











 nueva página del texto (beta)
nueva página del texto (beta) Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
Permalink
INTRODUCCIÓN
El urohial es un hueso laminar alargado, es angosto en dirección anterior y ancho en dirección posterior, está incrustado en los músculos del istmo en el centro de la mandíbula inferior de los teleósteos (Ghasemzadeh, 2016). Esta estructura ha sido considerada de amplia utilidad en estudios sobre osificación en etapas larvarias (Aprieto, 1973); descripciones anatómicas (Chollet-Villalpando, 2009; Ghasemzadeh, 2016; Kobelkowsky & Resendez, 1972); determinación de edad (Johal et al., 2000); identificación de restos ictioarqueológicos (Patón et al., 1991); identificación de especies (De La Cruz-Agüero & Chollet-Villalpando, 2012); discriminación entre familias (principalmente Gerreidae), géneros y especies de peces, describiendo su morfología con fines taxonómicos y filogenéticos (Arratia & Schultze, 1990; Chollet-Villalpando, 2012; Chollet-Villalpando et al., 2014; González-Acosta et al., 2014; Jawad et al., 2016); y también su probable rol en el movimiento y comportamiento de alimentación en teleósteos (Khaah et al., 2014).
La relación entre la forma de las estructuras de los organismos y el papel de las especies en su comunidad permite conocer la variación morfológica existente entre especies y taxa derivados de la variación adquirida en el medio ambiente (Leisler & Winkler, 1985). Es así como, dada la enorme variedad de tipos de presas disponibles para los peces, el funcionamiento en la alimentación, definido por la forma y el desempeño de éstos, establece límites discretos en lo que pueden ingerir las especies, definiendo además lugares preferentes de alimentación (Bohórquez-Herrera, 2009).
De acuerdo con Cardona (2016) la dieta de la mayoría de las especies del género Mugil, se basa en materia orgánica presente en el sedimento, aunque también pueden consumir invertebrados bentónicos, macroalgas verdes filamentosas y plancton, siendo el microfitobentos un componente importante de su dieta. Según Sánchez-Rueda et al. (1997), M. curema y M. cephalus se caracterizan por presentar un espectro trófico y un hábitat muy similar, no obstante, M. Curema presenta afinidad por los limos gruesos 63-32 mm mientras que M. cephalus tiende a consumir una mayor proporción de arenas muy finas 125-63 mm lo que confirma su carácter selectivo. Igualmente, la selectividad en el espectro trófico posiblemente se relacione con diferentes zonas de alimentación lo que evitaría la competencia interespecífica por alimento. A lo largo de las zonas costeras mexicanas ambas especies coexisten, pero los picos máximos de desove se presentan desfasados (muestran tres meses de retraso en el norte del Golfo de México), dando como resultado retrasos en el reclutamiento de juveniles a las zonas estuarinas posiblemente para evitar la sobreposición en la alimentación de los juveniles que consumen el zooplancton limitante en las zonas costeras (Ibáñez & Colín, 2014; Ibáñez & Gutiérrez-Benítez, 2004).
De acuerdo con Kusaka (1974) citado en González-Acosta et al. (2014) el hueso urohial constituye un elemento fundamental para el mecanismo de apertura bucal de los peces, por lo que la morfología de esta estructura podría sugerir un acoplamiento causal tanto entre los mecanismos de alimentación como en la discriminación entre especies y poblaciones de distintas localidades. En este sentido sería útil en la discriminación entre especies y posiblemente también entre localidades.
Con el objeto de determinar la utilidad de la forma del hueso urohial se analizaron ejemplares de Mugil curema y M. cephalus de distintas localidades, dos del Golfo de México y una del Pacífico Mexicano, con el objeto de discriminar entre especies y localidades.
MATERIAL Y MÉTODOS
Los ejemplares se obtuvieron de la pesca comercial ribereña de tres zonas de estudio: una del Pacífico Mexicano, Puerto Madero, Chiapas; y dos del Golfo de México, Puerto La Frontera, Tabasco; y Laguna de Alvarado, Veracruz (Fig. 1).

Figura 1 Localidades de colecta. Puerto Madero, Chiapas; Frontera, Tabasco y laguna de Alvarado, Veracruz. Modificada de INEGI, Ita-Rubio, Escamilla-Herrera, C. & Soto-Núñez (1990).
Los organismos fueron congelados y transportados al laboratorio de Biología Pesquera de la Universidad Autónoma Metropolitana unidad Iztapalapa, para su posterior análisis. Se obtuvieron 142 ejemplares en total; 91 de los cuales fueron de M. curema y 51 de M. cephalus (Tabla 1).
Tabla 1 Tamaño de muestra por especie y localidad. LS= Longitud standard (mm).
| Fecha de colecta | No. Huesos urohiales | LS (mm) | ||
| Rango (min-máx) | Media ± σ | |||
| M. curema | ||||
| Puerto Madero, Chiapas | Oct-2019 | 31 | 163 - 255 | 212 ± 21.9 |
| Frontera, Tabasco | Nov-2018 | 31 | 222 - 300 | 267 ± 22.4 |
| Alvarado, Veracruz | Oct-2018 | 29 | 210 - 283 | 239 ± 17.3 |
| Total | 91 | |||
| M. cephalus | ||||
| Puerto Madero, Chiapas | Oct-2019 | 8 | 294 - 373 | 327 ± 25.3 |
| Frontera, Tabasco | Oct-Dic-2018 | 17 | 248 - 354 | 308 ± 36.3 |
| Alvarado, Veracruz | Oct-2018 y Jul-2019 | 26 | 323 - 468 | 398 ± 39.5 |
| Total | 51 | |||
La extracción del urohial se llevó a cabo por medio de una serie de incisiones en la parte ventral del organismo, separando las membranas branquióstegas hasta la altura del istmo y desprendiendo los ligamentos anteriores que lo unen con los basibranquiales (Bemis et al., 2004). Posteriormente, se realizó la extracción de cada urohial junto con su paquete muscular. Cada urohial se sometió a cocción en una parrilla eléctrica modelo American 6185, con agua corriente dentro de un frasco metálico durante 10 minutos a una temperatura aproximada de 80°C. Subsecuentemente se retiró por completo cualquier tipo de residuo empleando una aguja de disección, un bisturí, solución de agua con jabón convencional y agua corriente para su enjuague. Finalmente, cada urohial se preservó en frascos plásticos y alcohol al 70%.
Para el análisis morfométrico se emplearon tres vistas de la estructura ósea urohial: lateral izquierda, dorsal y ventral. Los urohiales fueron fotografiados con una cámara digital marca Sony, modelo DSC-W650, usando un trípode para cámara, una escala de medición y fondo oscuro. Cada imagen obtenida fue renombrada con respecto al número de ejemplar, especie, localidad de origen y vista de la estructura.
La definición de landmarks se realizó con respecto a la homología observada de la estructura del urohial de todos los ejemplares, es decir, aquellas partes consistentes y de clara identificación visual, así se seleccionaron solamente landmarks tipo I. Aquellos urohiales que estuvieron rotos e incompletos fueron descartados para el análisis.
Al no contar con estudios de esta índole y metodología previa, se consideraron apropiadas seis landmarks para las tres vistas, lateral, dorsal y ventral (Fig. 2). Los landmarks se registraron utilizando el software tpsDig2 versión 2.31 (Rohlf, 2017).

Con el programa Morphologika2 versión 2.5 (O’Higgins & Jones, 2007) se realizó el análisis generalizado de Procrustes (AGP) el cual consistió en escalar, trasladar y rotar el conjunto de landmarks para generar una imagen consenso y obtener los componentes principales (CP) con los cuales se realizaron los análisis discriminantes para explorar patrones de agrupación en los datos y reducción de la dimensionalidad de los datos multivariados (Zelditch et al., 2004). También se realizó un análisis multivariado de la covarianza (MANCOVA) utilizando como covariable y factores de agrupación la longitud estándar, especie y localidad, respectivamente. Finalmente, la forma fue visualizada en los extremos de las funciones discriminantes (Bookstein, 1989).
RESULTADOS
Discriminación entre especies. El primer CP explicó 39.5, 55.4 y 43.7% de la varianza total mientras que el segundo CP explicó 25.8, 18.4 y 25.9% en las tres vistas del urohial, lateral, dorsal y ventral respectivamente. En la figura 3 A, B y C se observa claramente la separación de las dos especies, donde la variación de la forma del hueso urohial se aprecia en la representación de las cuadrículas en cada figura, así para M. curema la forma de la vista lateral del urohial (Fig. 3A) muestra que dicho hueso es cóncavo en dirección ventro-dorsal donde la distancia entre la ubicación relativa de los landmark 1 y 6 es muy corta, mientras que en M. cephalus el urohial es convexo en dirección dorso-ventral y los landmark 1 y 6 tienen una ubicación relativa más alejada entre sí. La forma del hueso urohial de M. curema, en la vista dorsal (Fig. 3B) y ventral (Fig. 3C), se diferencia de la de M. cephalus por ser más angosto, es decir, todos los landmarks tienen distancias relativas muy cercanas entre sí, mientras que en M. cephalus la forma del hueso es más ancho (Fig. 3).
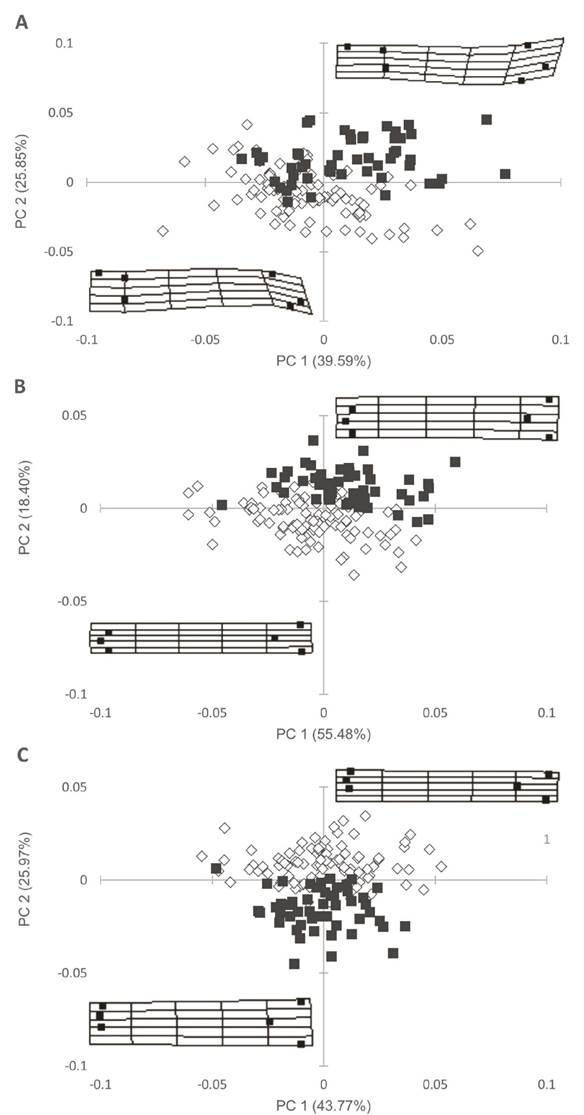
Figura 3 Análisis de componentes principales entre especies; A, vista lateral; B, vista dorsal; C, vista ventral; M. curema (rombo) y M. cephalus (cuadrado).
La discriminación entre especies fue de 86.6, 96.5 y 95.1% (Wilks’ λ = 0.422; 0.234 y 0.252, respectivamente y p < 0.001 para los tres casos), para la vista lateral, dorsal y ventral, respectivamente. Los análisis de MANCOVA confirman diferencias significativas en la forma del urohial entre especies de todas las vistas donde la longitud estándar mostró diferencias significativas para la vista lateral y ventral y no para la vista dorsal (Tabla 2). El porcentaje de la forma explicada por la longitud estándar fue 10.81, 2.38 y 5.45% para la vista lateral, dorsal y ventral, respectivamente.
Tabla 2 Resultados de la clasificación empleando validación cruzada entre las especies M. curema y M. cephalus. LS= Longitud standard (mm).
| Predicción de pertenencia al grupo | MANCOVA | ||||||||
| Vista | % | M. curema | M. cephalus | Total | Wilks’ λ | p | Efecto | Wilks’ λ | p<0.05 |
| Laterala | M. curema | 89.0 | 100 | 0.422 | <0.001 | LS | 0.874 | 0.020 | |
| M. cephalus | 82.4 | 100 | Especie | 0.650 | <0.001 | ||||
| Dorsalb | M. curema | 95.6 | 100 | 0.234 | <0.001 | LS | 0.906 | 0.102 | |
| M. cephalus | 98.0 | 100 | Especie | 0.404 | <0.001 | ||||
| Ventralc | M.curema | 93.4 | 100 | 0.252 | <0.001 | LS | 0.812 | <0.001 | |
| M. cephalus | 98.0 | 100 | Especie | 0.443 | <0.001 | ||||
a 86.6% de la validación cruzada correctamente clasificada.
b 96.5% de la validación cruzada correctamente clasificada.
c 95.1% de la validación cruzada correctamente clasificada.
Discriminación entre localidades. El porcentaje de varianza explicada por el primer CP para las localidades en M. curema fue de 47.3, 61 y 60.9% mientras que el segundo CP mostró 16.1, 15.3 y 17.5%, con una varianza total acumulada de 63.4, 76.3 y 78.4%, para la vista lateral, dorsal y ventral respectivamente. El resultado del análisis discriminante por localidades, de esta especie, fue de 65.9, 54.9 y 60.4% (Wilks’ λ = 0.354; 0.478 y 0.448, respectivamente y p < 0.001 para los tres casos), para la vista lateral, dorsal y ventral, respectivamente donde el mejor porcentaje de clasificación para todas las vistas del urohial fue la localidad de Chiapas (Tabla 3). El MANCOVA confirma diferencias significativas en la forma del urohial de todas las vistas entre localidades para M. curema, mientras que la longitud estándar no resultó ser significativa para la forma. El porcentaje de la forma explicada por la longitud estándar fue 4.39, 5.07 y 6.97% para la vista lateral, dorsal y ventral, respectivamente.
Tabla 3 Resultados de la discriminación con validación cruzada entre las localidades de Mugil curema. LS= Longitud standard (mm).
| Predicción de pertenencia al grupo | MANCOVA | |||||||||
| Vista | % | Chiapas | Tabasco | Veracruz | Total | Wilks’ λ | p | Efecto | Wilks’ λ | p<0.05 |
| Chiapas | 77.4 | 100 | 0.354 | <0.001 | LS | 0.926 | 0.607 | |||
| Laterala | Tabasco | 54.8 | 100 | Localidad | 0.471 | <0.001 | ||||
| Veracruz | 65.5 | 100 | ||||||||
| Chiapas | 77.4 | 100 | 0.478 | <0.001 | LS | 0.840 | 0.070 | |||
| Dorsalb | Tabasco | 38.7 | 100 | Localidad | 0.546 | <0.001 | ||||
| Veracruz | 48.3 | 100 | ||||||||
| Chiapas | 77.4 | 100 | 0.448 | <0.001 | LS | 0.922 | 0.560 | |||
| Ventralc | Tabasco | 48.4 | 100 | Localidad | 0.474 | <0.001 | ||||
| Veracruz | 55.2 | 100 | ||||||||
a 65.9% de la validación cruzada correctamente clasificada.
b 54.9% de la validación cruzada correctamente clasificada.
c 60.4% de la validación cruzada correctamente clasificada.
El porcentaje de varianza explicada por el primer CP para las localidades en M. cephalus fue de 45.4, 58 y 37.7% mientras que el segundo explicó 17.3, 18.4 y 23.7%, con una varianza total acumulada de 62.7, 76.5 y 61.4%, para la vista lateral, dorsal y ventral, respectivamente. El resultado del análisis discriminante por localidades fue de 66.7, 49 y 56.9% (Wilks’ λ = 0.316; 0.468 y 0.351 respectivamente con p < 0.001 y p < 0.05, este último en la vista dorsal) para la vista lateral, dorsal y ventral, respectivamente; con mejor porcentaje de discriminación las localidades Veracruz y Chiapas (Tabla 4). Los MANCOVA confirman diferencias significativas en la forma del urohial de todas las vistas entre localidades, mientras que la longitud estándar no resultó ser significativa para la forma con excepción de la vista ventral. El porcentaje de la forma explicada por la longitud estándar fue 7.28, 6.99 y 5.57% para la vista lateral, dorsal y ventral, respectivamente.
Tabla 4 Resultados de la discriminación con validación cruzada entre las localidades de Mugil cephalus. LS = Longitud standard (mm).
| Predicción de pertenencia al grupo | MANCOVA | |||||||||
| Vista | % | Chiapas | Tabasco | Veracruz | Total | Wilks’ λ | p | Efecto | Wilks’ λ | P<0.05 |
| Laterala | Chiapas | 50 | 100 | 0.316 | <0.001 | LS | 0.766 | 0.180 | ||
| Tabasco | 52.9 | 100 | Localidad | 0.450 | 0.005 | |||||
| Veracruz | 80.8 | 100 | ||||||||
| Dorsalb | Chiapas | 50 | 100 | 0.468 | <0.05 | LS | 0.782 | 0.227 | ||
| Tabasco | 41.2 | 100 | Localidad | 0.472 | 0.008 | |||||
| Veracruz | 53.8 | 100 | ||||||||
| Ventralc | Chiapas | 75 | 100 | 0.351 | <0.001 | LS | 0.688 | 0.042 | ||
| Tabasco | 47.1 | 100 | Localidad | 0.380 | <0.001 | |||||
| Veracruz | 57.7 | 100 | ||||||||
a 66.7% de la validación cruzada correctamente clasificada.
b 49.0% de la validación cruzada correctamente clasificada.
c 56.9% de la validación cruzada correctamente clasificada.
DISCUSIÓN Y CONCLUSIÓN
La forma del urohial permitió discriminar entre especies, donde la vista dorsal fue la que mostró mayor porcentaje de discriminación (96.5%) sin influencia de la talla. Este resultado es relevante considerando que, cuando se quiere discriminar entre especies, las diferencias en la talla suelen ser notorias. Para las especies en este estudio, en las costas del Atlántico, Mugil cephalus es la de mayor talla con un valor máximo de 120 cm de longitud estándar, mientras que M. curema alcanza un máximo 91 cm (Harrison, 2002).
Nuestros resultados son similares a los encontrados con métodos que utilizan escamas y en los cuales se han constatado altos porcentajes de discriminación (98.1%) en identificación de las mismas especies analizadas en este estudio (Ibañez et al., 2007; Ibáñez et al., 2009). Asimismo, los resultados se comparan razonablemente con tasas entre 70% y 86% registradas en otros estudios que utilizan otolitos para el mismo fin (Leguá et al., 2013; Tuset et al., 2006), así como con el patrón de escamación rómbica que ha mostrado porcentajes de clasificación entre 70 y 76% en la discriminación entre especies (Ibáñez et al., 2020).
A diferencia de la discriminación entre especies, utilizando la forma de escamas y otolitos se han reportado valores de discriminación entre localidades generalmente superiores al 70% (Ibáñez et al., 2007; Ibáñez et al., 2012; Ibáñez et al., 2017). En este estudio la forma del hueso urohial mostró menores porcentajes de discriminación entre localidades (66.3% como promedio de los valores más altos en ambas especies). La vista del urohial que resultó más exitosa en la discriminación de localidades fue la vista lateral seguida por la ventral y por último la dorsal, lo cual se atribuye posiblemente a los hábitos alimenticios, pues se considera una estructura importante en el mecanismo de apertura y cierre de la boca en los peces (Jawad et al., 2016), no obstante, debido a la cercanía entre localidades del Golfo de México y posible entrecruzamiento no hay variación morfológica significativa entre las localidades analizadas. Cabe destacar que en términos genéticos se han reportado que las diferencias entre especies y entre localidades pueden deberse a que Mugil curema ha mostrado estructura en los estudios genéticos poblacionales (Pacheco-Almanzar et al., 2017; Pacheco-Almanzar, 2019) además de mostrar comportamiento filopátrico tanto en el Pacífico como en el Golfo de México (Colín, 2020) a diferencia de M. cephalus que muestra una población panmíctica en el Golfo de México (Colín et al., 2020).
Gauldie (1988) explica la estabilidad intraespecífica como aquello que obedece a una limitación biológica específica debido a su función, en este caso está claramente asociada con una mayor discriminación interespecífica que con la diferenciación entre localidades debido a la permanencia de sus características que lo colocan como un buen marcador interespecífico. Para algunos investigadores, el sistema óseo constituye el fundamento sobre el cual se apoya la definición de los grandes grupos y la discusión de afinidades y diferencias entre los mismos (Greenwood et al., 1966). De esta manera, la osteología de peces ha permitido aportar elementos para la descripción y diferenciación de especies (Campello & Bemvenuti, 2002). Consecuentemente, fue posible determinar la utilidad de la forma del urohial como un método sencillo para discriminar entre las especies Mugil curema y M. cephalus, este resultado es relevante considerando que esta familia presenta una morfología muy similar con amplia problemática para distinguir especies. En este sentido es que se sugiere contar con un catálogo de referencia de urohiales para diferentes especies de la familia Mugilidae.

