











 nova página do texto(beta)
nova página do texto(beta) Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO 
 Permalink
PermalinkIntroducción
La familia Goodeidae es un grupo de peces de agua dulce distribuidos principalmente en la mesa central de México. Se le considera como “tesoro natural” porque tiene adaptaciones morfológicas asociadas a sus atípicas estrategias de reproducción y desarrollo embrionario (Domínguez & Pérez, 2007). Está compuesta por 45 especies vivíparas divididas en dos subfamilias Empetrichthynae y Goodeinae. Dentro de la segunda subfamilia, dos especies se consideran extintas, ocho en peligro de extinción y cuatro amenazadas (Domínguez & Pérez, 2007). La familia tiene una amplia distribución a lo largo de la vertiente del Pacífico mexicano, incluida la cuenca Lerma-Grande de Santiago, el río Ameca, el lago Magdalena y las cuencas de Balsas y Armería (Domínguez et al., 2008; Miranda et al., 2010). Dentro de esta subfamilia se encuentra Goodea atripinnis (Jordan, 1880), que es un pez omnívoro cuyo tamaño va desde pequeño -longitud estándar (LS) de 50 mm- hasta mediano -150 mm de LS- (Webb et al., 2004) y una relación peso-longitud de 5.2 a 13 (Miranda et al., 2009). Una de las características más importantes de este pez es que sólo se localiza en el estado de Aguascalientes, pero su población se está reduciendo drásticamente y se encuentra en un estatus de amenazada (Martínez & Rojas, 2008), por lo que se están implementando programas de reproducción ex situ (Silva-Santos et al., 2016). Sin embargo, para concretar estos proyectos de conservación es necesario encontrar planes de alimentación que permitan satisfacer las necesidades nutricionales de la especie durante sus diferentes etapas fisiológicas, debido a que se ignora la mayoría de los requerimientos nutricionales de esta familia. Así, el objetivo de este experimento fue evaluar el efecto de la inclusión de diferentes niveles de proteína en la dieta (25 a 45% de CP), la velocidad de crecimiento y otros parámetros productivos de G. atripinnis.
Material y métodos
Colecta y reproducción de organismos. Se realizó una colecta de organismos (G. atripinnis) en Potrero de los López, San José de Gracia, Aguascalientes (22° 07’ 13” N, 102° 30’ 19” O). Los organismos fueron recibidos en estanques y mantenidos en aislamiento por 20 días. Los peces se trataron con una solución de azul de metileno y NaCl y se mantuvieron en tanques con 20 individuos, bajo las siguientes condiciones: temperatura 20-22 °C, oxígeno disuelto 5-7 mg L-1, pH 8-9, alcalinidad 192 ppm y dureza del agua 88.5 ppm. Se utilizó un sistema cerrado de recirculación en todas las fases experimentales y se monitoreó la calidad del agua con un equipo Hass® en todos los acuarios. Se indujo la reproducción de los organismos colectados con temperatura y fotoperiodo (12 h de luz y 12 h de oscuridad) para obtener la primera generación. Estos organismos se utilizaron para el ensayo de crecimiento.
Ensayo de crecimiento. Se utilizó un diseño en bloques completamente aleatorio con cinco niveles de proteína, para lo cual se formularon dietas experimentales isoenergéticas pero con diferente nivel de proteína. Los niveles de inclusión calculados de este nutriente fueron de 250, 300, 350, 400 y 450 g kg-1 (Tabla 1). El análisis proximal (AOAC, 1990) de la dieta se presenta en la Tabla 1.
Tabla 1 Composición de dietas experimentales (base seca, %) para el pez Goodea atripinnis (Jordan, 1880).
| Nivel de proteína | (%) | ||||
|---|---|---|---|---|---|
| 25 | 30 | 35 | 40 | 45 | |
| Harina de pescado | 315 | 378 | 441 | 505 | 567 |
| Harina de soya | 147 | 176 | 206 | 235 | 263 |
| Aceite de pescado | 15 | 3 | 0 | 0 | 0 |
| Aceite de soya | 75 | 75 | 64 | 50 | 42 |
| Almidón | 403 | 323 | 244 | 165 | 83 |
| Gelatina | 5 | 5 | 5 | 5 | 5 |
| Premezcla mineral | 10 | 10 | 10 | 10 | 10 |
| Premezcla vitamínica | 10 | 10 | 10 | 10 | 10 |
| Carboxyl celulosa | 20 | 20 | 20 | 20 | 20 |
| Composición de aminoácidos calculada | (%) | ||||
| Metionina | 0.0104 | 0.0125 | 0.0146 | 0.0167 | 0.0188 |
| Lisina | 0.0303 | 0.0364 | 0.0425 | 0.0486 | 0.0545 |
| Arginina | 0.0323 | 0.0387 | 0.0452 | 0.0517 | 0.0580 |
| Treonina | 0.0183 | 0.0220 | 0.0256 | 0.0293 | 0.0329 |
| Leucina | 0.0327 | 0.0392 | 0.0458 | 0.0524 | 0.0588 |
| Isoleucina | 0.0211 | 0.0253 | 0.0296 | 0.0338 | 0.0379 |
| Valina | 0.0226 | 0.0271 | 0.0317 | 0.0363 | 0.0407 |
| Histidina | 0.0114 | 0.0137 | 0.0160 | 0.0182 | 0.0205 |
| Composición proximal | |||||
| Proteína cruda (%) | 25.05 | 29.99 | 34.34 | 38.99 | 43.49 |
| Grasa (%) | 15.41 | 15.03 | 15.06 | 15.56 | 15.96 |
| Fibra cruda (%) | 0.42 | 2.13 | 3.66 | 5.31 | 7.62 |
| Cenizas (%) | 8.5 | 9.11 | 11.56 | 13.71 | 15.19 |
| Extracto libre de nitrógeno (%) | 50.55 | 46.42 | 33.54 | 27.827 | 19.71 |
*Rovimix® Premix, DSM Nutritional Products Colombia S. A.: vit A 7.5x106 UI, vit. D3, 3.75x106 UI, vit. E 10.83 g, vit. B1 1.83 g, vit. B2 2.91 g, vit. B6 1.83 g, ácido pantoténico 8.33 g, biotina 166.66 mg, ácido fólico 833.33 mg, ácido ascórbico 41.66 g, niacina 7.5 g, vit. B12 3.33 mg, vit. K3 1.66 g, magnesio 91.66, zinc 21.66, hierro 28.33 g, cobre 2.5, yodo 0.17, selenio 66.66 g, manganeso 2.5 g, inositol F.G. 58.33 g, luctanox E 25 g.
Cada dieta experimental fue asignada aleatoriamente a tres tanques (bloques) de 20 L que contenían 20 peces (peso medio: 0.47 ± 0.02 g) por tanque. Los peces fueron alimentados a saciedad una vez al día durante 165 días. Durante ese periodo se registraron el consumo diario de alimento, los cambios en peso y longitud cada 15 días, así como el peso y longitud final de cada pez. La ganancia total de peso se estimó como la diferencia entre el peso final e inicial de cada tanque dividida entre el número de peces que contenía.
Los parámetros productivos se evaluaron utilizando las fórmulas empleadas por Bonaldo et al. (2015).
Las estimaciones incluyeron:
-
Tasa de crecimiento específico (TEC; % día-1):
TEC, % d-1 = (100 x (lnPVF-lnPVI)) / días
Donde PVF y PVI representan los pesos promedio finales e iniciales de cada tanque, respectivamente, y días se refiere al número de días que duró el experimento (150).
-
Tasa de consumo de alimento (TCA) (% día-1).
TCA % día -1 = 100 x AtPmD
At = alimento total proporcionado
Pm = peso promedio
D = número de días
-
Conversión alimenticia (CA)
CA=AtGP
GP = ganancia de peso.
-
Eficiencia proteínica (EPr)
EPr=GPCPc
CPC = consumo de proteína cruda
-
Factor de condición de Fulton (K)
K=Px100xL-3
P = peso
L = longitud
El peso de los peces se utilizó para estimar los parámetros del modelo de Gompertz:
Dónde:
Y |
= peso del organismo en el tiempo t |
𝑎 |
= peso maduro calculado, que se obtiene cuando la curva se convierte en asintótica t va al infinito |
b |
= peso alcanzado cuando ocurre la inflexión de la curva |
c |
= tasa de crecimiento |
t |
= edad en días |
Análisis estadístico. Para realizar los análisis estadísticos, los datos fueron transformados utilizando su logaritmo natural, pero los resultados se presentan en su forma original para facilitar su interpretación biológica. Después de la transformación, la normalidad de las variables se evaluó mediante la prueba de Shapiro-Wilk, y la homogeneidad de las varianzas de los resultados se analizó con la prueba de Bartlett. Los resultados del experimento se analizaron de acuerdo con un diseño completamente al azar (usando el acuario como repetición), y la interacción repetición por tratamiento se utilizó como término de error. El análisis de varianza se realizó a través de un diseño de mediciones repetidas considerando el nivel de proteína dietética como tratamiento. Los datos se analizaron con el programa JMP 7. Cuando se encontraron diferencias significativas, se realizó una prueba de Tukey para comparar las medias, que se consideraron diferentes con un nivel de significancia igual a 0.05.
Resultados
Se encontraron diferencias tanto en el peso final como en la ganancia de peso total (p<0.05) entre los niveles altos (45 y 40%) y bajos de proteína (Tabla 2). Este aumento representó 18% de diferencia entre la dieta más baja y la más alta. Como resultado del incremento en la ganancia de peso, la tasa específica de crecimiento aumentó en el tratamiento con mayor nivel de proteína. Este incremento en la velocidad de crecimiento proviene de un aumento significativo (p< 0.05) en la tasa de consumo y el aumento de la ingesta total de alimento. Como resultado, la conversión alimenticia (CA) se redujo ligeramente, pero la eficiencia de utilización de la proteína (EPc) y el factor de condición corporal (K) mejoraron.
Tabla 2 Índices biológicos de Goodea atripinnis (Jordan, 1880), en crecimiento alimentados con dietas con diferente nivel de proteína
| Índices biológicos | 250 | 300 | 350 | 400 | 450 | EEM |
|---|---|---|---|---|---|---|
| Peso inicial (g) | 0.469 a | 0.476 a | 0.469 a | 0.472 a | 0.472 a | 0.013 |
| Peso final (g) | 3.50c | 3.64 bc | 3.70bc | 3.77b | 4.23 a | 0.058 |
| GPT (g) | 3.01c | 3.15 bc | 3.21 bc | 3.28b | 3.74a | 0.045 |
| TEC (%d-1) | 1.344b | 1.359b | 1.379ab | 1.388ab | 1.461a | 0.021 |
| TCA (%d-1) | 1.01b | 1.02b | 1.03b | 1.02b | 1.10 a | 0.007 |
| Consumo total de alimento (g) | 48.71b | 48.61b | 49.93b | 50.88b | 56.75 a | 0.041 |
| CA (Kg/kg) | 0.83 a | 0.77b | 0.77b | 0.76b | 0.75b | 0.005 |
| EP | 4.80 a | 3.63b | 3.67c | 3.23d | 2.92e | 0.021 |
| K | 1.18 | 1.11 | 1.10 | 1.03 | 0.88 | 0.021 |
GPT = ganancia de peso total; TEC = tasa específica de crecimiento; TCA = tasa de consumo de alimento; CA = conversión alimenticia; PER = eficiencia proteínica; K = condición corporal SEM = error estándar de la media.abc Medias con diferente literal en el mismo renglón son diferentes (p<0.05).
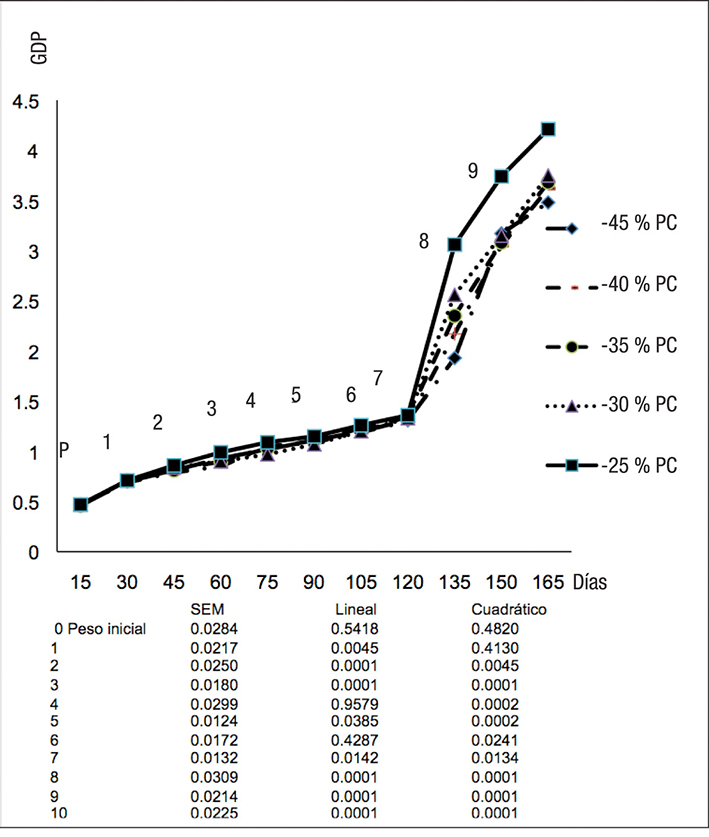
El modelo de Gompertz (Tabla 3) mostró que el peso final aumenta en forma significativa (p < 0.05) con el nivel alto de proteína, mientras que la velocidad de crecimiento se redujo de 0.016 a 0.015, lo cual no puede tener grandes implicaciones biológicas, puesto que el punto de inflexión b se alcanzó con el mismo peso en todos los casos. Sin embargo, el análisis de mediciones repetidas (Figura 1) mostró que, con los niveles altos de proteína, el peso al punto de inflexión se alcanza a una edad más temprana y permite un mayor desarrollo de los peces, lo que explica el mayor peso a la madurez obtenido en el modelo de Gompertz.
Tabla 3 Efecto del nivel de proteína en los parámetros del modelo de Gompertz describiendo los cambios de peso de Goodea atripinnis (Jordan, 1880).
| Nivel de proteína (g/Kg MS)/ Parámetros | 250 | 300 | 350 | 400 | 450 | EEM |
|---|---|---|---|---|---|---|
| a (g) | 22.01b | 22.90ab | 23.95 ab | 24.75 ab | 26.88 a | 0.96 |
| b (g) | 1.21a | 1.2a | 1.23a | 1.22a | 1.22 a | 0.013 |
| c | .016a | .016 ab | .016ab | .015ab | .015b | 0.00001 |
| R2 | 0.95 | 0.96 | 0.95 | 0.96 | 0.94 | |
| p-value | 0.0001 | 0.0001 | 0.0001 | 0.0001 | 0.0001 |
a = peso calculado a la madurez, b = peso alcanzado en el punto de inflexión de la curva, c = tasa de crecimiento; EEM = error estándar de la media con diferente literal en el mismo renglón son diferentes (p< 0.05).
Discusión
En contraste con otros trabajos donde la alimentación ad libitum fue proporcionada durante dos o tres veces al día (Shiau & Lan, 1996; Tekínay & Davies, 2001) en éste sólo se administró el alimento una vez al día, lo cual pudo impactar en los resultados. No obstante, de acuerdo con Kulczykowska y Sánchez-Vázquez (2010), si el alimento se proporciona siempre a la misma hora, los peces se sincronizarán con dicho momento y desarrollarán una adaptación que se conoce como actividad anticipada a la alimentación. Dicha adaptación les permite a los peces acoplar sus ciclos circadianos a la alimentación y reduce el estrés que sufren como consecuencia de la falta de alimento durante el resto del día.
La respuesta de la GP al incremento de la proteína en la dieta fue similar a la reportada por Catacutan y Coloso (1995), quienes encontraron un comportamiento similar al evaluar el crecimiento de Barramundi (Lates calcalifer Bloch, 1790), peces que alcanzaron su mayor crecimiento con una dieta con 50% de PC. También Uscanga-Martínez et al. (2012) observaron el mejor peso final y la mejor tasa de crecimiento al utilizar dietas con 50% de PC en Petenia splendida (Günther, 1862); sin embargo, en esa misma especie, Arredondo-Figueroa et al. (2012) encontraron una respuesta similar cuando administraron dietas con 45% de PC, lo que sugiere que la interacción con la energía en la dieta y el tipo de fuente de proteína utilizadas en el alimento modifican el requerimiento de proteína (Catacutan et al., 2001). Thoman et al. (1999) y Chou et al. (2001) encontraron una relación entre el aumento de peso corporal y la cantidad de proteínas y lípidos incluidos en las dietas experimentales cuando evaluaron diferentes especies, como la corvina roja (Scianops ocellatus Linnaeus, 1766) y la cobia (Rachycentron canadum Linnaeus, 1766). En el caso la corvina roja, un análisis de regresión múltiple que incluyó el peso vivo del pez, el nivel de proteína de la dieta y la energía digestible de ésta, mostró que la tasa de consumo de alimento estaba influenciada, aparentemente, por el peso vivo del pez y no por el contenido energético de la dieta, por lo que recomendaron una dieta con 44% de proteína cruda y 9.2% de lípidos; mientras tanto, la cobia requirió una dieta con 44.5% de proteína cruda y 5.76% de grasa.
El aumento en la tasa específica de crecimiento con el nivel más alto de proteína se relacionó con el incremento en la tasa de consumo y la cantidad total de alimento consumido en una forma similar a la reportada por Luo et al. (2004), quienes en Epinephelus coioides (Hamilton, 1822) encontraron que el consumo de alimento aumentó con niveles altos de proteína, mientras que los niveles reducidos de este nutrimento la disminuyeron. A su vez, los cambios en las tasas de consumo en este trabajo fueron similares a los de Bonaldo et al. (2015), quienes evaluaron el efecto del nivel de proteína en el crecimiento de rodaballo (Psetta maxima Linnaeus, 1758). Estos autores mostraron que, al aumentar la inclusión de harina de pescado para incrementar el porcentaje de proteína en la dieta en una forma indirecta, se incrementó su palatabilidad (Bonaldo et al., 2015). Sin embargo, el aumento en el consumo voluntario de las dietas altas en proteína puede estar relacionado con una reducción de los carbohidratos no fibrosos (CNF; Tabla 1), los cuales, en humanos han sido asociados con una reducción en el consumo voluntario al activar los centros de saciedad en el cerebro (Benelam, 2009). También se ha demostrado que esta reducción de CNF se asocia en el pez dorado (Carassius auratus Linnaeus, 1758) con una disminución de la concentración del polipéptido Y, el cual se encarga de estimular el consumo de alimento, por lo que una menor concentración de este polipéptido reduce el consumo de carbohidratos (Narnaware & Peter, 2002).
La ingesta más alta de proteína redujo su eficiencia de utilización (EP) en forma similar a la reportada por Walter et al. (2010). En el trabajo de estos investigadores la EP disminuyó cuando aumentó el nivel de proteína en las dietas. Dicha reducción puede explicarse debido a que el exceso de proteína no puede almacenarse como tal, por lo que es desaminada mediante el ciclo de la urea carnitina y eliminada como urea (Ip et al., 2014). Esta desaminación permite utilizar los carbonos para formar ATP, que es aprovechado como fuente de energía en los músculos (Jia et al., 2017) y favorece el almacenamiento de energía en forma de ácidos grasos (Wu, 2009), por lo que la energía retenida en los peces alimentados con dietas ricas en proteínas fue significativamente mayor que en aquellos alimentados con niveles bajos. Esto se confirma en otros experimentos donde la EP aumentó cuando se disminuyó la proteína dietética al mismo nivel de lípidos (Lee et al., 2002; Kim & Lee, 2005; Kim et al., 2012). También debe considerarse que el uso de la proteína puede verse afectado por diversos factores, como el estado de desarrollo del pez, la fuente de proteína y, más significativamente, por el contenido energético, de ácidos grasos y azúcares en la dieta (Barreto-Curiel et al., 2016).
El factor de condición de Fulton permite describir la condición individual de un pez (Froeze, 2006), por lo que, diferentes valores de K indican diferencias en la calidad de las fuentes de alimento (Sarkar et al., 2013). Sin embargo, los resultados de este experimento mostraron que el factor de Fulton no se modificó estadísticamente, lo que sugiere que la relación peso-longitud de los peces se mantuvo uniforme entre tratamientos, aunque el nivel de proteína limitó el crecimiento.
Los modelos de crecimiento permiten establecer una relación entre el peso y la edad a la madurez; es decir, permiten predecir el tamaño máximo del animal en un tiempo dado y se han establecido como un requisito previo para la determinación de los requerimientos nutricionales de una especie (Lima et al., 2014). Dentro de los modelos con mayor precisión, en algunos tipos de peces se encuentra el modelo de Gompertz (Lima et al., 2014). Bajo las condiciones de este trabajo, este modelo mostró (Tabla 3) que con el nivel alto de proteína el peso final aumenta en forma significativa (p <0.05), mientras que la velocidad de crecimiento se reduce de 0.016-0.015, lo cual no tiene grandes implicaciones biológicas, y el punto de inflexión b se alcanza con el mismo peso en todos los casos. No obstante, el análisis de mediciones repetidas (Figura 1) muestra que con los niveles altos de proteína el peso al punto de inflexión se alcanza a una edad más temprana y permite un mayor desarrollo de los peces, lo que explica el mayor peso a la madurez obtenido con el modelo de Gompertz. Este modelo se ha utilizado para describir el crecimiento de otros peces, como la tilapia del Nilo (Oreochromis niloticus, Linnaeus, 1758), y ha mostrado un mejor ajuste que otros modelos (Lima et al., 2014). Los cambios en el punto de inflexión antes de alcanzar la mitad del peso maduro en la fase lineal de crecimiento, confirman que la reducción de la proteína dietética para Goodea atripinnis retrasará el tiempo para alcanzar el peso maduro, pero con los niveles utilizados el crecimiento no se compromete. Por lo tanto, se puede concluir que es posible alimentar a los peces de la especie G. atripinnis en fase de crecimiento con niveles de proteína entre 40 y 45% y el uso de los niveles más altos de proteína resultará en organismos de mayor tamaño.

