











 nova página do texto(beta)
nova página do texto(beta) Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO 
 Permalink
PermalinkINTRODUCCIÓN
La calidad del agua en el litoral Pacífico mexicano continúa cambiando por efecto del desarrollo urbano, turístico e industrial (Lara-Lara et al., 2008). El aumento o disminución de los límites naturales en la concentración de compuestos químicos, como oxígeno disuelto, nitratos, fosfatos y metales en la columna de agua, pueden afectar la calidad de los ecosistemas (Botello et al., 2014) debido a tres factores principales: 1) Toxicidad 2) Persistencia y 3) Bioacumulación (Quintero-Rendón et al., 2010).
Las macroalgas se consideran indicadores biológicos, porque presentan ciclos de vida cortos, toman los nutrientes directamente de la columna de agua, son sensibles a contaminantes, son fáciles de observar y de recolectar (Ospina-Álvarez & Peña-Salamanca, 2004; Robledo & Friele-Pelegrín, 2014). Además, reflejan la magnitud del estrés con cambios en el contenido de minerales, así como, las características del hábitat y el grado de exposición a un estresor (Páez-Osuna et al., 2000). La selección de las mejores especies indicadoras, del vasto acervo natural, se obtiene evaluando la sensibilidad o tolerancia (intervalo) a la concentración de elementos en el agua de mar, en el que viven los distintos taxa; y que pueden variar naturalmente y por la contaminación antropogénica (De la Lanza-Espino, 2000).
Se ha observado, que no todas las macroalgas sobreviven en áreas con fuertes actividades antrópicas (Godínez, 2000); es decir, estas especies se distribuyen en intervalos estrechos respecto a las concentraciones de los elementos químicos disueltos en el agua. Por lo tanto, se esperaría que, ante el aumento en las concentraciones de compuestos disueltos, se produzcan cambios a nivel comunitario en la taxocenosis, i.e., florísticos. Dicho cambio, en la asociación de especies puede constituirse como bioindicador de contaminación; para establecer este indicador de cambio se requiere que la taxonomía y la taxocenosis de una comunidad, en un estado inicial de un sistema costero de interés, sean bien conocidas. En el área de Baja California Sur, México, Siqueiros-Beltrones & Riosmena Rodríguez (1995) identificaron 102 especies de macroalgas. Adicionalmente, para la zona se conocen otros aspectos comunitarios, como la variabilidad espaciotemporal de especies conspicuas (Cruz-Ayala, 1996), las especies con mayor producción de biomasa (Fajardo, 1993) y los cambios en contenidos de minerales ante los procesos de contaminación (Páez-Osuna et al., 2000; Hernández-Almaraz et al., 2014, 2016).
Los efectos de variables ambientales en la distribución de las especies pueden estudiarse a través del programa ModestR® que permite descargar, importar y depurar registros georreferenciados de las especies en bases de datos a nivel global (García-Rosello et al., 2014; GBIF, 2018). Las especies con distribución en un amplio intervalo de concentración de un compuesto en el agua indican especies tolerantes ante cambios de la variable. Mientras que, las especies presentes en intervalos estrechos revelan taxa sensibles al cambio (Quintero-Rendón et al., 2010) y muestran un estado particular del sistema costero. Teóricamente, un cambio extraordinario de los límites (máximos y mínimos) que se establecieron a partir de la distribución a nivel mundial, comprometería la supervivencia de cierta especie en el sistema costero, donde se presente dicha condición (Godínez, 2000).
En el presente trabajo, primero se analiza cada variable donde se establecen las concentraciones críticas para la supervivencia a nivel de especie y a nivel de clases. Y, luego, se plantea un modelo hipotético de variación taxocenótica por estados de calidad de sistemas costeros, para Baja California Sur ante escenarios de eutrofización.
MATERIALES Y MÉTODOS
Las variables bióticas o la taxocenosis conformada por 102 especies de macroalgas publicadas en GBIF (Siqueiros-Beltrones et al., 2017) para muestreos de 1994 y 1995 de las bahías La Ventana y La Paz en Baja California Sur (Siqueiros-Beltrones & Riosmena Rodríguez, 1995) fueron actualizadas taxonómicamente mediante la base de datos ALGABASE (Guiry & Guiry, 2018) y utilizando el Integrated Taxonomic Information System (ITIS, 2018). Sin embargo, dado que el programa ModestR® utiliza esta última base para obtener los registros, los nombres derivados se ajustan a esta e incluyen las sinonimias registradas.
La lista taxonómica lograda se consideró como “taxocenosis modelo” de un sistema costero en condiciones iniciales, sin perturbación antropogénica; ésta se incorporó a la aplicación Data Manager, con el fin de obtener las presencias georreferenciadas para todo el planeta en el ambiente marino, de especímenes preservados y/o citados en la literatura de cada especie (García-Rosello et al., 2014; GBIF, 2018). Se obtuvieron 71 especies, con un número robusto de datos de presencia, para elaborar un modelo de distribución de Kernel y realizar una comparación estadística con promedios y desviaciones estándar.
La capa obtenida para cada especie se depuró (García-Rosello et al., 2014) y se superpuso a las capas de variables abióticas, que se obtuvieron del promedio de un conjunto de campos climatológicos, analizados objetivamente (cuadrícula 1 °) y campos estadísticos asociados de datos de perfil oceanográfico (5 °, 1 ° y 0.25 °) a niveles de profundidad estándar para períodos de composición anuales, estacionales y mensuales para el Océano Mundial del World Ocean Atlas 2013 versión 2 (WOA13 V2). Aquí se analizaron, la temperatura superficial del mar (°C) (Locarnini et al., 2013), salinidad, oxígeno disuelto (ml l-1), silicatos (µmol l-1), fosfatos (µmol l-1), nitratos (µmol l-1) y materia orgánica disuelta (µmol l-1) en agua de mar de 2005 a 2012 (García et al., 2013a, 2013b; Zweng et al., 2013); este análisis se sobrepuso con la capa obtenida del registro de presencias y con el modelo de distribución de Kernel de cada especie, creando y exportando esto a una base de datos cruzada (Guisande, 2013; García-Rosello et al., 2014) donde cada registro de presencia de la especie le corresponde un promedio para cada valor fisicoquímico y de nutrientes.
Se determinó el intervalo de tolerancia a partir de la amplitud, el método más simple para estimar la dispersión de los datos, es decir, la diferencia entre el mínimo y el máximo de la serie de datos obtenidos. Sin embargo, para evitar muestras sobreestimadas o subestimadas (valores atípicos) se tiene en cuenta aquí, la desviación típica o estándar que mide la variabilidad de los datos respecto a la media en la muestra (Guisande et al., 2006) o tolerancia de la especie. Para visualizar esto, las especies se ordenaron en el eje X, en función del incremento de las concentraciones del compuesto químico en el agua de mar. En el eje Y, se establecieron los valores críticos en donde se perdería el 95% de las especies que conforman la taxocenosis de Baja California Sur. Es así como los valores óptimos que se establecieron corresponden al intercuartil del recorrido del intervalo definido, o donde se agrupa el mayor número de presencia de cada especie.
Se planteó el escenario actual de Baja California Sur con promedios de los últimos seis años, de temperatura superficial del agua de mar (2012-2017) (https://seatemperature.info/es/mexico/la-paz-temperatura-del-agua-del-mar.html), lo cual varía según la época del año, invierno (noviembre a febrero) y verano (marzo-octubre) entre 17 y 28 °C (Fig. 2) y se sobrepuso a los intervalos de tolerancia a la temperatura superficial de agua de mar de cada especie (eje x1), ordenadas en función del incremento de esta variable.

Figura 2 Intervalos de temperatura (°C) superficial en el agua de mar donde se
distribuyen las especies de macroalgas  = promedio
= promedio
 =
desviación estándar
=
desviación estándar  = valores máximos y
= valores máximos y
 =
valores mínimos mensuales de 6 años (2012 a 2017) de temperatura del
agua de BCS, México. Posibles escenarios futuros
=
valores mínimos mensuales de 6 años (2012 a 2017) de temperatura del
agua de BCS, México. Posibles escenarios futuros  -2°C y/o
-2°C y/o
 +
1°C en el promedio del agua de mar en BCS.
+
1°C en el promedio del agua de mar en BCS.
Además, se hipotetizan posibles escenarios futuros que refieren el descenso en la temperatura promedio en -2°C, así como, el aumento del promedio en 1°C del agua superficial; para ello, se establecieron tolerancia mínima a 15 °C y máxima de 29 °C en el agua de mar de sistemas costeros.
Para determinar si es posible discriminar si las clases de macroalgas crecen en diferentes condiciones fisicoquímicas y de concentración de nutrientes del agua, se llevó a cabo un análisis de clasificación, discriminante, usando la aplicación StatR del programa Rwizard. Esta técnica estadística multivariante caracteriza clases de elementos a partir de una clasificación inicial en función de los valores observados de un conjunto de variables, como en este caso de fisicoquímicos y nutrientes. Así, este asigna en qué clase debe estar cada elemento a partir de la proximidad (o similitud) y buscando la variable que hace máxima la separación entre grupos, lo que equivale a minimizar la variabilidad, usando el criterio de Bayes: cada elemento se asigna a la clase, para la que tiene mayor probabilidad de pertenecer, condicionada por los valores que toman las funciones discriminantes (Guisande et al., 2016).
Las hipótesis de normalidad de residuos y homogeneidad de varianzas (M de Box) se utilizan para el cálculo de las probabilidades de asignación de los elementos a las distintas clases; no obstante, estas son muy sensibles a pequeñas desviaciones y, con datos reales, prácticamente siempre nos dirán que no se cumplen. Por tanto, el discriminante es razonablemente robusto, frente a un incumplimiento parcial de estas hipótesis y ese incumplimiento no es crítico, si el porcentaje de acierto en la clasificación (especialmente en la validación cruzada) es elevado, no solamente en el conjunto de los elementos, sino también en cada uno de los grupos. No es necesario ni conveniente verificar las hipótesis, salvo que tengamos una clara evidencia de que se incumplen de forma manifiesta. Si el porcentaje de acierto es bajo, ello podría deberse al incumplimiento de las hipótesis (Guisande et al., 2016).
Como referencia visual, se generó un mapa de calor que muestra con distintos colores y tonos, la intensidad de la relación entre las variables estudiadas entre filas (bióticas: especie) y columnas (abióticas: fisicoquímicos y nutrientes en el agua), clasificando de acuerdo con el valor de correlación entre las variables, uniendo en primer lugar, los dos elementos más próximos entre sí y elaborando dendrogramas marginales para las filas y otro para las columnas. Los colores indican la intensidad de la relación o los valores de la variable representada, desde el blanco (máxima relación o valor más alto), pasando por el amarillo y naranja, hasta el rojo intenso del valor mínimo (Guisande et al., 2016).
RESULTADOS
En este trabajo se establecieron los intervalos de tolerancia (promedios y la desviación estándar) de las especies de macroalgas, que fueron registradas para Baja California Sur en 1995, a variables fisicoquímicos del agua de mar (Tabla 1, se incluyen las autoridades de cada taxón). Asimismo, se establecieron intervalos y valores críticos para cada Clase y para la taxocenosis de macroalgas de Baja California Sur, lo que se discute a continuación por variable abiótica estudiada.
Tabla 1 Intervalos de tolerancia (promedio y desviación estándar) de las especies de macroalgas a diversas concentraciones de nutrientes y condiciones fisicoquímicas en el océano mundial.
| Especies | Fisicoquímicos | Nutrientes | ||||||||||||||||||||||
| Temperatura °C | pH | OD ml l-1 | Salinidad | Silicato µmol l-1 | Fosfatos µmol l-1 | Nitrato µmol l-1 | MOD µmol l-1 | |||||||||||||||||
| Ahnfeltia plicata (Hudson) Fries | 10,72 | ± | 2,52 | 8,19 | ± | 0,09 | 6,71 | ± | 0,57 | 27,48 | ± | 7,85 | 5,24 | ± | 4,08 | 0,44 | ± | 0,21 | 4,09 | ± | 2,78 | 3,08 | ± | 1,18 |
| Amphiroa beauvoisii J.V.Lamouroux | 24,79 | ± | 3,42 | 8,23 | ± | 0,06 | 4,95 | ± | 0,32 | 34,91 | ± | 0,80 | 3,73 | ± | 3,22 | 0,37 | ± | 0,27 | 1,42 | ± | 0,89 | 3,62 | ± | 1,07 |
| Amphiroa misakiensis Yendo | 27,21 | ± | 2,18 | 8,25 | ± | 0,02 | 4,78 | ± | 0,17 | 34,45 | ± | 0,35 | 3,69 | ± | 2,76 | 0,42 | ± | 0,23 | 1,58 | ± | 0,61 | 3,59 | ± | 0,88 |
| Amphiroa rigida J.V. Lamouroux | 21,61 | ± | 4,15 | 8,21 | ± | 0,09 | 5,15 | ± | 0,34 | 36,81 | ± | 1,24 | 2,15 | ± | 1,51 | 0,21 | ± | 0,16 | 1,19 | ± | 0,43 | 4,21 | ± | 1,31 |
| Amphiroa valonioides Yendo | 25,85 | ± | 2,78 | 8,25 | ± | 0,02 | 4,86 | ± | 0,27 | 34,63 | ± | 0,54 | 4,67 | ± | 4,10 | 0,43 | ± | 0,33 | 1,62 | ± | 1,10 | 3,33 | ± | 1,68 |
| Antithamnionella elegans (Berthold) J.H. Price et D.M. John | 21,02 | ± | 2,31 | 8,23 | ± | 0,02 | 5,28 | ± | 0,18 | 36,62 | ± | 1,09 | 5,03 | ± | 5,57 | 0,39 | ± | 0,44 | 1,88 | ± | 1,22 | 4,70 | ± | 0,94 |
| Antithamnionella glandulifera (Kylin) E.M. Wollaston | 15,09 | ± | 4,87 | 8,09 | ± | 0,18 | 5,55 | ± | 0,57 | 32,98 | ± | 3,64 | 17,81 | ± | 18,45 | 0,85 | ± | 0,77 | 4,13 | ± | 3,70 | 3,69 | ± | 0,88 |
| Asparagopsis taxiformis (Delile) Trevisan | 24,88 | ± | 3,33 | 8,25 | ± | 0,06 | 4,88 | ± | 0,27 | 35,41 | ± | 0,88 | 2,38 | ± | 1,22 | 0,20 | ± | 0,11 | 0,96 | ± | 0,62 | 2,67 | ± | 1,54 |
| Botryocladia neushulii E.Y. Dawson | 17,08 | ± | 0,17 | 8,22 | ± | 0,00 | 5,80 | ± | 0,01 | 33,51 | ± | 0,01 | 2,53 | ± | 0,00 | 0,36 | ± | 0,00 | 0,06 | ± | 0,00 | 3,59 | ± | 0,05 |
| Bryopsis pennata J.V. Lamouroux | 26,48 | ± | 2,31 | 8,20 | ± | 0,09 | 4,78 | ± | 0,27 | 35,33 | ± | 0,85 | 3,07 | ± | 2,17 | 0,24 | ± | 0,19 | 1,31 | ± | 0,71 | 2,82 | ± | 1,52 |
| Caulerpa sertularioides (S.G. Gmelin) M. Howe | 26,79 | ± | 1,76 | 8,22 | ± | 0,11 | 4,74 | ± | 0,17 | 35,39 | ± | 0,88 | 2,82 | ± | 1,35 | 0,21 | ± | 0,17 | 0,94 | ± | 0,51 | 3,57 | ± | 1,44 |
| Centroceras clavulatum (C. Agardh) Montagne | 22,39 | ± | 4,99 | 8,23 | ± | 0,08 | 5,10 | ± | 0,45 | 35,35 | ± | 0,90 | 2,63 | ± | 1,62 | 0,23 | ± | 0,18 | 1,56 | ± | 1,55 | 3,37 | ± | 1,45 |
| Ceramium caudatum Setchell et N.L. Gardner | 24,04 | ± | 3,77 | 8,23 | ± | 0,03 | 5,02 | ± | 0,30 | 34,50 | ± | 0,64 | 5,08 | ± | 3,50 | 0,57 | ± | 0,26 | 1,65 | ± | 1,09 | 4,02 | ± | 1,29 |
| Ceramium fimbriatum Setchell et N.L. Gardner | 25,36 | ± | 0,84 | 8,25 | ± | 0,01 | 4,82 | ± | 0,07 | 34,96 | ± | 0,23 | 3,09 | ± | 2,46 | 0,20 | ± | 0,20 | 1,51 | ± | 0,73 | 2,38 | ± | 1,33 |
| Ceramium flaccidum (Harvey ex Kützing) Ardissone | 23,69 | ± | 5,39 | 8,26 | ± | 0,06 | 5,00 | ± | 0,46 | 35,02 | ± | 0,71 | 2,35 | ± | 1,40 | 0,23 | ± | 0,13 | 1,65 | ± | 1,48 | 2,77 | ± | 1,42 |
| Ceramium paniculatum Okamura | 23,72 | ± | 5,70 | 8,25 | ± | 0,01 | 5,14 | ± | 0,55 | 33,94 | ± | 0,41 | 4,30 | ± | 2,42 | 0,27 | ± | 0,05 | 1,49 | ± | 0,09 | 3,50 | ± | 0,51 |
| Ceramium procumbens Setchell et N.L. Gardner | 25,34 | ± | 0,00 | 8,28 | ± | 0,00 | 4,91 | ± | 0,00 | 34,63 | ± | 0,00 | 4,50 | ± | 0,00 | 0,53 | ± | 0,00 | 1,27 | ± | 0,00 | 3,98 | ± | 0,00 |
| Chaetomorpha linum (O.F. Müller) Kützing | 14,52 | ± | 5,55 | 8,18 | ± | 0,07 | 6,10 | ± | 0,75 | 32,38 | ± | 6,33 | 3,75 | ± | 2,97 | 0,38 | ± | 0,23 | 4,34 | ± | 3,88 | 4,01 | ± | 1,41 |
| Champia parvula (C. Agardh) Harvey | 22,98 | ± | 5,70 | 8,22 | ± | 0,11 | 5,09 | ± | 0,56 | 34,83 | ± | 1,67 | 2,67 | ± | 1,68 | 0,24 | ± | 0,19 | 1,49 | ± | 1,43 | 3,06 | ± | 1,48 |
| Chondria acrorhizophora Setchell et N.L. Gardner | 17,31 | ± | 0,46 | 8,22 | ± | 0,01 | 5,80 | ± | 0,04 | 33,50 | ± | 0,01 | 2,62 | ± | 0,10 | 0,36 | ± | 0,01 | 0,17 | ± | 0,11 | 3,34 | ± | 0,25 |
| Cladophora columbiana Collins | 14,16 | ± | 3,64 | 8,19 | ± | 0,06 | 6,16 | ± | 0,53 | 32,83 | ± | 1,56 | 8,99 | ± | 8,16 | 0,67 | ± | 0,26 | 2,96 | ± | 2,11 | 3,25 | ± | 0,64 |
| Cladophora hesperia Setchell et N.L. Gardner | 24,18 | ± | 0,78 | 8,25 | ± | 0,02 | 5,13 | ± | 0,21 | 35,06 | ± | 0,33 | 9,69 | ± | 4,15 | 0,88 | ± | 0,26 | 2,73 | ± | 1,08 | 4,15 | ± | 0,70 |
| Cladophora microcladioides Collins | 24,76 | ± | 4,94 | 8,24 | ± | 0,02 | 5,06 | ± | 0,50 | 34,38 | ± | 0,59 | 4,87 | ± | 3,77 | 0,52 | ± | 0,29 | 1,90 | ± | 1,10 | 3,55 | ± | 0,87 |
| Cladophoropsis gracillima E.Y. Dawson | 25,01 | ± | 1,93 | 8,25 | ± | 0,04 | 5,14 | ± | 0,37 | 35,15 | ± | 0,47 | 9,17 | ± | 5,46 | 0,75 | ± | 0,44 | 2,36 | ± | 1,46 | 2,24 | ± | 1,18 |
| Codium cuneatum Setchell et N.L. Gardner | 23,01 | ± | 2,01 | 8,26 | ± | 0,01 | 5,15 | ± | 0,30 | 34,48 | ± | 0,53 | 5,68 | ± | 3,05 | 0,62 | ± | 0,22 | 1,60 | ± | 1,04 | 4,31 | ± | 1,22 |
| Codium setchellii N.L. Gardner | 9,28 | ± | 3,67 | 8,12 | ± | 0,10 | 6,62 | ± | 0,52 | 31,90 | ± | 1,30 | 20,36 | ± | 9,56 | 1,17 | ± | 0,42 | 7,52 | ± | 3,54 | 2,67 | ± | 0,83 |
| Codium simulans Setchell et N.L. Gardner | 22,74 | ± | 2,42 | 8,24 | ± | 0,03 | 5,17 | ± | 0,26 | 34,72 | ± | 0,63 | 8,34 | ± | 4,86 | 0,79 | ± | 0,32 | 2,29 | ± | 1,51 | 3,71 | ± | 0,79 |
| Colpomenia ramosa W.R. Taylor | 23,94 | ± | 0,91 | 8,24 | ± | 0,02 | 5,17 | ± | 0,20 | 35,12 | ± | 0,26 | 11,12 | ± | 3,89 | 0,97 | ± | 0,25 | 3,10 | ± | 1,07 | 4,27 | ± | 0,92 |
| Corallina vancouveriensis Yendo | 13,46 | ± | 5,63 | 8,12 | ± | 0,12 | 6,16 | ± | 0,78 | 31,74 | ± | 2,66 | 16,82 | ± | 12,12 | 0,95 | ± | 0,51 | 4,73 | ± | 3,15 | 3,27 | ± | 1,12 |
| Dasya sinicola (Setchell et N.L. Gardner) E.Y. Dawson | 22,77 | ± | 3,98 | 8,25 | ± | 0,02 | 5,15 | ± | 0,42 | 34,36 | ± | 0,65 | 4,93 | ± | 3,18 | 0,54 | ± | 0,24 | 1,35 | ± | 1,11 | 4,00 | ± | 1,23 |
| Derbesia marina (Lyngbye) Solier | 13,65 | ± | 6,89 | 8,21 | ± | 0,06 | 6,27 | ± | 0,90 | 32,87 | ± | 3,72 | 4,23 | ± | 4,55 | 0,43 | ± | 0,25 | 4,00 | ± | 3,20 | 3,21 | ± | 1,08 |
| Dictyopteris delicatula J.V. Lamouroux | 26,75 | ± | 2,05 | 8,16 | ± | 0,10 | 4,76 | ± | 0,22 | 35,79 | ± | 0,99 | 2,75 | ± | 1,10 | 0,19 | ± | 0,10 | 1,17 | ± | 1,09 | 4,06 | ± | 1,54 |
| Dictyosphaeria cavernosa (Forsskål) Børgesen | 27,11 | ± | 1,83 | 8,23 | ± | 0,09 | 4,67 | ± | 0,17 | 35,15 | ± | 0,80 | 2,66 | ± | 0,93 | 0,15 | ± | 0,08 | 0,90 | ± | 0,61 | 2,84 | ± | 1,66 |
| Dictyota crenulata J. Agardh | 26,40 | ± | 1,60 | 8,26 | ± | 0,03 | 4,82 | ± | 0,11 | 34,68 | ± | 0,57 | 3,62 | ± | 1,74 | 0,40 | ± | 0,19 | 1,23 | ± | 0,45 | 4,04 | ± | 0,80 |
| Dictyota dichotoma (Hudson) J.V. Lamouroux | 14,35 | ± | 3,75 | 8,22 | ± | 0,05 | 5,91 | ± | 0,44 | 35,20 | ± | 1,00 | 2,36 | ± | 1,41 | 0,31 | ± | 0,18 | 3,72 | ± | 3,02 | 3,90 | ± | 1,04 |
| Dictyota flabellata (Collins) Setchell et N.L. Gardner | 22,42 | ± | 3,29 | 8,24 | ± | 0,03 | 5,24 | ± | 0,37 | 34,51 | ± | 0,69 | 6,43 | ± | 4,25 | 0,65 | ± | 0,28 | 1,70 | ± | 1,31 | 3,83 | ± | 0,86 |
| Digenea simplex (Wulfen) C. Agardh | 23,62 | ± | 3,62 | 8,23 | ± | 0,08 | 5,02 | ± | 0,35 | 36,43 | ± | 1,11 | 2,37 | ± | 1,64 | 0,19 | ± | 0,16 | 0,89 | ± | 0,42 | 4,52 | ± | 1,30 |
| Ectocarpus acutus Setchell et N.L. Gardner | 10,87 | ± | 3,94 | 8,03 | ± | 0,14 | 6,29 | ± | 0,59 | 30,51 | ± | 1,76 | 18,73 | ± | 13,46 | 1,01 | ± | 0,54 | 4,84 | ± | 2,99 | 3,81 | ± | 1,30 |
| Fosliella paschalis (Me. Lemoine) Setchell et N.L. Gardner | 24,61 | ± | 2,48 | 8,26 | ± | 0,01 | 4,92 | ± | 0,24 | 34,81 | ± | 0,47 | 6,36 | ± | 2,07 | 0,69 | ± | 0,18 | 2,00 | ± | 0,70 | 4,85 | ± | 1,21 |
| Gelidiella acerosa (Forsskål) Feldmann et Hamel | 27,28 | ± | 1,57 | 8,19 | ± | 0,11 | 4,66 | ± | 0,17 | 35,69 | ± | 1,09 | 2,97 | ± | 1,06 | 0,18 | ± | 0,11 | 0,89 | ± | 0,43 | 3,89 | ± | 2,00 |
| Gracilaria crispata Setchell et N.L. Gardner | 26,26 | ± | 2,02 | 8,26 | ± | 0,02 | 4,83 | ± | 0,16 | 34,58 | ± | 0,43 | 4,95 | ± | 3,04 | 0,54 | ± | 0,23 | 1,71 | ± | 0,75 | 4,45 | ± | 1,29 |
| Gracilaria pachydermatica Setchell et N.L. Gardner | 24,48 | ± | 1,02 | 8,26 | ± | 0,02 | 5,05 | ± | 0,22 | 34,88 | ± | 0,35 | 7,87 | ± | 4,26 | 0,75 | ± | 0,27 | 2,20 | ± | 1,10 | 4,10 | ± | 0,87 |
| Gracilaria spinigera E.Y. Dawson | 21,29 | ± | 7,32 | 8,19 | ± | 0,08 | 5,23 | ± | 0,71 | 34,04 | ± | 1,31 | 4,79 | ± | 2,21 | 0,54 | ± | 0,31 | 3,00 | ± | 3,34 | 3,45 | ± | 1,05 |
| Gracilaria subsecundata Setchell et N.L. Gardner | 23,67 | ± | 1,28 | 8,24 | ± | 0,02 | 5,08 | ± | 0,22 | 34,92 | ± | 0,53 | 9,55 | ± | 4,58 | 0,85 | ± | 0,27 | 2,67 | ± | 1,23 | 4,96 | ± | 0,95 |
| Gracilaria textorii (Suringar) Hariot | 22,85 | ± | 3,36 | 8,27 | ± | 0,11 | 5,10 | ± | 0,32 | 34,64 | ± | 0,97 | 6,19 | ± | 3,31 | 0,44 | ± | 0,31 | 1,45 | ± | 1,35 | 3,88 | ± | 1,14 |
| Gracilaria turgida E.Y. Dawson | 22,47 | ± | 2,97 | 8,25 | ± | 0,03 | 5,26 | ± | 0,40 | 34,40 | ± | 0,65 | 5,90 | ± | 4,27 | 0,60 | ± | 0,28 | 1,45 | ± | 1,37 | 3,84 | ± | 0,83 |
| Griffithsia pacifica Kylin | 14,12 | ± | 6,13 | 8,13 | ± | 0,13 | 6,12 | ± | 0,83 | 31,59 | ± | 2,34 | 19,82 | ± | 10,68 | 1,06 | ± | 0,49 | 4,74 | ± | 2,35 | 3,30 | ± | 1,19 |
| Halimeda discoidea Decaisne | 26,35 | ± | 1,80 | 8,24 | ± | 0,08 | 4,76 | ± | 0,16 | 35,28 | ± | 0,72 | 2,62 | ± | 1,35 | 0,21 | ± | 0,15 | 0,97 | ± | 0,60 | 2,58 | ± | 1,28 |
| Laurencia pacifica Kylin | 21,33 | ± | 3,50 | 8,25 | ± | 0,03 | 5,34 | ± | 0,39 | 34,11 | ± | 0,54 | 3,83 | ± | 1,20 | 0,48 | ± | 0,10 | 0,89 | ± | 0,56 | 3,66 | ± | 0,66 |
| Laurencia papillosa (C.Agardh) Bertoloni | 24,96 | ± | 3,82 | 8,22 | ± | 0,10 | 4,89 | ± | 0,36 | 36,03 | ± | 1,38 | 2,64 | ± | 1,40 | 0,16 | ± | 0,12 | 0,88 | ± | 0,55 | 4,19 | ± | 1,69 |
| Liagora californica Zeh | 24,72 | ± | 1,82 | 8,27 | ± | 0,02 | 4,99 | ± | 0,22 | 34,62 | ± | 0,33 | 4,86 | ± | 0,86 | 0,57 | ± | 0,08 | 1,37 | ± | 0,35 | 3,68 | ± | 0,51 |
| Lithophyllum imitans Foslie | 21,53 | ± | 5,44 | 8,18 | ± | 0,10 | 5,25 | ± | 0,44 | 34,41 | ± | 1,59 | 15,14 | ± | 8,30 | 1,08 | ± | 0,38 | 4,03 | ± | 1,80 | 4,64 | ± | 1,16 |
| Lithophyllum proboscideum (Foslie) Foslie | 23,09 | ± | 2,79 | 8,22 | ± | 0,03 | 5,17 | ± | 0,26 | 34,90 | ± | 0,65 | 10,86 | ± | 5,32 | 0,90 | ± | 0,33 | 2,87 | ± | 1,62 | 4,38 | ± | 1,51 |
| Lomentaria hakodatensis Yendo | 21,44 | ± | 5,65 | 8,22 | ± | 0,09 | 5,35 | ± | 0,71 | 33,53 | ± | 2,28 | 6,93 | ± | 8,93 | 0,33 | ± | 0,28 | 1,97 | ± | 1,68 | 3,12 | ± | 1,50 |
| Padina durvillaei Bory | 24,96 | ± | 3,75 | 8,23 | ± | 0,09 | 4,84 | ± | 0,30 | 35,58 | ± | 0,99 | 2,55 | ± | 1,69 | 0,18 | ± | 0,16 | 1,03 | ± | 0,72 | 3,20 | ± | 1,57 |
| Padina mexicana E.Y. Dawson | 26,19 | ± | 1,67 | 8,26 | ± | 0,02 | 4,85 | ± | 0,14 | 34,65 | ± | 0,40 | 5,56 | ± | 3,73 | 0,58 | ± | 0,26 | 1,80 | ± | 0,90 | 4,01 | ± | 0,92 |
| Porphyra thuretii Setchell et E.Y. Dawson | 20,35 | ± | 5,93 | 8,23 | ± | 0,05 | 5,48 | ± | 0,73 | 33,87 | ± | 1,56 | 9,17 | ± | 6,24 | 0,72 | ± | 0,24 | 2,38 | ± | 1,31 | 3,71 | ± | 0,69 |
| Prionitis cornea (Okamura) E.Y.Dawson | 18,19 | ± | 2,30 | 8,25 | ± | 0,01 | 5,67 | ± | 0,25 | 33,60 | ± | 0,39 | 6,35 | ± | 1,52 | 0,30 | ± | 0,10 | 1,35 | ± | 0,43 | 3,84 | ± | 0,77 |
| Pterocladia capillacea (S.G.Gmelin) Bornet in Bornet & Thuret | 15,89 | ± | 0,72 | 8,26 | ± | 0,01 | 5,65 | ± | 0,08 | 35,57 | ± | 0,12 | 0,97 | ± | 0,35 | 0,17 | ± | 0,05 | 0,76 | ± | 0,52 | 3,80 | ± | 0,77 |
| Ralfsia confusa Hollenberg | 26,92 | ± | 1,29 | 8,26 | ± | 0,01 | 4,76 | ± | 0,07 | 34,47 | ± | 0,40 | 4,14 | ± | 2,25 | 0,48 | ± | 0,20 | 1,55 | ± | 0,57 | 4,14 | ± | 1,03 |
| Rosenvingea intricata (J.Agardh) Børgesen | 26,14 | ± | 1,71 | 8,23 | ± | 0,13 | 4,78 | ± | 0,17 | 35,39 | ± | 0,67 | 2,75 | ± | 1,80 | 0,20 | ± | 0,20 | 0,92 | ± | 0,60 | 3,95 | ± | 1,73 |
| Sargassum lapazeanum Setchell et N.L. Gardner | 25,15 | ± | 0,09 | 8,27 | ± | 0,00 | 4,92 | ± | 0,01 | 34,63 | ± | 0,07 | 4,83 | ± | 0,33 | 0,56 | ± | 0,03 | 1,39 | ± | 0,11 | 3,97 | ± | 0,41 |
| Sphacelaria californica Setchell. et N.L. Gardner, N.L. | 20,76 | ± | 3,57 | 8,23 | ± | 0,03 | 5,45 | ± | 0,41 | 34,06 | ± | 0,66 | 5,01 | ± | 4,21 | 0,53 | ± | 0,27 | 1,14 | ± | 1,31 | 3,76 | ± | 0,98 |
| Spyridia filamentosa (Wulfen) Harvey | 21,19 | ± | 4,55 | 8,24 | ± | 0,07 | 5,25 | ± | 0,47 | 35,65 | ± | 1,44 | 2,48 | ± | 1,94 | 0,24 | ± | 0,20 | 1,52 | ± | 1,91 | 3,99 | ± | 1,51 |
| Ulva clathrata (Roth) C. Agardh | 18,17 | ± | 6,32 | 8,24 | ± | 0,08 | 5,71 | ± | 0,86 | 32,89 | ± | 5,35 | 4,43 | ± | 5,28 | 0,36 | ± | 0,28 | 2,22 | ± | 2,54 | 3,31 | ± | 1,49 |
| Ulva flexuosa Wulfen | 19,33 | ± | 7,61 | 8,20 | ± | 0,09 | 5,53 | ± | 0,86 | 33,61 | ± | 4,57 | 3,43 | ± | 2,89 | 0,32 | ± | 0,19 | 3,14 | ± | 3,31 | 3,36 | ± | 1,49 |
| Ulva rigida C. Agardh | 18,86 | ± | 4,45 | 8,21 | ± | 0,08 | 5,36 | ± | 0,60 | 35,79 | ± | 1,46 | 3,35 | ± | 3,66 | 0,39 | ± | 0,41 | 2,50 | ± | 2,89 | 3,65 | ± | 1,15 |
| Valoniopsis pachynema (G. Martens) Børgesen | 24,90 | ± | 2,55 | 8,28 | ± | 0,08 | 4,94 | ± | 0,31 | 35,19 | ± | 0,52 | 5,19 | ± | 4,10 | 0,40 | ± | 0,37 | 1,29 | ± | 1,28 | 3,45 | ± | 1,54 |
| Veleroa subulata E.Y. Dawson | 26,14 | ± | 5,03 | 8,24 | ± | 0,01 | 4,91 | ± | 0,47 | 34,06 | ± | 0,32 | 2,16 | ± | 0,14 | 0,29 | ± | 0,05 | 1,14 | ± | 0,62 | 3,24 | ± | 0,37 |
Temperatura. Las especies de macroalgas de BCS agrupadas por Clase muestran que las Florideophyceae y Ulvophyceae se distribuyen en un intervalo más amplio (desviación estándar; entre 0 a 34 °C, Fig. 1); mientras que, las especies de Bangiophyceae (11 a 25 °C) y Phaeophyceae (8 a 26 °C) se restringen a intervalos estrechos. Sin embargo, el promedio de temperatura en la que se encuentran las algas pardas (Phaeophyceae) (15 °C) revela que estas se desarrollan en aguas más frías que otras clases. La temperatura superficial del agua de mar “óptima” (intercuartil del intervalo) para macroalgas se encuentra entre 12 y 27 °C.

Figura 1 Intervalo de la tolerancia de las clases de macroalgas a la
temperatura superficial (°C) del agua de mar — = promedio,
 =
intercuartil,
=
intercuartil,  = desviación
estándar, ° = atípicos,
= desviación
estándar, ° = atípicos,  = óptimos, --- =
críticos.
= óptimos, --- =
críticos.
Oxígeno disuelto. El promedio de OD en el que habitan las Phaeophyceae (5.6 ml l-1) es significativamente mayor a las demás clases. Mientras que, Florideophyceae (4.1 a 8 ml l-1) y Ulvophyceae (4.1 a 7.6 ml l-1) toleran intervalos más amplios de concentración de oxígeno en el agua, Phaeophyceae (4.2 a 7.1 ml l-1) y Bangiophyceae (5 a 6.9 ml l-1) se distribuyen en intervalos más estrechos (desviación estándar; Fig. 3).

Figura 3 Intervalos de tolerancia de macroalgas a la concentración de oxígeno
disuelto (ml l-1) en el agua de mar — = promedio,
=
intercuartil, = desviación
estándar, ° = valores atípicos, = concentraciones
óptimas.
Las especies evaluadas prosperan en concentraciones de oxígeno mínimas de 4.6, a máximas a 7.4 ml l-1, es decir, aguas oxigenadas. Sin embargo, el óptimo o intervalo de distribución del mayor número de especies se encuentra entre los 4.8 y 6.2 ml l-1, siendo el valor máximo de tolerancia para la taxocenosis el de 7.5 ml l-1 (Fig. 4).
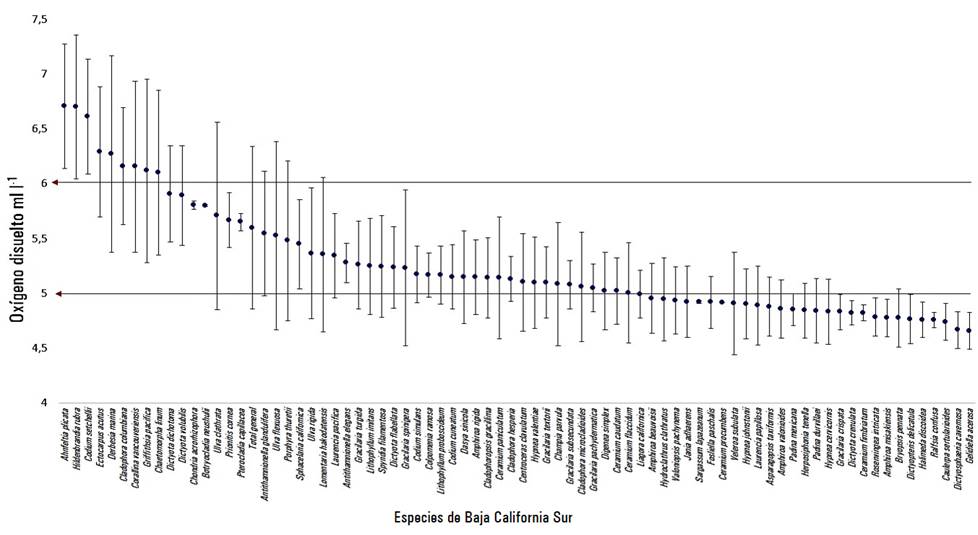
Figura 4 Intervalos de tolerancia de macroalgas a la concentración de oxígeno
disuelto (ml l-1) en el agua de mar  = promedio,
= promedio,
 =
desviación estándar (2005 -2012)
=
desviación estándar (2005 -2012)  = concentraciones
críticas para cambios en la taxocenosis de Baja California Sur.
= concentraciones
críticas para cambios en la taxocenosis de Baja California Sur.

Figura 5 Intervalos de tolerancia de las especies de macroalgas al pH del agua
de mar
= pH promedio en agua de mar, = desviación
estándar,  = concentraciones óptimas, = línea de tendencia, --- =
concentraciones críticas para la taxocenosis de Baja California
Sur.
= concentraciones óptimas, = línea de tendencia, --- =
concentraciones críticas para la taxocenosis de Baja California
Sur.
pH. Las macroalgas estudiadas se desarrollan en aguas con valores de pH entre 7.88 y 8.37. El intervalo donde se desarrollarían el mayor número de especies es de 8.1 a 8.3 (línea de tendencia) (Fig. 5).
Materia orgánica disuelta. Las especies de macroalgas estudiadas para BCS se distribuyen en el planeta en aguas de sistemas costeros con un intervalo de concentración de 1.1 a 7.2 µmol 1-1 de materia orgánica disuelta en el agua. Los valores donde se presenta el mayor número de registros se establecieron entre 2 y 5 µmol 1-1 (Fig. 6).

Figura 6 Intervalos de tolerancia de macroalgas a la concentración de materia
orgánica disuelta (µmol l-1 1 MOD) en agua de mar
 =
promedio,
=
promedio,  = desviación estándar, …… = línea de tendencia,
=
concentraciones óptimas.
= desviación estándar, …… = línea de tendencia,
=
concentraciones óptimas.
Nitratos. Las macroalgas se desarrollan en intervalos de concentraciones de 0 a 13 µmol l-1 de nitrato en la columna de agua con óptimos entre 0.5 y 5.2 µmol l-1; las Phaeophyceae se presentan a mayor concentración promedio de este nutriente y mayor tolerancia con amplitudes de 13 µmol l-1; no obstante, Florideophyceae y Ulvophyceae, aunque presentan valores atípicos en concentraciones mayores a 20 µmol l-1, el mayor número de datos se distribuye en un estrecho intervalo con máximos de desviación estándar de 4 y 5 µmol l-1 respectivamente (Fig. 7).

Figura 7 Diagrama de caja del intervalo de tolerancia de la concentración de
nitrato (NO3 mol l-1), — = promedio del agua de
mar, =
intercuartil, = desviación
estandar, ° = residuos,  = óptimos, --- =
críticos para supervivencia de macroalgas de B.C.S., México, agrupadas
por Clase.
= óptimos, --- =
críticos para supervivencia de macroalgas de B.C.S., México, agrupadas
por Clase.
Una concentración superior a 3 µmol l-1 de NO3 es crítica para la mayor parte de las especies, sin embargo, será óptima para, por lo menos, 18 de ellas. Por el contrario, el aumento a promedios de 5 µmol l-1 de NO3 constituye un valor crítico para todas las especies de macroalgas registradas para Baja California Sur (Fig. 8).

Figura 8 Intervalos de tolerancia de cada especie a concentraciones de nitrato
(NO3 µmol l-1) en agua de mar = promedio y
 =
desviación estándar, = concentraciones
críticas para cambios en la taxocenosis de B.C.S.
=
desviación estándar, = concentraciones
críticas para cambios en la taxocenosis de B.C.S.
Fosfatos. Las macroalgas estudiadas se presentan en concentraciones de 0 a 1.8 µmol l-1 PO4 disueltos en la columna de agua; 0.4 y 0.9 µmol l-1 muestran ser valores críticos para la taxocenosis de Baja California Sur. Pocas especies muestran intervalos amplios de distribución a pesar de las bajas concentraciones de este compuesto en el agua. Ninguna especie se distribuye en promedios superiores a 1.8 µmol l-1 PO4 (Fig. 9).

Figura 9 Intervalos de tolerancia de las macroalgas a la concentración de
fosfatos (PO4 µmol l-1) en agua de mar
=
promedio y = desviación
estándar,  = concentraciones óptimas para las especies de macroalgas de
B.C.S.
= concentraciones óptimas para las especies de macroalgas de
B.C.S.
Silicatos. Las macroalgas se distribuyen dentro de intervalos de concentración de silicatos de 0.1 a 35 µmol l-1 en la columna de agua. La mayor parte de las especies se desarrollan en concentraciones menores a 10 µmol l-1. Muy pocas especies de los litorales de BCS se presentan en concentraciones de silicato promedio mayores a 20 µmol l-1 (Fig. 10).

Figura 10 Intervalo de tolerancia de macroalgas a la concentración de silicatos
(SO4 µmol l-1) en agua de mar,  = promedio y
=
desviación estándar, = concentraciones
óptimas para las especies de macroalgas de B.C.S
= promedio y
=
desviación estándar, = concentraciones
óptimas para las especies de macroalgas de B.C.S
Salinidad. Todas las especies de macroalgas estudiadas son esencialmente marinas y se desarrollan en concentraciones promedio entre 32 y 36.5 (línea de tendencia; Fig. 11); el límite superior es 38.5 y a partir de este valor no se registra ninguna especie evaluada y el límite inferior será de 19.

Figura 11 Intervalos de tolerancia de macroalgas a la salinidad del agua de mar
=
promedio, = desviación estándar, = valor mínimo y
máximo para bahía de La Paz, BCS, México, AGC = Agua del Golfo de
California, ASE =Agua Superficial Ecuatorial (Obeso-Nieblas et al., 2007), --- = Límite superior
e inferior (± 0.43), máxima diferencia actual como cambio hipotético en
BCS.
En la figura 11 se plantea un escenario en donde se muestran los valores mínimo y máximo para bahía de La Paz, BCS y los cambios en la salinidad por intrusión de Agua del Golfo de California (AGC) y Agua Superficial Ecuatorial (ASE) (Obeso-Nieblas et al., 2007); además, se plantea el escenario hipotético donde se suma la diferencia entre las dos masas de agua, como límite superior e inferior (± 0.43) donde se observa cambios taxocenóticos relacionados con la intrusión de las dos masas de agua a partir de 35.5.
Especies tolerantes. Las especies tolerantes presentaron los intervalos de mayor amplitud para cada variable (Tabla 2). En el caso de la temperatura, se encontraron especies que toleran cambios mayores a 5°C en el promedio del agua superficial del mar. Por otra parte, otras especies toleran cambios hasta de 1.5 µmol l-1 en las concentraciones de materia orgánica disuelta y mayores de 0.1 de pH. Las especies más tolerantes al nitrato presentaron intervalos mayores a 3 µmol l-1 y hasta 3.8 µmol l-1 en el promedio de la concentración en el agua de mar, así como, de mayores a 0.4 µmol l-1 en fosfato; mientras que, para silicato los intervalos fueron mayores a 10 µmol l-1.
Tabla 2 Especies más tolerantes por variable fisicoquímica en el agua de mar.
| Especies tolerantes | Temperatura > 5 °C | MOD >1.5 µmol l-1 | pH >0.1 | Nitrato >3 µmol l-1 | Fosfato >0.4 µmol l-1 | Silicato >10 µmol l-1 |
| Corallina vancouveriensis | X | X | X | X | X | |
| Griffithsia pacifica | X | X | X | X | ||
| Derbesia marina | X | X | ||||
| Gracilaria spinigera | X | X | ||||
| Ulva flexuosa | X | X | ||||
| Veleroa subulata | X | |||||
| Ceramium flaccidum | X | |||||
| Lithophyllum imitans | X | |||||
| Chaetomorpha linum | X | |||||
| Ceramium paniculatum | X | |||||
| Champia parvula | X | |||||
| Porphyra thuretii | X | |||||
| Ulva clathrata | X | |||||
| Rosenvingea intricata | X | X | ||||
| Gelidiella acerosa | X | |||||
| Laurencia papillosa | X | |||||
| Amphiroa valonioides | X | |||||
| Hypnea valentiae | X | |||||
| Jania adhaerens | X | |||||
| Padina durvillaei | X | |||||
| Valoniopsis pachynema | X | |||||
| Asparagopsis taxiformis | X | |||||
| Dictyopteris delicatula | X | |||||
| Hydroclathrus clathratus | X | |||||
| Bryopsis pennata | X | |||||
| Lithophyllum proboscideum | X | |||||
| Spyridia filamentosa | X | |||||
| Ectocarpus acutus | X | X | X | |||
| Antithamnionella spirographidis | X | X | X | X | ||
| Codium setchellii | X | X | ||||
| Dictyota dichotoma | X | |||||
| Hildenbrandia rubra | X | |||||
| Chaetomorpha linum | X | |||||
| Antithamnionella elegans | X |
Especies sensibles. Las especies sensibles son las que se distribuyen en intervalos estrechos de tolerancia de cada variable. Algunas especies se distribuyen en intervalos de amplitud menores a 1°C de temperatura superficial de agua de mar (Tabla 3), 0.5 µmol l-1 de concentración de materia orgánica disuelta, así como la variación menor a 0.5 µmol l-1 de nitrato, 0.1 µmol l-1 de fosfato y/o 1 µmol l-1 de silicato.
Tabla 3 Especies sensibles a las variaciones de fisicoquímicos en el agua de mar.
| Especies sensibles | Temperatura < 1 °C | MOD < 0.5 µmol l-1 | pH <0.01 | Nitrato < 0.5 µmol l-1 | Fosfato < 0.1 µmol l-1 | Silicato < 1 µmol l-1 |
| Dictyosphaeria cavernosa | X | |||||
| Ceramium procumbens | X | X | X | X | X | X |
| Sargassum lapazeanum | X | X | X | X | X | X |
| Botryocladia neushulii | X | X | X | X | X | |
| Chondria acrorhizophora | X | X | X | X | X | |
| Pterocladia capillacea | X | |||||
| Cladophora hesperia | X | |||||
| Ceramium fimbriatum | X | X | ||||
| Colpomenia ramosa | X | |||||
| Gracilaria pachydermatica | X | |||||
| Liagora californica | X | X | X | X | ||
| Veleroa subulata | X | |||||
| Ceramium paniculatum | X | X | ||||
| Digenea simplex | X | |||||
| Prionitis cornea | X | X | ||||
| Gelidiella acerosa | X | |||||
| Amphiroa rigida | X | |||||
| Dictyota crenulata | X | |||||
| Hypnea johnstonii | X | |||||
| Veleroa subulata | X | X | ||||
| Pterocladia capillacea | X | X | ||||
| Dictyosphaeria cavernosa | X | X | ||||
| Laurencia pacifica | X |
Variables fisicoquímicas y nutrientes que más afectan la taxocenosis. El análisis discriminante (Fig. 12) muestra que el canónico 1, clasificó con el 91.9% de confianza las clases de macroalgas, según las concentraciones de compuestos químicos en el agua de mar donde se han registrado. Se determinó que las Ulvophyceae y las algas rojas (Florideophyceae) se distribuyen en intervalos amplios de concentración de oxígeno, nitrato y silicato, mientras que, las Phaeophyceae se restringen en intervalos estrechos y toleran menor concentración de silicatos en el agua (7.78 % de la varianza) y requieren mayores concentraciones de nitrato en la columna de agua y oxígeno disuelto.

Figura 12 Prueba de análisis discriminante que separó las clases de macroalgas de Baja California Sur, México, respecto a los promedios (2005 -2012) de parámetros fisicoquímicos y nutrientes del agua de mar, de los lugares donde se han registrado en el planeta. El canónico 1 corresponde al vector(es) más largo(s) que son las variables que más separan las Clases.
DISCUSIÓN
El desarrollo de un modelo de distribución de especies implica múltiples ajustes; por ejemplo, tener en cuenta las sinonimias. Ello debido a la determinación taxonómica inicial y los cambios de nomenclatura de las especies, así como, por el uso de sinonimias válidas en la base general ITIS durante el proceso de obtención de las ocurrencias de las especies en el planeta; estas han sido actualizados en otras bases especializadas, como ALGABASE y el World Register of Marine Species (WORMS).
De la lista taxonómica lograda se presentan varios casos, v.g., Antithamnionella glandulifera que cambió en 1989 a Antithamnionella spirographidis (Schiffner) E.M.Wollaston. Ceramium fimbriatum del que se hizo una revisión del género en 2008 y cambió a Gayliella fimbriata, así como, C. flaccidum sinonimia de Gayliella flaccida (T.O.Cho & L.J.McIvor, 2008). Laurencia papillosa, que pasa a ser Palisada perforata (Bory) K.W.Nam en el 2007. Porphyra thuretii, donde la especie es la misma, pero cambió de género a Pyropia thuretii en 2011. Mientras que, Prionitis cornea pasó a ser Grateloupia cornea en 2001. Y, por último, Pterocladia capillacea cambió a Pterocladiella capillacea (S.G.Gmelin) Santelices & Hommersand. El uso de la información de registros para las sinonimias obtenidas por el programa confiere robustez a la base de datos obtenida.
Las variaciones observadas en la taxocenosis de macroalgas se asocian principalmente a la variación en la temperatura superficial del agua de mar, que determina la distribución espacial y el cambio temporal de los taxa. Esto produce cambios a nivel de atributos comunitarios de las macroalgas, por ejemplo, en la riqueza y composición de especies encontradas por mes (Siqueiros-Beltrones, 1998); mientras que, la diferencia de temperatura en el agua de mar superficial entre invierno (febrero) y verano (septiembre) para 1995 en la bahía de La Paz fue de 16 grados centígrados; actualmente encontramos esa diferencia en 11°C (Fig. 2). La tolerancia de las especies estudiadas aquí, se determinó entre 9 y 26 °C de temperatura superficial del agua de mar, un intervalo de 15°C. Así, son tolerantes a temperaturas más bajas, que los promedios actuales en aguas de BCS, lo que sugiere que estas especies podrían distribuirse actualmente en aguas más frías.
Por tanto, esto explica que la taxocenosis en BCS pudo cambiar e incluiría especies introducidas, que han arribado a costas sudcalifornianas en los últimos años (Riosmena et al., 2014). Las especies de macroalgas agrupadas en las clases Bangiophyceae y Ulvophyceae tendrían mayor tolerancia ante un escenario de aumento de temperatura, mientras que, promedios de temperatura en el agua de mar entre 10 y 16 °C favorecerían el crecimiento de macroalgas pardas (Phaeophyceae); temperaturas menores a los 10 °C originarían nuevas taxocenosis, dado que la mayor parte de las especies que fueron analizadas, no presentan intervalos en estos promedios de temperatura, en su distribución a nivel mundial. En un escenario de cambio, el aumento del promedio de la temperatura máxima en 1 °C en BCS tendría un mayor efecto en la composición de las especies de la comunidad de macroalgas (Fig. 2), debido a que pocas especies presentan amplitudes que superen los máximos actuales; mientras que, si disminuyera la temperatura en un promedio de 2 °C, un mayor número de especies de esta taxocenosis sobreviviría, debido a las condiciones iniciales del sistema.
Cambios globales de temperatura, también implicarían cambios en los promedios decadales en las concentraciones de oxígeno y a su vez a diferentes niveles taxonómicos de las macroalgas. Por ejemplo, la disminución a concentraciones promedio menores a 4.2 ml l-1 de oxígeno, promovería la variación en la composición de las especies registradas para Baja California Sur. El oxígeno disuelto (OD) en agua de mar fue la variable con mayor aporte a la diferenciación de los ambientes, donde se desarrollan las clases de macroalgas. Cruz-Ayala (1996) encontró en la bahía de La Paz (BCS) mayor riqueza en áreas con corrientes y atribuyó esto a la mayor concentración de oxígeno en el agua.
Se ha observado, que la disminución en la velocidad de la circulación de las masas de agua con bajas concentraciones de OD, los cambios en los patrones del viento, el aumento en la estratificación y la baja concentración inicial de OD en aguas superficiales de temperatura alta (el oxígeno preformado), pueden ser los responsables de los eventos de hipoxia en el mar abierto; es así como, la concentración de oxígeno disuelto en el agua del Pacífico mexicano varía entre lo hipóxico (<1 ml l-1) en aguas profundas hasta la sobresaturación (>10 ml l-1) (Rubach et al., 2017). Adicionalmente, en los sistemas costeros la materia orgánica que ingresa por cualquier vía consume oxígeno durante su descomposición y favorece la anoxia (De La Lanza-Espino et al., 2011; De La Lanza-Espino & Gutiérrez-Mendieta, 2017).
La pérdida de especies de macroalgas en un hábitat presume disminuciones en el promedio de las concentraciones de oxígeno en la columna de agua (Fig. 4). En BCS, en donde probablemente la disminución o el aumento de 1 ml l-1 en la concentración promedio de oxígeno en el agua propiciaría diferentes taxocenosis, debido que aquí se encontró, que ninguna especie presenta intervalos de tolerancia mayores a 2 ml l-1. Concentraciones promedio mayores a 5.3 ml l-1 de OD propiciará, al menos para un tercio de las especies, salir de concentraciones óptimas; otro punto crítico sería el aumento a promedios mayores a 5.8 ml l-1 a las que probablemente sobreviva apenas un tercio de la composición actual; las mismas que muestran intervalos de tolerancia a mayores concentraciones de oxígeno en la columna de agua.
Se conoce que el OD está estrechamente relacionado con el pH del agua de mar, es decir, procesos como la respiración generan la liberación de CO2, lo que disminuye el pH. Por tanto, aguas hipóxicas son también aguas de menor pH. Por ejemplo, aguas de bajo pH sobre la plataforma continental se presentan desde Canadá hasta las costas de Baja California en México (Feely et al., 2008); si las corrientes de California se hicieran más severas por disminución en la temperatura del océano, probablemente el fenómeno descrito anteriormente alcanzaría las costas de Baja California Sur, presentándose aguas anóxicas o hipóxicas en áreas de surgencias.
Las macroalgas rojas (Florideophyceae y Bangiophyceae) se han registrado en ambientes más básicos (mayor pH) que Ulvophyceae y Phaeophyceae; es así como, se encontró que las características fisicoquímicas y de concentración de los nutrientes de los ambientes donde se desarrollan algas pardas (Phaeophyceae), algas rojas (Florideophyceae y Bangiophyceae) y algas verdes (Ulvophyceae) son diferentes.
Las Ulvophyceae presentan mayor tolerancia al cambio en el pH en el agua (desviación estándar; Fig. 5), este grupo es el más tolerante ante un escenario de cambio en conjunto con bajas concentraciones de oxígeno disuelto, como lo sugieren Rubach et al. (2017). El límite superior de pH para la supervivencia de las macroalgas (8.37) puede deberse a la acumulación de la materia orgánica; por ejemplo, en el sedimento de las lagunas costeras, dado que los nutrientes procedentes de la columna de agua ya remineralizados (fracción reciclada) y de aportes fluviales (fracción nueva) sin descomponerse, se convierten en una fuente potencialmente tóxica, porque el amonio se transforma en amoniaco (gas) en condiciones alcalinas por arriba de pH 8 (De La Lanza-Espino & Gutiérrez-Mendieta, 2017).
La materia orgánica disuelta contiene formas de nitrógeno y fósforo orgánicos, las cuales al descomponerse o remineralizarse se transforman en nutrientes inorgánicos y puede generar condiciones de eutrofia (De la Lanza-Espino, 2014). En sistemas costeros, los compuestos inorgánicos nitrogenados varían espacial y temporalmente según condiciones locales, desde lo indetectable hasta más de 30 µM para NO3+, según las condiciones prevalecientes de óxido-reducción (De la Lanza-Espino, 2014; Okolodkov et al., 2014). En este ejercicio se encontró que las especies de la taxocenosis propuesta se desarrollan en concentración de nitratos menores a 10 µmol l-1.
La concentración de ortofosfatos disueltos en las lagunas costeras puede oscilar desde lo indetectable hasta 5 µM, sin embargo, pueden aumentar a valores cercanos a 10 µM como resultado de la complejidad local y la contaminación (De la Lanza-Espino, 1994). Así, altas concentraciones de ortofosfatos en sistemas costeros de BCS pondrían en riesgo la supervivencia de las especies que actualmente exhiben máximos de tolerancia de 0.5 µmol l-1 de fosfato en su distribución a nivel mundial; en consecuencia, las especies de macroalgas estudiadas para BCS, no sobrevivirían a condiciones con concentración de fosfatos mayor a 0.9 µmol l-1 en promedio. Los ortofosfatos pueden retirarse del agua hasta en un 60% por adsorción en sedimentos (efecto amortiguador) (Hobbie et al., 1975), lo que explica las bajas concentraciones de este nutriente en la columna de agua de mar.
Pocas especies de macroalgas toleran concentraciones de silicato mayores a 20 µmol l-1. Efectivamente, estas especies serán más tolerantes a mayores variaciones en la concentración de silicatos en el agua. Mientras que, especies que presentaron intervalos menores a 1 µmol l-1, se proponen aquí como las especies más sensibles.
La salinidad superficial en bahía de La Paz muestra un patrón estacional e inverso a la temperatura que responde a la presencia de diferentes masas de agua en la región. Por ejemplo, en mayo de 2001 se registró la presencia de Agua del Golfo de California con salinidad de 35.35 y esta disminuyó para julio (35.17) por la mayor influencia de Agua Superficial Ecuatorial combinada con Agua del Golfo de California, presentando un valor mínimo durante octubre (34.92) (Obeso-Nieblas et al., 2007). La diferencia en la variación de la salinidad en 2001 en la bahía de La Paz fue de 0.43, lo que indica la presencia de diferentes masas de agua. Sí solo aumentara el doble de lo normal, es decir 0.86 y llega a 35.81, probablemente se conservarían 18 de las especies de las 71 filtradas, que se han registrado en estas concentraciones (Fig. 11).
La salinidad en el agua también cambia respecto a la profundidad (Flores-Verdugo et al., 1993). Además, las mayores variaciones en la salinidad en el ambiente marino, también se presentan en la desembocadura de los grandes ríos, en zonas estuarinas. Probablemente, el que las macroalgas se desarrollen en las costas que recibe escorrentías, explique por qué más especies estudiadas toleran menores concentraciones de salinidad y otros compuestos en el agua que las especies, que toleran altas concentraciones. Es decir, más especies sobrevivirían si disminuyera la salinidad en el agua de mar, que las que sobrevivirían ante un aumento de la salinidad en sistemas costeros de BCS.
Como conclusión, la sensibilidad o tolerancia de las especies al cambio (aumento o disminución extraordinaria de compuestos en el agua) determina los cambios a nivel comunitario. Así, se puede aproximar cuantitativamente la tolerancia de cada especie a las concentraciones de un compuesto químico en el agua, como el intervalo de distribución estadística de la especie en la variable, misma que representa su distribución física en el planeta. Las variaciones naturales y por exceso en sistemas costeros en proceso de eutrofización producirán distinción en composición florística (taxocenosis). Así, se propone incluir especies aquí llamadas tolerantes a variación de concentraciones fisicoquímicas y/o nutrientes para conformar protocolos de referencia, que permitan comparar la distribución del contenido de minerales, con otras áreas del mundo, porque esto clasificará áreas donde se presenta contaminación (Tabla 2). Mientras que, las especies con intervalos estrechos pueden ser indicadores de la concentración del nutriente y/o fisicoquímico, en donde estas se presenten (Tabla 3).
La variación en la concentración de materia orgánica disuelta y silicatos influirá directamente en cambios en la taxocenosis de macroalgas, debido a los consecuentes cambios en el oxígeno disuelto y el pH en el agua, desde grandes grupos como clases. Las especies de algas pardas (Phaeophyceae) son más sensibles al cambio, porque requieren aguas con mayores concentraciones de nutrientes, bien oxigenadas y templadas para su supervivencia; por tanto, una disminución del promedio anual en el oxígeno en la columna de agua podría ocasionar, que los taxa de este grupo desaparecieran primero, mientras que, macroalgas verdes (Ulvophyceae) y rojas (Florideophyceae y Bangiophyceae) persistirían ante mayores condiciones de estrés.
Se proponen posibles taxocenosis por cambios en el estado de calidad de agua, como resultado de una prueba de relación entre variables estudiadas en un mapa de calor (Fig. 13); las variables pH, silicatos, materia orgánica disuelta y el oxígeno disuelto, conforman el grupo que proporciona mayor aporte a la agrupación de especies, por similitud del hábitat, (mayor variedad de tonalidades o gradiente) (Fig. 13), donde se distinguen cuatro asociaciones que indicarían condiciones específicas del agua. Se propone una clasificación de estados de sistemas costeros, que a partir de condiciones fisicoquímicas y de la concentración de nutrientes específicas, debería producir una taxocenosis de macroalgas dependiente de la condición: “buena, moderada, mala y muy mala”.

Figura 13 Mapa de calor que muestra las posibles taxocenosis de macroalgas de Baja California Sur, México, de acuerdo con el estado de calidad del agua. Los colores indican la intensidad de la relación o los valores de la variable representada, desde el blanco (máxima relación o valor más alto) pasando por el amarillo y naranja hasta el rojo intenso (mínimo valor). Las agrupaciones por similitud entre especies, discrimina en 4 posibles taxocenosis que indicarían condiciones específicas del agua.
Las especies sensibles Dictyosphaeria cavernosa, Botryocladia neushulii, Chondria acrorhizophora, Ceramium fimbriatum, P. capillacea, Veleroa subulata, Digenea simplex, Gelidiella acerosa, Amphiroa rigida, Dictyota crenulata, Hypnea johnstonii y Laurencia pacifica se proponen como indicadoras dentro de la asociación macroalgal con condiciones “buenas” de calidad de agua; requerirán pH básicos y menores concentraciones promedio de silicatos y materia orgánica disuelta en la columna de agua, así como, aguas oxigenadas y oligotróficas con bajas concentraciones de materia orgánica.
Se propone un estado de calidad con promedios de concentración intermedias o “moderadas” donde sobreviven probablemente la mayor parte de las especies, según la teoría del disturbio intermedio (Roxburgh et al., 2004). Y las especies Ceramium procumbens, Ceramium paniculatum, Sargassum lapazeanum y Liagora californica serían indicadoras de este estado, que principalmente se ha enriquecido en materia orgánica disuelta y silicatos.
Un estado de calidad del agua “malo” por el contrario, implica altas concentraciones de silicatos y materia orgánica disuelta en el agua, al igual que pH más bajos y menores promedios de oxígeno, como se ha establecido para otros índices de estados tróficos (Vollenweider et al., 1998). Las especies representativas serían Cladophora hesperia, Colpomenia ramosa, Gracilaria pachydermatica, Grateloupia cornea.
En los estados “muy malos” las condiciones descritas se intensifican, especialmente con el aumento en los silicatos en el agua y menores concentraciones en el promedio del oxígeno disuelto, lo que determinaría una taxocenosis conformada por pocas especies (Fig. 13) de las estudiadas y/o las más tolerantes a cambios fisicoquímicos del agua. En condiciones muy malas sobrevivirían pocas especies. En su revisión en campo, Cruz-Ayala (1996) también encontró que la Familia Corallinaceae se presentó con mayor frecuencia en todas las localidades, así como, el género Codium, tal y como fue asociado en este estudio en el mapa de calor, con datos satelitales y los datos de registros de presencia en el planeta.
Esta métrica teórica deberá ser probada en condiciones ambientales que suponen las condiciones aquí propuestas, para analizar la idoneidad del índice, así como su precisión. Otras formas de bioindicación incluyen la abundancia o cobertura y la presencia de especies oportunistas (Juanes et al., 2008), este índice con macroalgas es una propuesta desde la asociación de especies y las especies sensibles características de cada estado de calidad.

