











 nueva página del texto (beta)
nueva página del texto (beta) Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
Permalink
Introducción
En México, la producción de trucha arcoíris (Oncorhynchus mykiss Walbaum) en el 2014 fue de 15,695 toneladas con un valor cercano a los 876 millones de pesos (Conapesca, 2015). Como se puede apreciar, la trucha representa un importante recurso económico y social en los estados de México, Michoacán, Veracruz y Puebla, que son los principales productores en el país. Durante el proceso de cultivo de esta especie, el alimento representa entre el 50 y 60% del costo de producción (FAO, 2014), y actualmente las formulaciones comerciales de alimento balanceado para trucha tienen como ingredientes base la harina y el aceite de pescado. La producción global de ambos insumos no se incrementará en los próximos años, y por ende, el desarrollo de la producción de esta especie será insostenible si ambos ingredientes se mantienen como las fuentes principales de proteína y lípidos en las dietas comerciales (Hardy, 2010). Los productos de origen vegetal se han propuesto como una alternativa seria a los ingredientes de pescado, y la expectativa es que permitan el crecimiento de las especies acuáticas con un mínimo de impacto en el ambiente y a un costo menor (Gatlin et al., 2007). Uno de los productos de origen vegetal que se ha propuesto como fuente de proteína para su inclusión en las dietas de trucha es el gluten de maíz (GM). Resultado de la molienda húmeda de la semilla del maíz, el GM tiene un alto contenido de proteína cruda (60%) y es altamente digerible, con coeficientes de digestibilidad aparentemente mayores al 95% (Saez et al., 2016). El GM se utiliza en diversas formulaciones de dietas de algunas especies comerciales marinas y de salmón, con inclusiones en un rango de entre 10 y 15% (Gatlin et al., 2007). En el caso de la trucha arcoíris, Sáez et al. (2016) reportaron que una inclusión de hasta 18% de GM en la dieta de truchas de aproximadamente 500 g no afecta el crecimiento y la deposición de nutrientes en el cuerpo. Sicuro et al. (2017) reportaron que el crecimiento y los parámetros hematológicos no fueron afectados en juveniles trucha de 50 g alimentados con dietas de GM al 40% en sustitución de harina de pescado. Gerile & Pirhonen (2017) encontraron en truchas de 80 g alimentadas con una dieta con GM como fuente de proteína no se afectó el consumo de oxígeno durante periodos de nado forzado.
Por otro lado, el estatus nutricional tiene una fuerte influencia en la respuesta inmune de los organismos acuáticos (Kiron, 2012). La cantidad y calidad de las fuentes de proteína de la dieta tienen un efecto sobre las respuestas inmunes (Hardy, 2001), ya que un consumo insuficiente, afectará el contenido de proteínas de las células y eventualmente las incapacitará. De acuerdo con Tacon (1995), el uso de proteínas de origen vegetal puede afectar la respuesta inmune de los organismos acuáticos con hábitos carnívoros. Por ello, el objetivo del presente trabajo es determinar el efecto de dietas con altos niveles de inclusión de GM en el crecimiento, la deposición de nutrientes, la excreción de fósforo (P) y nitrógeno (N), así como la actividad explosiva de los macrófagos en juveniles de trucha arcoíris.
Materiales y métodos
Dietas experimentales. Se formularon 3 dietas con un contenido mínimo de 40% de proteína cruda y 12% de lípidos, para cubrir los requerimientos previamente reportados para la especie (NRC, 2011). La harina de pescado se sustituyó con gluten del maíz (Glutimex, 60% proteína cruda, CP Ingredientes S. A. de C. V., México) en 50, 75 y 100% de la porción de la proteína (Tabla 1). Como fuente de lípidos se utilizó una mezcla 1:1 de aceites de soya y linaza, así como aceite de pescado (Drotasa S. A. de C. V., México). Se utilizó dextrina (Sigma Aldrich Co., Estados Unidos) como única fuente de carbohidratos. Asimismo, se empleó una mezcla de vitaminas y minerales (Micro Rovimix para peces carnívoros, DSM Nutrional Products de México S. A. de C. V., México) y gluten de trigo (Sigma Aldrich Co., Estados Unidos) como aglutinante. Finalmente, se adicionó a-celulosa (Sigma Aldrich Co., Estados Unidos) para llevar la dieta al 100%. Las dietas se elaboraron de acuerdo con lo descrito por Cruz et al. (2011), y como dieta control se utilizó Api-trucha 2 (composición reportada por el fabricante de 45% proteína y 15% lípidos, Malta-Cleyton de México, México).
Tabla 1 Formulaciones y composición proximal de las dietas experimentales para juveniles de trucha arcoíris (Oncorhynchus mykiss Walbaum, 1792).
| Ingredientes (g kg-1) | Tratamientos | ||
|---|---|---|---|
| G50 % | G75 % | G100 % | |
| Harina de pescado | 307.5 | 153.85 | 0 |
| Gluten de maíz | 333.5 | 500.25 | 667 |
| Mezcla de aceites1 | 40 | 40 | 40 |
| Aceite de pescado | 30 | 30 | 30 |
| Lecitina de soya | 50 | 50 | 50 |
| Dextrina | 100 | 100 | 100 |
| Mezcla de vitaminas y minerales2 | 40 | 40 | 40 |
| Aglutinante | 50 | 50 | 50 |
| α-celulosa | 48.8 | 35.9 | 23 |
| Composición proximal (% en base seca) | |||
| Proteína | 40.6 ± 0.1 | 40.9 ± 0.2 | 37.5 ± 0.4 |
| Lípidos | 15.1 ± 0.1 | 16.4 ± 0.1 | 15.9 ± 0.2 |
| Cenizas | 8.1 ± 0.1 | 7.9 ± 1.4 | 6.2 ± 0.1 |
1 Aceites de soya y linaza, mezcla 1:1
2 Mezcla de vitaminas y minerales (g/kg): ácido ρ-aminobenzoico, 1.45; biotina, 0.02; myo-inositol, 14.5; ácido nicotínico, 2.9; pantotenato de Ca, 1.0; piridoxina-HCl, 0.17; riboflavina, 0.73; tiamina-HCl, 0.22; menadiona, 0.17; α-tocoferol, 1.45; cyanocobalamina, 0.0003; calciferol, 0.03, L-ascorbil-2-fosfato-Mg, 0.25; ácido fólico, 0.05; cloruro de colina, 29.65, NaCl, 1.838; MgSO4·7H2O, 6.85; NaH2PO4·2H2O, 4.36; KH2PO4, 11.99; Ca(H2PO4)2·2H2O, 6.79; citrato-Fe, 1.48; lactato de Ca, 16.35; AlCl3·6H2O, 0.009; ZnSO4·7H2O, 0.17; CuCl2, 0.0005; MnSO4·4H2O, 0.04; KI, 0.008 and CoCl2, 0.05.
Obtención de organismos. Se obtuvieron juveniles de trucha arcoíris del Centro de Producción Acuícola El Zarco y se trasladaron al Laboratorio de Producción Acuícola de la FES Iztacala, UNAM. Los organismos se mantuvieron en tanques de 500 L por un periodo de 15 días y se alimentaron con una dieta comercial (Api-trucha 2).
Prueba de alimentación. Para la prueba se utilizaron 8 tanques de polipropileno de 500 L. En cada tanque se colocaron aleatoriamente 45 juveniles con un peso de 14.7 ± 0.2 g (promedio ± error estándar), y cada dieta se suministró por duplicado en grupos de juveniles. Los peces se alimentaron diariamente con la dieta respectiva, considerando un 7% de la biomasa total por tanque. La ración diaria se dividió en 2 porciones, en la mañana y al atardecer. Para pesar a los organismos cada 10 días, se anestesiaron con MS-222 (ácido metasulfónico 3-aminobenzoato etílico, Sigma Aldrich Chemical, E.U.) a 150 mg L-1. El tamaño de la ración se ajustó de acuerdo con el nuevo peso. Treinta minutos después de cada alimentación se recogió el alimento no consumido y se cuantificó para determinar la tasa de ingesta. Posteriormente, y con la finalidad de mantener la calidad del agua, se limpiaron las heces fecales del fondo de cada tanque sifoneando con una manguera. Durante la prueba, los parámetros de la calidad del agua fueron (media ± error estándar): oxígeno disuelto 4.5 ± 1 mg L-1, pH 7.9 ± 0.5, temperatura del agua 14 ± 1 °C y concentración de amonio de 0.00 mg L-1. La prueba de alimentación duró 90 días. Los peces se mantuvieron sin alimento por 24 horas y se pesaron para obtener el crecimiento. Seis organismos de cada tratamiento se seleccionaron al azar y se utilizaron para determinar el consumo de oxígeno y la excreción de fósforo y nitrógeno. Cinco peces de cada repetición se seleccionaron al azar y se sacrificaron con una sobredosis del anestésico MS-222 (ácido metasulfónico 3-aminobenzoato etílico, Sigma Aldrich Chemical, Estados Unidos) a 200 mg L-1, después se tomaron muestras de sangre que se colocaron en tubos con anticoagulante (EDTAK2, tubos BD Vacutainer, BD Diagnósticos, México) y una solución balanceada de sales. Estas muestras se utilizaron para cuantificar el número de linfocitos. Finalmente, se realizó la disección de los riñones para la prueba del estallido respiratorio de los macrófagos, además de muestras para el análisis de contenido de proteína y lípidos en músculo e hígado.
Parámetros de crecimiento. El crecimiento se determinó mediante las siguientes fórmulas:
Ganancia en peso = [(peso final - peso inicial) / peso inicial] x 100;
Tasa de crecimiento especifico = [(ln peso final - ln peso inicial) / tiempo de alimentación] x 100
Consumo de oxígeno y excreción de fósforo (P) y nitrógeno (N). Se utilizó un sistema de recirculación cerrado que consistió en 10 frascos de 1 L conectados en serie por tubos de plástico (Cruz et al., 2011). Los frascos se llenaron lentamente con agua a 14 °C y se colocó un pez por cada frasco. Se tomaron muestras iniciales de agua (concentraciones de P y N = 0 mg L-1) y se determinó el oxígeno disuelto (oxímetro modelo 85, YSI Incorporated, OH, EU), posteriormente, los frascos fueron cerrados herméticamente. Después de 20 min, los frascos fueron abiertos, se midió nuevamente el oxígeno disuelto y se tomaron nuevas muestras de agua para determinar la excreción de P (como PO4 3-) y N (como NH3-N), de acuerdo con las técnicas de molibdovanadato y Nessler (Clesceri et al., 1998). El consumo de oxígeno se calculó como la diferencia entre la concentración inicial y la final.
Análisis químicos. Los contenidos de proteína de las muestras de músculo e hígado se determinaron mediante la técnica de la AOAC (1990), la cual emplea un sistema de destilación Kjeltec 2100 (Foss A/S, Dinamarca). El contenido de lípidos se determinó mediante la técnica de Blight y Dyer (1959), que utiliza cloroformo y metanol para la extracción.
Cuantificación de los linfocitos. Los tubos con la sangre, anticoagulante y la solución balanceada de sales se agregaron con 3 ml de ficoll (Ficoll-Paque PLUS, GE Healthcare Bio-Science AB, Suecia) y se centrifugaron a 2,260 rpm durante 30 min. Se retiró el sobrenadante y se aisló la capa con los linfocitos, para limpiarlos, se colocaron en un tubo nuevo con la solución balanceada de sales, y se centrifugaron a 810 rpm durante 10 min; se retiró el sobrenadante y se agregó una nueva solución balanceada de sales, con la cual se centrifugó nuevamente. Finalmente, se tomó la capa limpia con los linfocitos, se tiñeron con el colorante de Wright y se contaron en una cámara de Neubauer.
Estallido respiratorio de los macrófagos. El aislamiento y cultivo de los macrófagos del riñón se realizó de acuerdo con Chung y Secombes (1988) y Secombes (1990). La actividad de anión superóxido de los macrófagos (estallido respiratorio) se determinó por la reducción del colorante nitro-azul tetrazolium (NBT) con un lector de placa Sunrise (Tecan Group Ltd., Suiza) en una longitud de onda de 620 nm (OD620). Posteriormente, se determinó el número de macrófagos en cada uno de los pocillos. Los resultados están expresados como OD620 por 104 células.
Análisis estadístico. Los datos de crecimiento, de contenido de proteína y lípidos en los tejidos, del número de linfocitos y del estallido respiratorio de los macrófagos se analizaron para normalidad y homosedasticidad con las pruebas W de Shapiro y Wilk, así como de Barlett, respectivamente (Zar, 1999). Debido a que los datos mostraron normalidad y homosedasticidad, se trataron con un ANDEVA de una vía (Prism 6 for MAC OS X, GraphPad Software Inc., Estados Unidos). Las diferencias significativas entre los tratamientos se evaluaron con una prueba de LSD de Fisher (Zar, 1999) y las diferencias se determinaron con un error de 5% (p <0.05) para cada grupo de comparaciones.
Resultados
En la Tabla 2 se muestran los datos de crecimiento de los juveniles de trucha arcoíris alimentados con diferentes inclusiones de gluten de maíz durante 90 días. El peso final (PF) fue significativamente (p <0.0001) más bajo en el grupo de truchas alimentadas con la dieta G100%, con respecto a los demás grupos. De la misma forma, los valores de la ganancia en peso (GP) y la tasa de crecimiento específico (TCE) en los organismos alimentados con G100% fueron significativamente (p <0.0001) más bajos que los observados en los otros grupos.
Tabla 2 Peso inicial (PI), peso final (PF), ganancia en peso (GP) y tasa de crecimiento especifico (TCE) de juveniles de trucha arcoíris (Oncorhynchus mykiss Walbaum, 1792) alimentados con diferentes niveles de gluten de maíz. Los valores representan la media de tres réplicas ± error estándar. Medias con letras diferentes en la misma línea difieren significativamente (p <0.05).
| Control | G50 % | G75 % | G100 % | |
|---|---|---|---|---|
| PI (g) | 14.9 ± 0.03 | 14.7 ± 0.07 | 14.7 ± 0.1 | 14.8 ± 0.1 |
| PF (g) | 70 ± 5a | 63 ± 4a | 56 ± 3a | 29 ± 0.3b |
| GP (%) | 373 ± 29a | 322 ± 29a | 284 ± 22a | 102 ± 1b |
| TCE (% día-1) | 1.7 ± 0.06a | 1.5 ± 0.07ª | 1.5 ± 0.06a | 0.8 ± 0.005b |
La Figura 1 muestra el contenido de proteína (a) y de lípidos (b) en el músculo y en el hígado. En músculo se observó una deposición significativamente (p <0.0001) menor de proteína en las truchas alimentadas con la dieta G100% con respecto al resto de los grupos. Respecto al hígado, el contenido de proteína fue significativamente (p <0.0001) más bajo en el grupo control que en los grupos experimentales. La deposición de lípidos en el músculo se incrementó conforme aumentó la inclusión de GM en la dieta, por lo tanto, el grupo alimentado con la dieta G100% mostró un contenido significativamente mayor. En el hígado, el contenido de lípidos fue significativamente (p <0.0001) menor en los organismos alimentados con la dieta G75% con respecto a los demás.
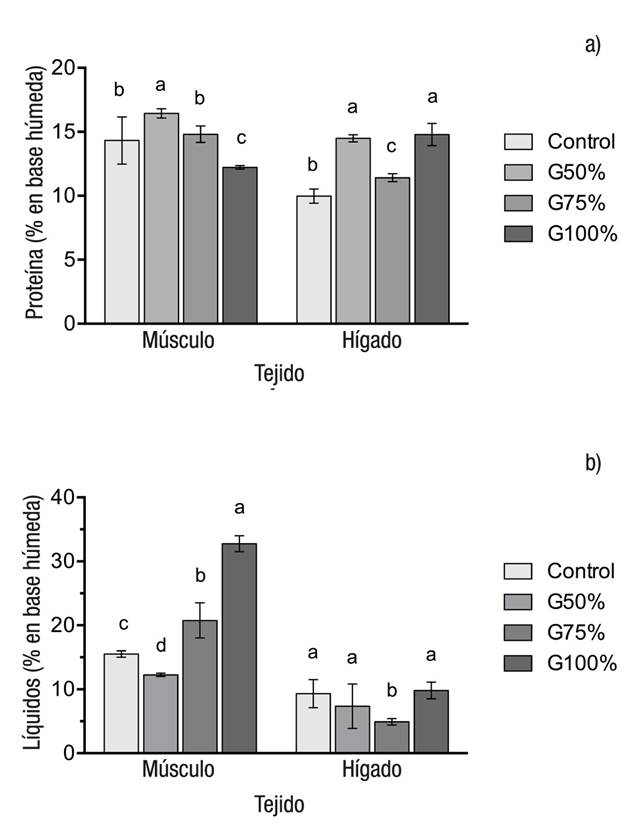
Figuras 1a-b Contenidos de proteína (a) y de lípidos (b) en el músculo e hígado de juveniles de trucha arcoíris (Oncorhynchus mykiss Walbaum, 1792), alimentados con diferentes niveles de gluten de maíz. Las barras representan la media de tres réplicas ± error estándar. Letras diferentes muestran diferencias significativas entre las medias (p <0.05).
La Figura 2 muestra el consumo de oxígeno (a), la excreción de N y de P (c). El consumo de oxígeno fue significativamente (p <0.0001) menor en el grupo de organismos alimentados con la dieta control respecto de aquellos alimentados con las dietas experimentales. Los organismos alimentados con las dietas G75% y G100% mostraron los valores más altos. De la misma forma, la excreción de N (como N-NH3) fue menor en los juveniles alimentados con la dieta control. Los organismos alimentados con las dietas experimentales no mostraron diferencias significativas entre ellos, pero los valores más bajos se observaron en la dieta G75%. Finalmente, la excreción de P (como PO4 3-) no mostró diferencias significativas entre los grupos, pero se observaron valores más bajos en los organismos alimentados con las dietas G50% y G75%. Los valores más altos se registraron en los organismos alimentados en la dieta control.

Figuras 2a-c Consumo de oxígeno (a), excreción de nitrógeno (b) y de fósforo de juveniles de trucha arcoíris (Oncorhynchus mykiss Walbaum, 1792) alimentados con diferentes niveles de gluten de maíz. Las barras representan la media de cinco réplicas ± error estándar. Letras diferentes muestran diferencias significativas entre las medias (p <0.05).
En la Figura 3a se muestra el número de linfocitos, donde el valor significativamente (p <0.0001) más alto fue observado en los organismos alimentados con la dieta G50% con respecto a los demás tratamientos. Respecto a la actividad explosiva de los macrófagos (Fig. 3b) no se observaron diferencias significativas entre los grupos.
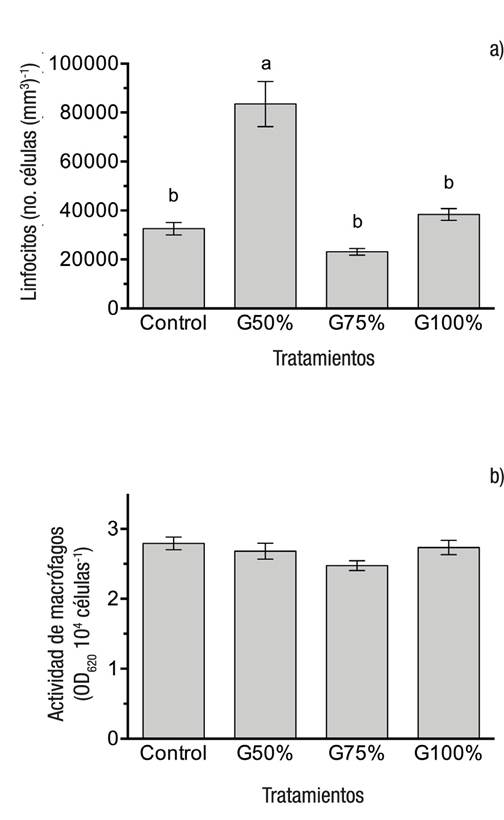
Figuras 3a-b Número de linfocitos (a) y actividad explosiva de los macrófagos (b) de juveniles de trucha arcoíris (Oncorhynchus mykiss Walbaum, 1792) alimentados con diferentes niveles de gluten de maíz. Las barras representan la media de cinco réplicas ± error estándar. Letras diferentes muestras diferencias significativas entre las medias (p <0.05).
Discusión
Uno de los retos que enfrenta la producción de trucha arcoíris en México es el desarrollo de nuevas formulaciones que permitan un cultivo más eficiente, tanto desde el punto de visto ambiental como económico. El GM se ha señalado como una fuente de proteína alternativa a la harina de pescado (Gatlin et al., 2007). De acuerdo con los resultados obtenidos en el presente trabajo, la dieta con una sustitución del 50% de la HP con el GM no afectó el crecimiento de los juveniles de trucha, la deposición de nutrientes en músculo e hígado así ni la actividad de los macrófagos.
Los organismos alimentados con la inclusión de GM mostraron una disminución en los parámetros de crecimiento determinados, particularmente aquéllos alimentados con las dietas G75% y G100%. De acuerdo con Gatlin et al. (2007) y Saez et al. (2016), la inclusión de GM en dietas de salmónidos y otras especies de peces marinos es de un máximo de 15%. Información reciente (Sicuro et al., 2017) señalan que hasta un 40% de sustitución con GM no causa efectos negativos en el crecimiento de juveniles trucha. En este trabajo se determinó que un 50% de la porción de la proteína puede ser provisto por GM. Por encima de este nivel es probable que un balance inadecuado de aminoácidos esenciales, particularmente la deficiencia de lisina del GM, afecte el crecimiento.
Los contenidos de proteína en el músculo fueron similares a los reportados por Aguillón et al. (2017) en juveniles de trucha arcoíris alimentados mediante dietas con aislado de proteína de soya y polvo de Spirulina. Se observó que la deposición de proteína en músculo muestra un comportamiento similar al observado por Sicuro et al. (2017), en el que a mayor inclusión de GM en la dieta, menor deposición de proteína en el músculo. De acuerdo con De la Higuera et al. (1999) la síntesis y deposición de proteína en el músculo se altera en la anguila europea (Anguilla anguilla Linnaeus, 1758) cuando se alimenta con una dieta deficiente en aminoácidos esenciales. Se ha reportado (Gatlin et al., 2007) que el GM es deficiente en lisina, un aminoácido esencial, y que cuando se ha incluido en más de 75% como fuente de proteína en la dieta, afecta no solo la deposición de proteína, sino también el crecimiento. A diferencia de la relación entre el crecimiento y la deposición de proteína en el músculo, los contenidos de proteína en el hígado muestran el papel regulatorio de este órgano, pues las dietas con las inclusiones de GM muestran una mayor deposición de proteína a pesar de la deficiencia de aminoácidos esenciales, particularmente en los organismos alimentados con las dietas G75% y G100%.
El contenido de lípidos en el músculo fue mayor a los reportados por Aguillón et al. (2017) y Enríquez et al. (2015) en crías de trucha arcoíris alimentados con diferentes productos provenientes de la soya, así como los reportados por Pratoomyot et al. (2010) en crías de salmón del Atlántico (Salmo salar Linnaeus, 1758). Se observó una tendencia de mayor contenido de lípidos en el músculo conforme aumentó la inclusión del GM. Previamente, se ha reportado que la deposición de lípidos en el músculo disminuye cuando se utilizan dietas con proteínas de origen vegetal, principalmente con proteína proveniente de la soya (Pratoomyot et al., 2010; Aguillón et al., 2017). Hasta el momento, no existen reportes de la manera en que los altos niveles de GM afectan la deposición de lípidos en el músculo, pero se considera que los organismos alimentados con las dietas G75% y G100% depositaron una mayor cantidad de lípidos para utilizarlos como fuente de energía. Los contenidos de lípidos en el hígado son similares a los previamente reportados por Aguillón et al. (2017) y son considerados como normales.
El consumo de oxígeno y la excreción de N pueden ser utilizados para estimar la oxidación de nutrientes y, usualmente, valores más bajos indican un mejor uso de la fuente de proteína (menos excreción) y menos energía en su oxidación (menos consumo) (Bureau et al., 2002; Azevedo et al., 2004). La inclusión de GM causó un aumento en la excreción de N y en el consumo de oxígeno, lo cual indica una baja utilización de la fuente de proteína, particularmente en las truchas alimentadas con las dietas G75% y G100%, y que se ve reflejado en el crecimiento. Esto ya se ha observado previamente en la trucha cuando se utilizan proteínas de origen vegetal, particularmente con harina de soya (Cruz et al., 2011). Por otro lado, la inclusión de GM causó un decremento en la excreción de P, aunque no se observaron diferencias significativas con respecto al grupo control. Cabe señalar que el fósforo es un constituyente del endoesqueleto, los fosfolípidos, los ácidos nucleicos y las membranas celulares (Kaushik, 2011) y que, regularmente, su fuente más importante para el organismo proviene de la dieta. Bureau y Cho (1999) señalan que el fósforo disuelto es una de las mayores preocupaciones ambientales en las operaciones de producción acuícola, pues el aumento de este elemento en las descargas de las granjas afecta a los sistemas acuáticos que las rodean al aumentar la disponibilidad de nutrientes. El uso de GM en las dietas de trucha ayudaría en la reducción de las descargas de P.
El efecto de las fuentes de proteína en la dieta puede ser evaluado con la respuesta inmune (Kiron, 2012), pues de acuerdo con Hardy (2001), un consumo insuficiente afecta el contenido de proteína en las células, lo cual puede incapacitarlas en sus funciones. Esto es particularmente crítico cuando se utilizan proteínas de origen vegetal (Tacon, 1995), por lo que la evaluación de algunos parámetros inmunológicos permite entender con mayor profundidad el efecto del GM en la trucha. De acuerdo con Scapigliati (2013), los linfocitos son células relacionadas con la respuesta inmune específica, particularmente en la producción de inmunoglobulina inducida por un antígeno. En la presente investigación se determinó el número total de linfocitos y se observaron valores similares a los reportados previamente por Jalili et al. (2013) y por Bohlouli y Sadeghi (2016) en juveniles de trucha arcoíris, excepto en el grupo alimentado con la dieta G50%, que mostró valores significativamente más altos. De acuerdo con Ye et al. (2013), el aumento en el conteo del número total de linfocitos indica una exposición a un patógeno. Durante la prueba, particularmente al final, no se observaron signos de alguna infección en los juveniles alimentados con la dieta G50%. Existe la posibilidad de que la infección estuviera en sus inicios al momento de tomar las muestras, pero no se debió al tratamiento con el GM.
La actividad explosiva es uno de los mecanismos de acción de los macrófagos, en la cual se forman especies de oxígeno reactivos, como anión superóxido y peróxido de hidrógeno (Rubio-Godoy, 2010). Esta prueba se utiliza como un indicador de cómo las dietas afectan las respuestas inmunológicas de los organismos. Las actividades de los macrófagos de los diferentes grupos reportadas en este trabajo son similares a los datos reportados por Aguillón et al. (2017) para juveniles de trucha arcoíris y a los de Taoka et al. (2006) en tilapia (Oreochromis niloticus Linnaeus, 1758). Aunque previamente Burrells et al. (1999) reportaron que la inclusión progresiva de proteínas de origen vegetal afecta la actividad de los macrófagos, no se observaron diferencias significativas entre los tratamientos ni tampoco respecto al control, por lo que la actividad se considera dentro de los límites normales, ya que el GM no alteró esta repuesta en los 90 días de alimentación.
La presente investigación muestra que el GM puede utilizarse en una sustitución de hasta el 50% de harina de pescado en dietas para juveniles de trucha arcoíris, sin afectar el crecimiento, la deposición de nutrientes (proteína y lípidos) en músculo y en hígado, ni la actividad explosiva de los macrófagos. La dieta G50% también disminuye la excreción de P en los organismos. La inclusión del GM en las dietas también podría tener un impacto en el costo de las dietas, pues es comparativamente más barato que la harina de pescado.

