaciones de localidades se muestran en la Tabla 2. Para el Grupo A cinco especies (tres bivalvos y dos gastrópodos) fueron las que más contribuyeron a la similitud dentro del grupo. Cavilinga prolongata fue la que presentó la mayor contribución entre las localidades (Tabla 2). Las diferencias registradas entre el Grupo A y las localidades que quedaron aisladas se debieron a 10 especies de moluscos (cinco bivalvos y cinco gastrópodos). Los bivalvos Chione compta (Broderip, 1835) y E. eburnea, conjuntamente con el gastrópodo O. dama fueron los que más contribuyeron a las diferencias halladas (Tabla 2). Para las localidades aisladas se observó que las diferencias entre ellas fueron debido a cuatro especies de gastrópodos, siendo S. maura la que más contribuye a esas diferencias debido a su mayor abundancia en La Bonanza (Tabla 2).
Tabla 2 Especies que definieron la similitud/disimilitud entre las localidades analizadas del archipiélago Espíritu Santo, a través del SIMPER.
| Similitud Grupo A | ||||
| Especies | AP | SP | CTB% | |
| Cavalinga prolongata (Carpenter, 1857) | 13.22 | 10.22 | 25.49 | |
| Euritellina eburnea (Hanley, 1844) | 14.38 | 7.45 | 16.42 | |
| Olivella gracilis (Broderip y GB Sowerby I, 1829) | 18.81 | 8.24 | 13.34 | |
| Olivella dama (W. Wood, 1828) | 9.45 | 3.21 | 10.14 | |
| Ameritella coani (Keen, 1971) | 8.49 | 4.34 | 7.17 | |
| Disimilitud Grupo A vs. RL (L Bon, C. IGG, IBa) | ||||
| Especies | AP (RL) | AP (Grupo A) | DIP | CTB% |
| Chione compta (Broderip, 1835) | 2.87 | 15.53 | 10.39 | 12.42 |
| Euritellina eburnea (Hanley, 1844) | 1.41 | 13.06 | 9.03 | 10.41 |
| Olivella dama (W. Wood, 1828) | 0.00 | 5.37 | 10.16 | 10.33 |
| Siphonaria maura GB Sowerby I, 1835 | 0.14 | 12.06 | 8.14 | 9.53 |
| Olivella gracilis (Broderip y GB Sowerby I, 1829) | 0.00 | 11.10 | 8.22 | 9.18 |
| Chama sórdida Broderip, 1835 | 0.00 | 6.88 | 8.69 | 8.84 |
| Neoterebra allyni (Bratcher y RD Burch, 1970) | 9.56 | 0.05 | 6.81 | 7.71 |
| Cavalinga prolongata (Carpenter, 1857) | 9.09 | 1.84 | 6.29 | 7.16 |
| Ameritella coani (Keen, 1971) | 7.52 | 0.14 | 5.84 | 6.70 |
| Trimusculus reticulatus (GB Sowerby I, 1835) | 1.00 | 3.31 | 5.74 | 6.34 |
| Disimilitud RL | ||||
| Especies | AP | AP | DIP | CTB% |
| LBon | C. IGG | |||
| Siphonaria maura GB Sowerby I, 1835 | 12.06 | 0 | 7.37 | 8.53 |
| LaBon | IBa | |||
| Siphonaria maura GB Sowerby I, 1835 | 12.06 | 2.74 | 9.3 | 12.52 |
| Trimusculus reticulatus (GB Sowerby I, 1835) | 7.52 | 1 | 6.4 | 8.62 |
| C. IGG | IBa | |||
| Neoterebra allyni (Bratcher y RD Burch, 1970) | 9.56 | 0 | 9.1 | 9.74 |
| Olivella dama (W. Wood, 1828) | 9.09 | 0 | 8.82 | 9.44 |
Grupo A (ensenadas La Partida, Candelero, Cardonal, Grande, bahía San Gabriel y Los Islotes), RL (C. IGG: Complejo de Islotes y Ensenadas Gallo y Gallina, LBon: La Bonanza, IBa: Isla Ballena). AP: abundancia promedio (número de organismos), SP: similitud porcentual, DIP: disimilitud porcentual, CTB%: contribución porcentual.
El análisis PERMANOVA para conocer el efecto del tipo de sustrato sobre la estructura de los ensambles de moluscos conformados por gastrópodos y bivalvos y solamente de bivalvos se muestra en el Tabla 3. De las fuentes de variación analizadas las localidades presentaron diferencias estadísticas significativas, tanto para el análisis de los gastrópodos y bivalvos en conjunto, como para los bivalvos. Nótese como las localidades tuvieron el mayor coeficiente de variación en ambos análisis. Las diferencias observadas cuando se analizaron en conjunto a gastrópodos y bivalvos fueron debidas a La Bonanza vs bahía San Gabriel y el Complejo de islotes y ensenadas Gallo y Gallina [pseudo-t = 2.99, p = 0.03 y pseudo-t = 3.10, p = 0.001, respectivamente] y a la bahía San Gabriel vs. Complejo de islotes y ensenadas Gallo y Gallina [pseudo-t = 1.49, p = 0.001]. Sin embargo, para los bivalvos se debieron a la ensenada La Partida respecto a las ensenadas Cardonal y Grande [pseudo-t = 1.26, p = 0.001 y pseudo-t = 1.38, p = 0.001, respectivamente] y a Los Islotes [pseudo-t = 1.47, p = 0.0011].
Tabla 3 PERMANOVA con base en las similitudes de Bray-Curtis de los datos de abundancia de especies de moluscos gastrópodos y bivalvos (A) y de las especies de bivalvos (B) de las localidades estudiadas del archipiélago Espíritu Santo. Abreviaturas: PseudoF: valor seudo estadístico F, p: 0.05, CV: porcentaje del componente de variación. Is.: Islas, Lo.: Localidades. Los paréntesis indican los factores a los cuales se anidan.
| A: PERMANOVA para el ensamble de moluscos Gastrópodos y Bivalvos | |||||
| Fuentes de variación | df | MS | Pseudo-F | p | CV % |
| Islas (Is.) | 1 | 8405.3 | 1.35 | 0.231 | 11.11 |
| Localidades [Lo.] (Is.) | 7 | 6775.3 | 2.93 | 0.002 | 32.35 |
| Sustratos (Lo. (Is.)) | 4 | 2026.3 | 0.94 | 0.566 | 6.53 |
| Residual | 31 | 2175.5 | 46.34 | ||
| Total | 43 | ||||
| B: PERMANOVA para el ensamble de moluscos Bivalvos | |||||
| Islas (Is.) | 1 | 7500.5 | 1.09 | 0.33 | 6.07 |
| Localidades [Lo.] (Is.) | 7 | 7403 | 3.212 | 0.001 | 35.02 |
| Sustratos (Lo. (Is.)) | 4 | 2088.1 | 0.99371 | 0.478 | 2.15 |
| Residual | 29 | 2101.3 | |||
| Total | 41 | ||||
DISCUSIÓN
Las proyecciones de las curvas de acumulación de especies demostraron que para tener una completitud del inventario de especies de moluscos del archipiélago faltarían por registrarse entre 4 y 12 especies. Por lo anterior, se puede confirmar que el esfuerzo de muestreo realizado fue suficiente para calcular el número teórico total de especies de moluscos de esta región. Resultado similar fue obtenido por Esqueda-González et al. (2014) al usar cuadrantes como técnica de muestreo en los ambientes de zona intermareal y submareal en la bahía de Mazatlán para estudiar la composición, riqueza y distribución de los moluscos bivalvos marinos de la bahía.
La riqueza de especies de moluscos de fondos blandos someros registrada en este trabajo fue mayor a la obtenida por Tripp-Quezada (2008) en otras localidades (Cabo Pulmo 84, Isla San José 58, Punta Chivato 86 y Bahía de los Ángeles 91 especies) del golfo de California. También fue superior a lo reportado para Isla Santa Catalina (65) y San Francisco (80) (Tripp-Quezada et al., 2014) e Isla Santa Cruz (44) (Tripp-Quezada et al., 2018), aunque inferior a la reportada en Isla Cerralvo (133) por Vázquez-Vega (2013) y en bahía Chamela con 293 (teniendo en cuenta a los gastrópodos, bivalvos y escafópodos) (Ríos-Jara et al., 2020). Estos hallazgos pueden ser atribuidos a las características de cada zona (p.e., hidrodinámica, variedad de fondos blandos) y/o al efecto del esfuerzo y método de recolecta de muestras. Por ejemplo, Tripp-Quezada (2008) realizó colectas de moluscos con un cilindro de 30 cm de diámetro, mientras que en el presente trabajo fue mediante un marco de metálico de 1m-2, lo que evidencia que, al menos, el área de muestreo que abarcó cada arte fue muy diferente y pudo haber tenido incidencia en las diferencias encontradas.
Respecto a los registros en las islas Santa Catalina, San Francisco, Santa Cruz y Cerralvo, en las cuales se aplicó el mismo método de colecta que en el presente estudio, las diferencias pueden atribuirse a factores ambientales típicos de cada isla en particular. En isla Santa Cruz, donde predominan las arenas gruesas, se observó que los moluscos se encontraron asociados con fragmentos de coral, algas calcáreas y conchas de moluscos muertos. En este tipo de sustrato fue frecuente la ocurrencia del gastrópodo S. maura, aunque es un organismo típico de sustratos rocosos. Situación similar ha sido reportada para isla San José (Tripp-Quezada, 2008). Para isla Cerralvo, donde se encontraron cinco tipos de sedimento, las arenas gruesas tuvieron mayor cobertura y presentaron la mayor abundancia y riqueza de moluscos (Vázquez-Vega, 2013), lo que demuestra que la complejidad y heterogeneidad de este tipo de sedimento (Méndez et al., 1986), puede influir de manera directa en la riqueza y/o composición de especie dentro y entre localidades. Sin embargo, la presencia de 37 (36.2%) especies con una amplia y frecuente distribución en el archipiélago dio evidencias de cierta estabilidad ambiental en el momento de realizar los muestreos. Una causa pudo haber sido la presencia de sustrato arenoso de grano medio en más del 50% de las localidades estudiadas.
La ocurrencia en el 100% de las localidades del bivalvo B. adamsianus y en un 88% de seis especies de bivalvos (E. erburnea, A. coani, C. prolongata, Chama sordida Broderip, 1835, Tivela byronensis (Gray, 1838) y Chione undatella (GB Swerby I, 1835)), las cuales han sido colectadas en el sustrato arenoso de grano medio, justifica que pueden ser consideradas como representativas del archipiélago, con potencialidad para ser usadas en estudios de monitoreo ambiental. Al respecto, B. adamsianus es una especie que ha sido registrada con una amplia distribución en regiones marinas prioritarias en el estado de Guerrero (López-Rojas et al., 2017). Además, E. eburnea que fue la especies más abundante (6178 indv.) en este estudio, resultado similar a los registrados en isla San José (153 indv.) y bahía de los Ángeles (274 indv.) (Tripp-Quezada et al., 2009), Cabo Pulmo (1749 indv.) (Tripp-Quezada, 2008) y las islas Santa Cruz (10% abundancia porcentual) (Tripp-Quezada et al., 2018) y Cerralvo (3717 indv.) (Vázquez-Vega, 2013), puede ser propuesta como indicador de cambio de las condiciones ambientales de esas islas.
La ocurrencia de especies en una localidad puede justificar las diferencias halladas en cuanto a la riqueza de especies entre localidades. Los gastrópodos Conasprella arcuata (Brodrip & GB Sowerby I, 1829), C. perplexa (GB Sowerby II, 1857) y C. ximenes (Gray, 1839), conjuntamente con Oliva polpasta Duclos, 1833 y O. porphyria (Linneo, 1758) fueron registradas solamente en ensenada La Partida. Esta localidad se caracterizó por la presencia de un sustrato de arena fina a diferencia a las halladas en isla Ballena y las ensenadas Candelero y Grande, donde sus ocurrencias fueron casi nulas y el tipo de sustrato mayoritario fue la arena de grano medio. En este contexto, se conoce que las especies del género Oliva tienen preferencias por sustratos arenosos finos y/o areno fangoso, donde les resulta fácil enterrase para refugiarse de sus depredadores y por la disponibilidad de alimento (i.e., la especie es de hábitos carroñeros aunque puede alimentarse de presas vivas de poca movilidad como los bivalvos, Torres-Palacio, 2008). Este hallazgo dio evidencias que las condiciones ambientales que prevalecieron en esta localidad fueron determinantes en limitar la distribución de estas especies en el archipiélago, al ofrecer las mejores condiciones para su desarrollo.
Un caso interesante fueron los escafópodos. Estos organismos estuvieron representados por la especie Antalis pretiosa (GB Sowerby II, 1860) con una baja abundancia y distribución limitada, al colectarse en tres localidades (Complejo de islotes y ensenadas Gallo y Gallina (3 indv. m-2), ensenada La Partida (8 indv. m-2) y Los Islotes (13 indv. m-2). Las mayores abundancias se localizaron en arena de grano medio, resultados que difieren de los reportados por Corgos et al. (2014), al registrar varias especies del género Dentalium con mayores abundancias (1290 indv. m-2) en sustratos de sedimentos finos (limos y arcillas) con alto contenido de materia orgánica. A su vez, son similares a los reportados por Ríos-Jara et al. (2020) al registrar organismos de este grupo en fondos arenosos (4 -10 m de profundidad) en dos lugares (afuera de Villa Polinesia e isleta San Andrés) en bahía de Chamela en el Pacífico mexicano, mostrando que son organismos que presentan una rareza ecológica relativamente alta. Estos autores asociaron esa distribución restringida a la preferencia de los escafópodos por hábitats con presencia de materia orgánica y alimento (p.e foraminíferos).
La escasa literatura acerca de los escafópodos en las costas del Pacífico mexicano (Ríos-Jara et al., 2003), no permite hacer un análisis más profundo sobre la ocurrencia de estos organismos en este estudio. Sin embargo, como los escafópodos son organismos excavadores y semisésiles de pequeño tamaño, son más abundantes en sedimentos finos, fangosos y fango-arcillosos, se sugiere que su presencia pudiera asociarse a la existencia de materia orgánica en el sedimento. Además, si se tiene en cuenta que en las muestras analizadas se observaron foraminíferos, los que son indicadores de enriquecimiento por materia orgánica (Gómez-Noguera & Hendrickx, 1997), y forman parte de la dieta de los escafópodos, es una evidencia de condiciones que pudieron favorecer la ocurrencia de este grupo, la cual debe ser analizada en futuras investigaciones.
Bouchet et al. (2002) argumentaron que cuando un muestreo se restringe a un solo periodo del año, es difícil tener una cobertura completa a todas las especies de una localidad, debido a que algunos organismos tienen ocurrencias muy estacionales o esporádicas. Con base en este argumento, el resultado obtenido acerca de la rareza de especie para el archipiélago no es suficiente y debe verse con cautela y sin llegar a conclusiones definitorias, ya que se precisa realizar muestreos en varias estaciones del año, el cual no fue realizado en este estudio.
La mayor abundancia de los bivalvos con respecto a los gastrópodos pudiera ser una consecuencia de la preferencia de estos organismos por fondos blandos con tamaño de grano de medio a fino y de los gastrópodos por sedimentos más gruesos, lo cual ha sido confirmado por varios autores (Domínguez-Orozco, 1996, Vázquez-Vega, 2013; Tripp-Quezada et al., 2018). La localidad Los Islotes, que se caracterizó por presentar sedimentos de grano medio, fue la que tuvo la mayor abundancia de moluscos debido al bivalvo C. prolongata, considerada un micro molusco, la cual parece tener preferencia por este tipo de sedimento. Además, cabe mencionar que las especies Septifer zeteki Hertlein & AM Strong, 1946 y E. eburnea, las que han sido reportadas en varias zonas del golfo con cierta abundancia (Tripp-Quezada, 2008; Tripp-Quezada et al., 2009, 2018) y que fueron consideradas como micro moluscos o juveniles, también mostraron preferencias por sedimentos de grano medio. Tapía-Diaz (2018) al realizar un estudio de los micro moluscos de Nayarit, registró un incremento en la diversidad de este grupo en zonas con arenas de grano medio, resultado similar al observado en el presente estudio. El termino micro molusco refiere a moluscos adultos cuya dimensión varía entre los 5 y 10 mm (Geiger, 2007; Ortigosa et al., 2018). Dentro de estas tallas se encuentran los estadios juveniles de muchos moluscos que en estado adulto poseen un tamaño mayor, hecho que dificulta la comparación entre trabajos realizados. Sin embargo, como se les consideran buenos indicadores de la estructura de las comunidades del bentos y pueden reflejar alteraciones ambientales (Kay, 1980), sería recomendable dirigir esfuerzos para conocer el estado de las comunidades de estos organismos en el archipiélago.
Las arenas de grano medio presentaron mayor cobertura en el archipiélago y las mayores abundancias y riqueza de las especies de bivalvos, mientras que los gastrópodos la tuvieron en los sedimentos de grano grueso, resultado que no difiere a lo reportado por Tripp-Quezada (2008), Tripp-Quezada et al. (2009, 2014, 2018) y Vázquez-Vega (2013) para otras Islas del golfo de California. Lo anterior sugiere que la prevalencia de un tipo de sustrato puede ser determinante en la abundancia y composición de estos organismos. En ensenada La Partida, localidad donde los sedimentos estuvieron dominados por grano fino a muy fino, se registró la mayor diversidad de bivalvos, coincidiendo con lo reportado por Domínguez-Orozco (1996).
La relación de las especies de ambas clases por cierto tipo de sustrato incidió en la formación de grupos de localidades. Por ejemplo, los bivalvos E. eburnea y C. prolongata fueron las que más contribuyeron a la similitud hallada en seis localidades (ensenadas La Partida, Candelero, Cardonal, Grande, bahía San Gabriel y Los Islotes), las que se caracterizaron por presentar sustratos con arenas de grano medio y fino. Mientras que el gastrópodo O. dama y los bivalvos E. eburnea y C. compta fueron quienes más contribuyeron a las diferencias entre esas seis localidades con respecto a La Bonanza, Complejo de islotes y ensenadas Gallo y Gallina e isla Ballena. Sin embargo, las diferencias de estas tres últimas localidades, las que se caracterizaron por presentar arena gruesa (principalmente en La Bonanza) y media, fueron debidas a las especies de gastrópodos S. maura y N. allyni fundamentalmente, y al bivalvo Hyotissa hyotis (Linnaeus, 1758) en isla Ballena, en arena de grano medio. Respecto a S. maura, Tripp-Quezada et al. (2018) registraron en isla Santa Cruz, la mayor abundancia de esta especie de gastrópodo asociada en granos de arena gruesa, fragmentos de coral, algas calcáreas y conchas de moluscos muertos, resultado similar al obtenido en este estudio. Este hallazgo es de interés, ya que justifica la presencia de esta especie en fondos blandos, cuando es típica de fondos rocosos. Por ejemplo, en La Bonanza, 10 (83.3%) de 12 sitios de muestreo, se caracterizaron por presentar material biogénico conformado por fragmentos de coral y conchas de moluscos muertos y fue la localidad donde se hallaron las mayores ocurrencias y abundancias de S. maura y de la mayoría de las especies de la familia Calyptraeidae, que son de fondos rocosos. Todo indica que la ocurrencia de especies típicas de fondos rocosos en fondos blandos esta mediada por la presencia de material biogénico (trozos de coral, algas calcáreas, conchas de moluscos muertos).
Otro aspecto importante para destacar es que el método de recolecta de moluscos realizado en el presente estudio favoreció, en alguna medida, la captura de estas especies facilitando su presencia en las muestras.
Ha sido demostrado que la distribución y abundancia de los moluscos está determinado por las características texturales de los sedimentos (Domínguez-Orozco, 1996; González-Medina et al., 2006; Vázquez-Vega, 2013, Tripp-Quezada et al., 2018). Sin embargo, los resultados del PERMANOVA, para las especies de gastrópodos y bivalvos, así como para los bivalvos (Tabla 3) demostraron que el factor localidad fue el que explicó las diferencias observadas en los ensambles de moluscos. Nótese como en ambos análisis el factor localidad tuvo el mayor coeficiente de variación.
La presencia de canales de mangles en algunas localidades del archipiélago, quienes tienen el potencial de aportar materia orgánica y nutrientes al medio, pudieron modificar las características de los sedimentos y por consiguiente haber determinado cambios en las comunidades de moluscos, pudiendo favorecer a unas especies más que a otras. Como ejemplo se puede considerar a ensenada El Candelero, localidad donde dominó la familia Veneridae con el mayor número de géneros, de los cuales Chione fue el mejor representado y que se caracteriza por preferir lugares ricos en materia orgánica para su desarrollo (García-Domínguez, 1991; Capetillo-Piñar et al., 2015). Este hecho estuvo probablemente relacionado con la presencia en esta localidad de un mayor número de canales de mangles, todo lo contrario, en bahía San Gabriel, en el que no hay presencia de estos. Lo anterior coincide con lo planteado por Snelgrove & Butman (1995), acerca de que además del tipo de sedimento, la materia orgánica puede ser un factor condicionante para el desarrollo de una comunidad, presentándose altas abundancias de organismos donde los sedimentos son más finos (De la Lanza, 1986).
La diversidad promedio de especies estimada en este archipiélago (3.2 bits/indv.) al compararla con otras regiones del golfo de California como Punta Chivato (3.8 bits/indv.) Cabo Pulmo (1.2 bits/indv) (Tripp-Quezada, 2008), isla San Francisco (2.7 bits/indv) (Tripp-Quezada et al., 2014), isla Santa Cruz (2.6 bits/indv), isla Cerralvo (1.58 bits/indv.) (Vázquez-Vega, 2013), mostró en la mayoría de los casos un valor alto, excepto a Punta Chivato. Este resultado puede deberse a las características ambientales (p.e., hidrodinámica, variedad de tipos de fondos blandos) de cada lugar, de las cuales en el archipiélago resalta como máxima diferencia la presencia de manglares y a su posición geográfica en el límite oriental de la bahía de La Paz (Hausback, 1984). La circulación de las aguas superficiales de la bahía está ligada al patrón de vientos estacionales y a un sistema relativamente estable de corrientes costeras generadas por las mareas (Obeso-Nieblas, 1986), las cuales pueden transportar sedimentos hacia el golfo. Por tanto, la posición geográfica del archipiélago puede favorecer que se depositen y acumulen en su costa occidental (donde se ubican las localidades de estudio) los sedimentos y materia orgánica, trayendo consigo una mayor abundancia y diversidad de moluscos. Además, en la época de verano los vientos del sureste generan surgencias en la costa peninsular (Lara-Lara et al., 1993), las cuales pueden adicionar un aporte más de nutrientes en el archipiélago.
La no diferencia significativa de equidad entre las localidades fue similar a los resultados obtenidos por Espinosa (1992), Tripp-Quezada (2008) y Capetillo-Piñar (2016). En este contexto, la constancia de las mismas especies entre las localidades aun siendo estas dominantes, así como la permanencia e intensidad de algunos factores limitantes a la dominancia (p.e la depredación), pueden explicar dicho resultado. Este hallazgo induce a pensar que las condiciones ambientales, de manera general, fueron estables en la región por lo que no han alterado significativamente la estructura de los ensambles de moluscos, al menos en la etapa en la que se realizó el presente estudio.
En sentido general, las variaciones espaciales observadas en los ensambles de moluscos de fondos blandos del archipiélago Espíritu Santo, parecen estar sujetas al efecto ocasionado por los manglares mediante el subsidio de diferentes cantidades de detritus hacia la zona marina, el que puede influir en el sustrato de algunas localidades. Jiménez-Ramos et al. (2019) en un sistema lagunar de la costa Oriental de Venezuela, observaron una riqueza de especies de moluscos inusual en sedimentos desprovistos de vegetación adyacentes a manglares. Este hallazgo les permitió afirmar que la convergencia de diferentes hábitats es un factor que influye en la riqueza y diversidad de organismos de una zona. Robertson et al. (1992) observaron en un manglar de Australia, que solo el 20% de la materia orgánica se degradaba y el resto era exportada a la zona marina, incidiendo en la composición de las comunidades bentónicas. Orihuela et al. (2004), al analizar el flujo de materia en un manglar de la costa de Chiapas, determinaron que el detritus exportado a las zonas adyacentes contenía un 54.4% de materia orgánica con calidad aceptable para tolerar un forrajeo intenso por diferentes organismos. Por lo expuesto con anterioridad, se puede decir que independientemente del tipo de sedimento la estructura de una comunidad puede ser condicionada por otros factores, como puede ser la materia orgánica. No obstante, se sugiere comenzar un estudio espacio temporal que abarque análisis del contenido de materia orgánica en sedimentos para comprobar la afirmación anteriormente expuesta.

