











 nueva página del texto (beta)
nueva página del texto (beta) Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroducción
La búsqueda de combustibles alternativos y de bajo impacto ambiental es cada vez más importante debido a factores económicos y medioambientales. La naturaleza no renovable de las reservas de combustibles fósiles y el cambio climático han suscitado preocupaciones sobre la seguridad energética, generando interés en la utilización de energías renovables como los biocombustibles (Acosta et al., 2009). Estudios previos han demostrado la capacidad de algunos alimentos (aceite de soya y palma) para la producción de biodiésel, sin embargo, existe el debate sobre la factibilidad de utilizarlos para la obtención de energía, ya que podría ser una competencia directa con el uso recursos para alimentación humana y animal. En este sentido, en México la Ley de Promoción y Desarrollo de Bioenergéticos establece las normas en temas de producción y comercialización de insumos y permisos para el uso de maíz en los bioenergéticos.
El biodiésel ha surgido como una alternativa viable para sustituir el diésel derivado del petróleo (Taher et al., 2014), puesto que presenta varias ventajas por su biodegradabilidad y mínima toxicidad, además, su combustión produce menores emisiones de sulfatos, compuestos aromáticos, dióxido de carbono, monóxido de carbono, humo; hay más oxígeno libre que conduce a una combustión completa y emisiones reducidas (Atabani et al., 2012).
En un estudio realizado en el 2006 por la Secretaria de Energía (SENER) de México se menciona que la producción de biodiésel a escala comercial puede ser factible a mediano plazo si se llevan a cabo acciones integrales, que incluyan aspectos técnicos, económicos y medioambientales con el sector agrario y agroindustrial, conjuntando esfuerzos en investigación y desarrollo tecnológico. Aca-Aca et al. (2009) aluden que México requiere 10 plantas industriales con capacidad de 100,000 ton/año cada una, sólo para sustituir el 5% del diésel de petróleo usado cada año y recomiendan que la producción y el procesamiento se realicen con tecnologías diseñadas y construidas en el propio país. Taher et al. (2014) argumentan que el uso de biodiésel reduciría en un 45% las emisiones de hidrocarburos, 47% de CO2, y hasta un 66% las emisiones de partículas al ambiente.
En este sentido, los ácidos grasos de cadena larga obtenidos a partir de biomasa renovable (aceites vegetales, grasas animales y aceites de microalgas) representan la principal materia prima para la producción de biodiésel líquido obtenido en forma de alquil-ésteres de alcoholes de cadena corta como etanol y metanol (Robles-Medina et al., 2009). Los procesos más usados para la obtención de biodiésel son la pirólisis y la transesterificación, sin embargo, en el primer caso el método es caro y ofrece rendimientos bajos, mientras que el segundo se presenta como el método más viable para la obtención de biodiésel (Gog et al., 2012). Este proceso de conversión de aceites a biodiésel es necesario debido a que los aceites vegetales o extraídos de microalgas presentan una alta viscosidad y baja volatilidad, causando una combustión incompleta y la disposición de depósitos de carbón (Akho et al., 2007).
La obtención de biodiésel a partir de microalgas lleva más de 20 años de investigación y, en la última década, este tema ha retomado vital importancia, especialmente en los sectores privados y académi cos. Actualmente, investigadores de todo el mundo han demostrado el potencial de varias especies de microalgas (p. ej. Chlorella minutissima Fott et Novakova, Thalassiosira fluviatilis (Grunow) G. Fryxell et Hasle, Thalassiosira pseudonana Hasle et Heimdal, etc.) como materia prima para producir biodiésel; no obstante, los costos de producción no han resultado competitivos en comparación con los obtenidos de recursos fósiles ($0.55 USD/L) (Dalmas-Neto et al., 2014; Guldhe et al., 2016). A pesar de estos retos, las microalgas son consideradas actualmente como una de las alternativas ingenieriles para la obtención de biodiésel por a su alto contenido de lípidos y aceites (1 a 90% base seca, depen diendo de la especie y condiciones de cultivo), además de presentar una rápida producción de biomasa en comparación con otros cultivos energéticos (Mathimani et al., 2015; Sibi et al., 2015) y su huella eco lógica es mínima en un análisis de ciclo de vida (Atabani et al., 2012). Adicionalmente, las microalgas pueden reducir la cantidad de gases de efecto invernadero y consumir otros contaminantes (Halim et al., 2012). El objetivo del presente trabajo fue revisar la situación actual del uso de microalgas para la producción de biodiésel, incluyendo métodos de procesamiento, especies más utilizadas y sus principales ventajas en comparación con otros biocombustibles. De igual manera, se hace un análisis de los avances biotecnológicos actuales en este campo.
Microalgas como fuente de biodiésel: avances y perspectivas
Las microalgas son microorganismos unicelulares, se clasifican como procariotas y eucariotas. Se dividen en diferentes grupos en función de su taxonomía, incluyendo azul-verde, verde, verde-amarillo, rojo, marrón y las algas de oro. Hay más de 50.000 especies de microalgas (Rashid et al., 2014).
El empleo de microalgas para la producción de biodiésel es una alternativa viable por su elevado contenido de lípidos y su perfil idóneo para la obtención de éste. Aunado a lo anterior, otros atributos de las microalgas son su elevada eficiencia fotosintética, su capacidad de crecer tanto en aguas marinas, dulces y residuales, así como su velo cidad de crecimiento relativamente alta (Garibay et al., 2009). Las microalgas pueden mitigar las emisiones de CO2 y producir lípidos, por lo que se consideran con potencial para la obtención de biocombustibles de tercera generación (Arias et al., 2013).
Es importante el estudio de factores, tales como el aislamiento de especies de microalgas, el mecanismo metabólico, las condiciones de cultivo y el modo de operación y el diseño del fotobiorreactor, ya que son aspectos determinantes para mejorar el desarrollo del biocom bustible (Zhu et al., 2013).
También se debe desarrollar un proceso sustentable de producción de biodiésel a partir de microalgas, que sea técnica y económicamente viable, donde se haya realizado la optimización de medios de cultivo, selección, manipulación de cepas y el diseño adecuado de fotobiorreactores (Arudchelvam & Nirmalakhandan, 2013). Además, se debe considerar el proceso de recuperación de biomasa, extracción de aceites, subproductos, tecnología de transesterificación, purificación y uso de subproductos (Dai et al., 2014).
El uso de microalgas cultivadas para la obtención de lípidos ha demostrado un alto rendimiento 59 m3 ha-1 año-1 (Lee et al., 2012), lo cual la convierte en una fuente potencial para la obtención de biodiésel. La gran mayoría de las especies conocidas de microalgas acumulan sus ácidos grasos, triglicéridos y glicolípidos de forma intracelular, con excepción de Botryococcus braunii Kuetzing, esto involucra procesos de extracción de lípidos que encarecen la producción de biodiésel (Lee et al., 2012). En cálculos recientes se estima que el costo de producción de biomasa de microalgas es elevado, alrededor de $5.8 USD Kg-1, y a su vez también son altos los requerimientos energéticos (33 MJ Kg-1 de biomasa seca) para la recuperación de los lípidos intracelulares. En contraparte, por ejemplo, el aceite de canola está disponible por un monto de $1.25 USD Kg-1, lo que demuestra que actualmente los precios del aceite extraído de microalgas no son competitivos para la obtención de biodiesel (Slade & Bauen, 2013).
La selección de microalga es el primer paso en el desarrollo de un proceso de producción, éstas deben tener las características adecuadas para las condiciones de cultivo específicas, con el fin de conseguir un determinado producto (Jaimes-Duarte et al., 2012). En la Tabla 1 se presenta el porcentaje en peso de lípidos de algunas microalgas, así como su productividad en lípidos y biomasa.

En este sentido, Slade & Bauen (2013) mencionan que los costos de la producción de biomasa microalgal se pueden reducir cultivando las microalgas en lagunas abiertas, puesto que los gastos que se requieren para la operación del sistema (mantenimiento y materias primas) son menores, en comparación al cultivo en fotobiorreactores, principalmente por la inversión para la compra del sistema. Señalan que otro aspecto que reduce significativamente el precio de producción (˃50%) es bajar los costos del CO2, agua y nutrientes. La optimización y diseño de equipos para el procesamiento postcultivo también representa un ahorro significativo para la obtención de biodiésel a partir de microalgas. Bagchi et al. (2015) destacan que el proceso de secado representa hasta un 30% del costo total, por eso diseñaron un horno de secado que ahorra el 50% (0.017 kWh) de la energía generalmente usada en este proceso.
Alva et al. (2013) sostienen que el uso de aguas residuales municipales para la producción de biomasa microalgal ofrece una alternativa viable para disminuir los costos de producción de lípidos debido al aho rro de agua y nutrientes (ej. N, P, K), estos últimos se pueden controlar mediante diluciones y si son necesarias concentraciones mayores se pueden suplementar con sales de bajo costo u otros efluentes anaero bios ricos en estos nutrientes (ej. N y P). Esta alternativa contribuye en la recuperación de nutrientes presentes en aguas residuales.
Existen otros estudios que han buscado soluciones para reducir costos en el cultivo de microalgas utilizando fuentes alternativas de carbono y otros nutrientes. En este sentido, Tripathi et al. (2015) infor maron que la búsqueda nuevas especies con capacidad de fijar altas concentraciones de CO2 (15%) o utilizar el carbono presente en las ro cas (como el mármol) representa un potencial para la obtención de biomasa microalgal útil para la formulación de biodiésel con menores costos. Rattanapoltee & Kaewkannetra (2014) apuntan que el uso de sacarosa en bajas concentraciones para la acumulación de lípidos en Chlorella vulgaris Beijerinck resulta ser el sustrato más económico en relación costo-beneficio. Chi et al. (2016) indican que el costo del ni trógeno requerido para el crecimiento microalgal debe ser considerado en los procesos de escalamiento para la producción de biodiésel. Estos autores reportaron un 66.5% de acumulación de lípidos en Isochrysis zhangjiangesis Hu et Liu y una productividad de lípidos de 3.28 g/d por cada gramo de nitrógeno alimentado al cultivo, lo cual consideran un uso eficiente del nitrógeno.
Biosíntesis de ácidos grasos saturados, insaturados y triglicéridos en microalgas
Las rutas metabólicas sobre la biosíntesis de lípidos de microalgas son un campo todavía por explorar en comparación con el metabolismo de las plantas. Sin embargo, se cree que las rutas metabólicas seguidas son similares (Yu et al., 2011). La síntesis de ácidos grasos en las algas se realiza en los cloroplastos y generalmente se producen cadenas de ácidos grasos de 16C y 18C, los cuales son usados por la célula para la síntesis de membranas, triglicéridos, almacenamiento de lípidos neu tros y cloroplastos. Específicamente, la biosíntesis de ácidos grasos se inicia con la conversión del Acetil-CoA a Malonil-CoA catalizada por la enzima Acetil-CoA Carboxilasa (ACCasa). La molécula de Malonil-CoA es la donadora de carbono para la síntesis de nuevos ácidos grasos, donde es transferida de la CoA a un cofactor proteico (ACP) para la ob tención del Malonil-ACP, el cual participa en una serie de reacciones de condensación con los aceptores acil-ACP y acetil-CoA. El ciclo termina cuando el grupo acil es removido del sitio ACP por la acil-ACP tioestera sa y una vez sintetizados los ácidos grasos libres son trasportados del ACP al glicerol-3-fosfato o al monoacilglicerol-3-fosfato (Ohlrogge & Browse, 1995). La ruta metabólica se muestra en la Figura 1.
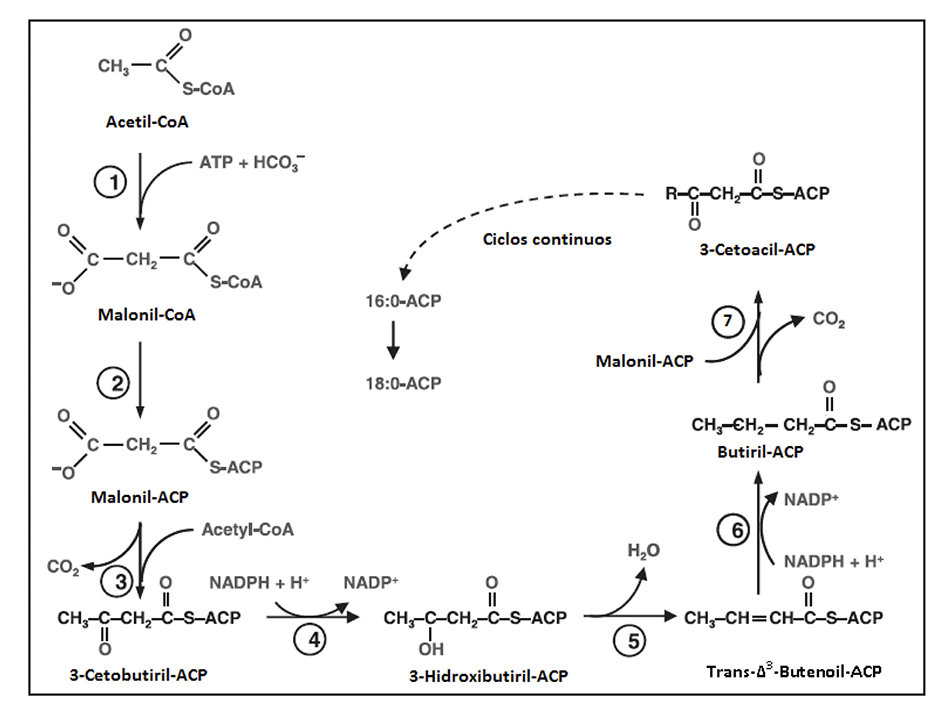
Figura 1 Bioquímica de síntesis de lípidos por microalgas. Adaptada de Hernández et al., 2009. *Reacción 1: El Acetil-CoA entra al ciclo como sustrato para la enzima ACCasa y se obtiene el Malonil-CoA. Reacción 2: es catalizada la conversión de Malonil-CoA a Malonil-ACP por la Malonil-CoA: ACP tranferasa (el Malonil-CoA será la molécula donadora de carbón en las siguientes reacciones de elongación). Reacción 3: descarboxilación del Malonil-CoA y la condensación con el Acetil-CoA. Reacción 4: reducción del 3-Cetobutiril-ACP por la 3-cetoacil-ACP reductasa Reacción 5: deshidratación para la obtención de una instauración en el carbono α por la enzima 3-hidroxiacil-ACP deshidrasa. Reacción 6: reducción de la doble ligadura por la enoil-ACP reductasa. Reacción 7: ciclo de elongación (repetición del paso 3 al 6) hasta la obtención de ácidos grasos saturados de 16C o 18C o ambos.
Así mismo, la producción de ácidos grasos poliinsaturados (PUFAs) a base de microalgas marinas o de agua dulce es de vital importancia para la elaboración de biodiésel o como aditivos alimentarios. Las microalgas tienen mayor capacidad para sinterizar y almacenar PU FAs de cadena larga ˃18C (ácido eicosapentanoico (EPA), C20:5ω3, ácido docosahexanoico (DHA), C22:6 ω3 y ácido araquidónico, C20:4 ω6) (Harwood & Guschina, 2009). De igual forma, Guschina & Harwood (2006) mencionan que existen principalmente dos rutas metabólicas para la síntesis de EPA denominadas ω-3 y ω-6 aunque la ω-6 es la ruta más usada. En la ruta ω-6 el ácido linoleico es desaturado para la obtención de ácido γ-linolénico, después es elongado para conseguir el ácido dihomo-γ-linolénico y su posterior desaturación para producir el ácido araquidónico y finalmente el EPA. En la ruta ω-3 el ácido linoleico es desaturado, lo que resulta en el ácido α-linolénico, para enseguida ser desaturado y generar el ácido estearidónico, a continuación, se realiza una elongación y se obtiene el ácido eicosatetranoico y finalmente su conversión a EPA. Las rutas biosintéticas para el resto de los ácidos grasos se muestran en la Figura 2, ahí las desaturaciones se abreviaron con D y el número adjunto muestra el sitio de reacción (E, elogación; PKS policétido sintasa). Las líneas punteadas indican reacciones que no han sido descritas completamente.
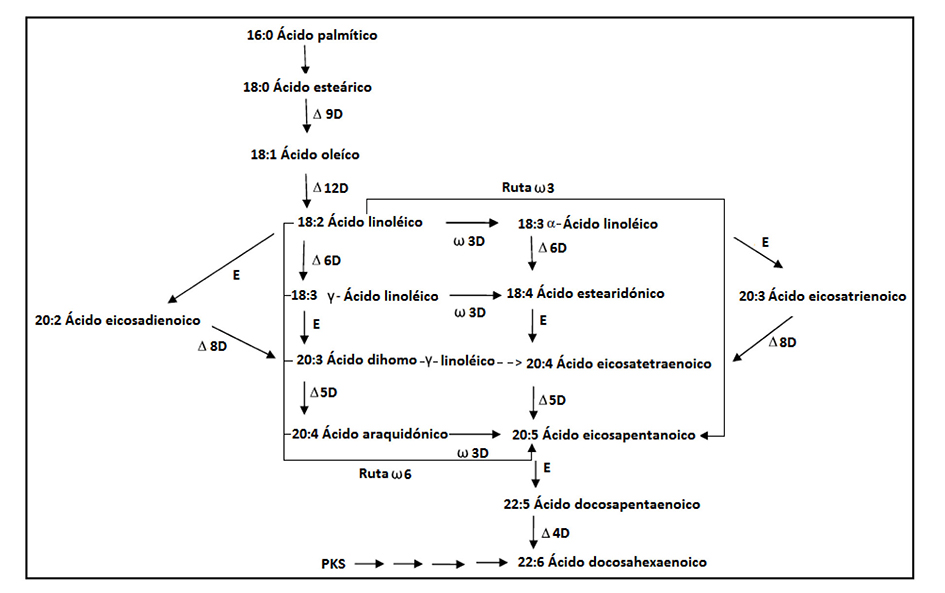
Figura 2 Biosíntesis de ácidos grasos poliinsaturados en algas eucariotas. Adaptado de Harwood y Gushina, (2009).
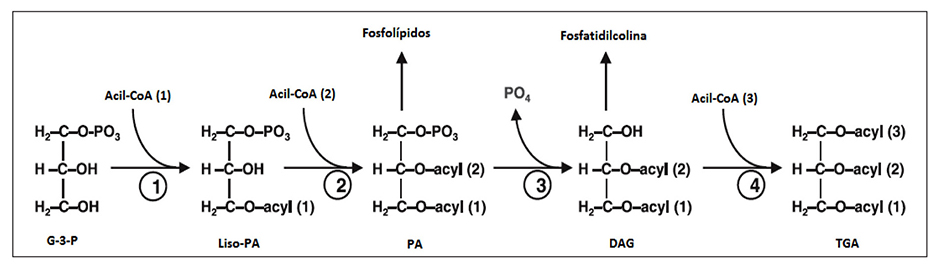
Los triglicéridos son comúnmente encontrados como grasas almace nadas o aceites y pueden ser neutros. Están constituidos de tres ácidos grasos esterificados vía grupos hidroxilos de un grupo glicerol. En la síntesis de triglicéridos las microalgas usan la ruta directa de glice rol (ver Figura 3) (Hu et al., 2008). Específicamente, los ácidos grasos producidos en los cloroplastos secuencialmente son transferidos por la CoA a la posición 1 y 2 del glicerol-3-fosfato, obteniendo el ácido fosfatídico (PA) como metabolito central. La desfosforilación del PA catalizado por la fosfatasa libera diacilglicerol y finalmente un tercer ácido graso es unido a la posición 3 libre del DGA mediante la enzima diacilglicerol aciltranferasa. Además, las moléculas PA y DGA pueden ser usadas directamente en la síntesis de lípidos polares como la fos fatidilcolina (PC) y galactolípidos (Yu et al., 2011).
Técnicas para la obtención de biodiésel a partir de microalgas
El proceso de obtención de biodiésel a partir de microalgas se divide en 3 etapas: 1) cultivo de la cepa para la obtención de biomasa, 2) cosecha de biomasa y disrupción celular y 3) reacción de esterificación para la conversión a biodiésel (Koller et al., 2012).
Etapa 1: métodos para aumentar la biomasa y lípidos. Reciente mente se han desarrollado nuevas estrategias apoyadas con técnicas biotecnológicas con el fin de mejorar los rendimientos y reducir conta minantes (p. ej. fitorremediación, biopelículas para filtración, bioadsor bentes). Sin embargo, el alto costo del carbono heterotrófico (p. ej. glu cosa) usado como sustrato representa la principal barrera económica para la producción de lípidos a partir de microalgas (Wang et al., 2014). En este sentido, Mu et al. (2015) señalaron que con el uso del bagazo hidrolizado de caña de azúcar para el cultivo de Chlorella protothecoi des (Krueger) Kalina et M. Puncoch en condiciones heterotróficas se alcanzó una concentración de biomasa de 24.01 g/L y una productivi dad lipídica de 1.19 g/L/d, y suponen que la xilosa y la arabinosa invo lucradas en el ciclo de las pentosas fosfato pueden estar predominando sobre la ruta de la glucólisis mejorando la acumulación de lípidos. De igual forma, Cho et al. (2015) observaron que el uso de mioinositol en medios de cultivo a una concentración de 500 mg/L aumentó un 48% la producción de biomasa y un 30% la acumulación de lípidos en Dunalie lla salina Ben-Amotz et Avron, también se incrementó la producción de ácido linoleico, linolénico y linolelaídico. Por otro lado, Gui et al. (2014) aplicaron un tratamiento de hidrólisis ácida a los residuos lignocelulósicos de la planta Glycyrrhiza uralensis con el objetivo de generar un sustrato para el cultivo heterotrófico de Chlorella protothecoides, lo que incrementó en un 62% el rendimiento de lípidos y biomasa.
Se ha demostrado que mantener a la microalga bajo condiciones de estrés incrementa la acumulación de lípidos, por lo que se han utilizado métodos como limitación de nitrógeno y adición de iones metálicos y sales como EDTA, para inducir las condiciones de estrés (Ren et al., 2014; Liu et al., 2016). En este sentido, Liu et al. (2016) refieren que mediante una continua limitación de nitrógeno en condi ciones fotoautótrofas en cultivo por lote, C. vulgaris logra un rendimien to máximo de lípidos de 305.71 mg/L/d y una biomasa de 4.61 g/L. Morales-Sánchez et al. (2014) reportan que bajo limitación de nitró geno en condiciones heterotróficas de lote post-alimentado Neochloris oleoabundans (S.Chantanachat et H.C.Bold) J. Komarek acumula hasta un 53.8% de lípidos, con una concentración de biomasa de 20.9 g/L y una productividad de 1020 mg/L/d de lípidos. Por su parte, Ren et al., (2014) evaluaron el efecto de la adición de metales y sales, con lo que lograron un incremento del 28.2% de lípidos, 29.7% (275.7 mg/L/d) en la productividad y 3.49 g/L de biomasa bajo condiciones heterotróficas y mediante la adicción de Fe3+, Mg2+ y Ca2+ y EDTA en Scenedesmus sp. De manera general, se considera que la limitación de nitrógeno y el uso de lote alimentado bajo condiciones de estrés nutricional de nitrógeno ofrecen los mejores rendimientos de acumulación de lípidos y biomasa en cultivos de microalgas, esto debido a que la falta de nitrógeno redu ce la síntesis de proteínas y estimula la formación de lípidos.
El uso de técnicas modernas enfocadas en modificaciones gené ticas o la selección de cepas mutadas son algunas de las alternativas para aumentar los rendimientos de lípidos en microalgas y abaratar los costos de producción de biodiésel (Anandarajah et al., 2012; La Russa et al., 2012). Investigaciones previas demostraron que las mutaciones de los 3 tipos de genes homólogos DGATs tipo-2 (Acil-CoA: diacilglice rol aciltransferasa) asociados con la producción de triacilgleceridos no presentan un aumento significativo en la acumulación de lípidos, ade más, provocan alteraciones en el perfil de los ácidos grasos en Chla mydomonas reinhardtii P. A. Dangeard (La Russa et al., 2012). Por su parte, Xue et al. (2015) realizaron una modificación genética en Phaeo dactylum tricornutum Bohlin para mejorar la acumulación de lípidos neutros. La modificación se realizó en la enzima málica, involucrada en el metabolismo del piruvato y fijación de carbono. Los resultados mostraron un incremento del 57.8% en la producción de lípidos, man teniéndose una tasa de crecimiento celular similar a la cepa silvestre. En otra investigación, Zhang et al. (2016) bloquearon la enzima β-1-3 gluconasa, con el producto comercial Micafungin, inhibiendo la for mación decrisolaminarina y así fomentar la acumulación de lípidos en Isochrysis zhangjiangensis. Con esto lograron obtener un 50% más en el rendimiento de lípidos y una disminución del 10% de carbohidratos.
Etapa 2: técnicas de extracción de lípidos. No obstante el potencial biotecnológico que representan las microalgas en la producción de lí pidos útiles para generar biodiesel, actualmente no existen métodos a escala industrial factibles para la extracción de lípidos y solamente se han reportado nuevos métodos para su uso a escala laboratorio (Neto et al. 2013). Entre los métodos más utilizados para la extracción de lípidos de microalgas están el uso de solventes, fluidos supercríticos, campo eléctrico de pulso, microondas y ultrasonicación (Batista et al., 2016; Taher et al., 2014). En este sentido, el enfoque actual es desarrollar o mejorar los métodos de disrupción celular para disminuir el uso de sol ventes, producir menos residuos, aumentar los rendimientos y mejorar la calidad del producto final (preservar los PUFAs) (Hidalgo et al., 2016).
La sonicación asistida es uno de los métodos físicos más atractivos para romper células y facilitar la extracción de lípidos de microalgas. Una investigación realizada por Suarsini y Subandi (2011) indica que la ultrasonicación presenta mejores rendimientos de extracción en com paración con el método de soxhlet y maceración y aparte reduce los tiempos. Neto et al. (2013), por su parte, notificaron que la ultraso nicación asistida logra una extracción de lípidos del 40.3 y 39.5% en Thalassiosiria fluviatilis y Thalassiosiria pseudonana, respectivamente. Otro de los métodos recientemente más estudiados es el uso de fluidos supercríticos, donde en investigaciones como la de Nobre et al. (2013) emplean fluidos supercríticos para la extracción de lípidos y pigmen tos de Nannochloropsis sp. En su investigación señalan que el uso de etanol como cosolvente y CO2 supercrítico mejora los rendimientos de extracción, alcanzando los 45 glípidos/100 gbiomasa seca de lípidos y 70% de los pigmentos, también demostraron que la masa remanente puede uti lizarse para la producción de biohidrógeno mediante una fermentación con Enterobacter aerogenes Hormaeche et Edwards. Esta tecnología también ha sido aplicada en la extracción de ácidos grasos esenciales a partir de C. protothecoides, Scenedesmus obliquus (Turpin) Hegewald et Hanagata y Nannochloropsis salina D. J. Hibberd, logrando la re cuperación selectiva del ácido α-linolénico y ácidos grasos ω-3 en proporciones similares para las tres microalgas, con rendimientos de extracción de lípidos del orden del 18.15% (Solana et al., 2014).
En relación con el uso de solventes, investigaciones actuales se han enfocado en disminuir su uso y aumentar el rendimiento de ex tracción. Abomohra et al. (2016) resaltan que es de vital importancia optimizar la extracción de ácidos grasos esterificados porque son los únicos que se usan para la producción de biodiésel a partir de mi croalgas. En este sentido, mencionan que la relación 2:1 de solventes cloroformo: metanol durante 2 h muestran los mejores rendimientos de extracción de ácidos grasos esterificados, sin la necesidad de apli car procesos de ruptura celular cuando se trabaja con S. obliquus. Recomiendan un tratamiento previo con agua caliente por 5 min para inactivar las lipasas y aumentar hasta un 13.7% los rendimientos de extracción de ácidos grasos esterificados. De igual forma, Abomohra et al. (2016) mejoraron la extracción de lípidos de S. obliquus mediante la optimización de mezclas de solventes y tiempos de extracción aunados a un proceso de ruptura celular. Los resultados muestran que la rela ción óptima de solventes cloroformo: metanol fue 2:1 con un tiempo de extracción de 2 h sin necesidad de realizar lisis celular para la recupe ración de los ácidos grasos esterificados. En otro estudio, Hidalgo et al. (2016) ensayaron con mezclas de solventes polares y no polares para la extracción de lípidos esterificables de B. braunii demostrando que la mezcla cloroformo-metanol (75 % v/v metanol) fue la más eficiente, con un 98.9% de rendimiento de extracción de lípidos.
Una de las tecnologías emergentes prometedoras para la extrac ción de compuestos debido a su bajo costo y bajas temperaturas de operación, es la extracción por presurización cíclica. En esta técnica se utilizan presiones moderadas (˂1013 kPa), lo que reduce los costos en comparación con fluidos supercríticos, por ejemplo, e incluso menores que el uso de microondas o sonicación asistida (Ortiz et al., 2015; Ba tista et al., 2016). Batista et al. (2016) aplicaron esta tecnología para la extracción de lípidos de Chlorella pyrenoidosa H.Chick, demostrando que con bajos ciclos de presurización (300 s a 91.4 kPa + 300 s a 200 kPa por ciclo) por un tiempo de 7 h, en medio etanólico se alcanza un 72% de remoción de lípidos, con un rendimiento de 11.3% de biomasa. De forma similar, Pieber et al. (2012) realizaron la extracción del ácido eicosapentaenoico (EPA) de Nannochloropsis oculata Droop con fluidos presurizados y los resultados mostraron que el metanol fue el mejor solvente, comparado con el n-hexano, obteniendo un rendimiento del 16.7 ± 6% y 3.7 ± 0.1% de ácidos grasos totales y EPA, respectiva mente.
Otro de los métodos novedosos es el uso de pulsos eléctricos para la lisis celular y realizar extracciones in situ. Flisar et al. (2014) probaron un prototipo de generador de pulsos para la extracción de lípidos de microalgas y reportan un 22% de extracción de lípidos. Sin embargo, atribuyen que los rendimientos son bajos debido a problemas de recu peración y cuantificación de lípidos, ya que la ruptura celular se realizó directamente en el reactor. En otras investigaciones similares, como la de Seo et al. (2015) desarrollaron un método para romper la célula y extraer los lípidos usando UV en conjunto con la reacción de Fenton lo grando una eficiencia de remoción del 85% con bajas concentraciones de H2O2 (0.3%).
Etapa 3: métodos químicos usados en la reacción de transesterificación. La reacción de transesterificación es uno de los métodos más estudiados y por lo tanto más utilizados en la producción de biodiésel. En este proceso se da la transferencia de un grupo acilo, que puede suceder entre un éster y un ácido (acidólisis), un éster y otro éster (interesterificación) o entre un éster y un alcohol (alcohólisis) (Freire et al., 2011) (ver Figura 4).

El desarrollo de nuevos catalizadores que mejoren las condiciones de los procesos de esterificación, reduciendo los impactos ambienta les y la corrosión que causa el uso de catalizadores convencionales (NaOH, KOH y H2SO4), representa un reto para la sustentabilidad de los procesos de esterificación (Hwa et al., 2014a). En este sentido, Hwa et al. (2014a) probaron el metóxido de calcio (Ca(OCH3)2) como cata lizador en presencia de metanol y lípidos recuperados de N. oculata y señalaron un rendimiento del 92% a 60 °C, siendo este 22% superior al registrado con el catalizador de Mg-Zr. En otro análisis, Hwa et al. (2014b) formularon un catalizador mixto de óxido de calcio y magnesio (CaMgO) soportado en alúmina (Al2O3) con el cual se alcanzaron rendi mientos del 85.3% de transesterificación y un soporte estable y reusa ble. Macías-Sánchez et al. (2015) aplicaron un método de transesterifi cación directa de Nannochloropsis gaditana L. M. Lubin. El proceso de catálisis lo realizaron con metanol usando como catalizador cloruro de acetilo al 5% a 100 °C durante 105 min, y consiguieron una pureza del 82.7% de ácidos grasos metil-esterificados. De igual forma, Mathimani et al. (2015) mencionan que el H2SO4 al 3.5% por 2.4 h en presencia de metanol logra los mejores rendimientos para la reacción de metil-esterificación (60%), utilizado Chlorella sp. BDUG 91771.
Métodos biotecnológicos. Actualmente las lipasas usadas para la pro ducción de biodiésel son obtenidas principalmente de hongos (levadu ras y hongos filamentosos) y algunas se encuentran disponibles en el mercado (Novozym 435, Lyposime RM IM, Lypozime TL IM) (Yan et al., 2014). Estas lipasas presentan diversas ventajas, como el poco reque rimiento energético, la alta calidad de biodiésel producido y la casi nula generación de residuos (Meng et al., 2014). Aarthy et al. (2014) sostie nen que es posible aplicar procesos biotecnológicos para realizar una biocatálisis utilizando lipasas de levaduras como Candida antarctica (Goto, Sugiyama & Iizuka) Q. M. Wang, Begerow, F. Y. Bai & Boekhout, C. rugosa (H.W. Anderson) Diddens & Lodder, Crytococcus sp., Trichospo rum asahii Akagi ex Sugita, A. Nishikawa & Shinoda y Yarrowia lypolitica (Wick., Kurtzman et Herman) Van der Walt et Arx alcanzado rendimien tos de hasta 96% con bajas temperaturas. Por su parte, Aguierias et al. (2015) reportan la obtención de lipasas para la producción de bio diésel de hongos filamentosos como Rhizomucor miehei (Cooney et R. Emers.) Schipper, Rhizopus oryzae Went et Prins. Geerl., Thermomyces lanuginosus Tsikl, Aspergillus niger Tiegh y Penicillium expansum Link, y señalan que otra alternativa viable es utilizar las células completas para la biocatálisis.
Guldhe et al. (2016) proponen la obtención de biodiésel utilizando lipasas de A. niger como biocatalizadores alcanzando, un porcentaje de conversión del 53.76% a 35 °C en presencia de metanol. Sin embargo, estos rendimientos pueden considerarse bajos en comparación con los obtenidos con enzimas comerciales como Novozym 435. En virtud de ello, Navarro-López et al. (2015) dieron a conocer la elaboración de biodiésel a partir de Nannochlorpsis gaditina L. M. Lubin empleando la enzima comercial Novozym 435 a partir de 4 lipasas. La conversión alcanzada en este caso fue de 94.7% de ácidos grasos metil-esterifi cados (biodiésel), en un proceso de reacción de 48 h con 10 mL alcohol t-butanol/g de lípidos saponificables (LS), y una relación molar 11:1 metanol/LS. De igual forma, Castillo-López et al. (2015), utilizaron No vozym 435 con lipasas de C. antartica logrando un 92.6% de grado de esterificación, produciendo un biodiésel con 83% en peso de pureza a partir de N. gaditana. Las condiciones óptimas previamente expuestas se alcanzaron con una relación 1.5:1(metanol/ácidos grasos libres) y 15:1 (Novozym 435/ácidos grasos libres) a 25 °C durante 4 h. Es po sible deducir que la eficiencia de las enzimas comerciales Novozym 435 es similar (mayor al 90%) aun cuando se trabaja con diferentes especies de microalgas y bajo diferentes condiciones de extracción.
A pesar de los altos rendimientos de conversión de biodiésel usan do enzimas comerciales, se considera que la producción enzimática de biodiésel a base de microalgas no es factible debido al elevado costo de los procesos de aislamiento, purificación, inmovilización y a la baja estabilidad de las lipasas en medio etanólico (Ognjanovic et al., 2009), de modo que se ha trabajado en el desarrollo de métodos libres de solventes, lo cual evita riesgos de flamabilidad, toxicidad y reduce el costo por el uso de solventes y procesos de separación, además del desarrollo de soportes para enzimas económicos y biosustentables, que mejoren el rendimiento de producción de biodiésel (Li et al., 2014).
Surendhiran et al. (2014) desarrollaron un método para la obten ción de biodiésel a partir de Chlorella salina inmovilizando la levadura Rhodotorula mucilaginosa (A. Jorg.) F. C. Harrison ‘MTCC9737’ en un soporte a base del gabazo de la caña de azúcar en medio no alcohó lico. Los resultados mostraron un 85.2% de rendimiento de biodiésel con una relación 1:12 acetato de metilo/aceite a 40 °C durante 60 h. Duraiarasan et al. (2016), por su parte, diseñaron un soporte mediante nanopartículas magnéticas para la extracción y conversión directa de biodiésel a partir de los ácidos grasos obtenidos de C. salina Butcher. El método implementó la inmovilización de celulasas y lipasas con un rendimiento máximo de 93.56% con una eficiencia hasta por 10 ciclos, alcanzando resultados similares a los obtenidos con sistemas comer ciales.
Cultivo de microalgas. En general, las microalgas dependen para su crecimiento de un suministro de carbono y de una fuente de luz para realizar la fotosíntesis. Sin embargo, como respuesta a los cambios en las condiciones ambientales, pueden poner en funcionamiento diversos tipos de metabolismo (fotoautotrófico, heterotrófico, mixotrófico, foto heterotrófico). Al considerar el empleo de microalgas para la produc ción de biodiésel, es importante definir cuantitativamente la influencia de factores que intervienen en su crecimiento y que favorecen la pro ducción de lípidos, así como su interrelación para poder manipularlos en la obtención de biomasa con las características deseadas (Arias et al., 2013).
En la actualidad, la producción fotoautotrófica es la única técnica económicamente viable a gran escala para la producción de bioma sa (Borowitzka, 1997). Los dos sistemas más ampliamente utilizados son las lagunas abiertas y los fotobiorreactores cerrados (Borowitzka, 1999). La viabilidad técnica de cada uno de estos sistemas se ve in fluenciada por las propiedades intrínsecas de la cepa seleccionada, así como las condiciones climáticas y los costos por espacio y agua (Bo rowitzka, 1992).
Un fotobiorreactor es un reactor en el que células de plantas o algas fotótrofas se cultivan o se utilizan para llevar a cabo una reacción fotobiológica. Los sistemas de cultivo abierto son normalmente menos caros de construir y operar, más duraderos que los grandes reacto res cerrados y con gran capacidad de producción en comparación con los sistemas cerrados (Yen et al., 2014). En la Tabla 2 se muestran las ventajas y desventajas de los fotobiorreactores y las lagunas abiertas.

Biorrefinería: sistemas de doble propósito para el cultivo de microalgas
El concepto de biorrefinación es similar al de refinerías de petróleo en que múltiples combustibles y productos químicos se obtienen utilizando el petróleo crudo como material de partida. Biorrefinería, por lo tanto, se define como el procesamiento de biomasa sostenible para obtener energía, biocombustibles y productos de alto valor a través de procesos y equipos para la transformación de biomasa (Trivedi et al., 2015).
También es importante tener en cuenta que el costo de la producción de microalgas es alrededor del 90% de la producción de biodiésel, esto indica que los esfuerzos en el desarrollo y la investigación deben centrar se en reducir los costos de producción de microalgas (Cheali et al., 2015).
El costo de producir biodiésel puede ser teóricamente compensado por los ingresos generados por otros coproductos de la biomasa (Vo nortas & Papayannakos, 2014). Las microalgas contienen cantidades significativas de proteínas y carbohidratos, así como pequeñas cantida des de compuestos funcionales de alto valor, por ejemplo, astaxantina, carotenos, clorofila, ácidos grasos libres y ácido linoleico (Rashid et al., 2014). Cada uno de éstos puede ser utilizado para generar produc tos de interés industrial en una biorrefinería. Estudios recientes han concluido que la producción de biodiésel a escala industrial puede ser económicamente sostenible si la estrategia de la refinería se basara en la producción de biomasa (Wijffels et al., 2010).
En una biorrefinería, los lípidos crudos son fraccionados en com puestos de alto valor agregado y lípidos para biodiésel (acilgliceroles) (Yan et al., 2014). Si las especies de microalgas contienen un alto nivel de proteínas, la biomasa residual de la producción de biodiésel puede ser utilizada como alimentación de ganado. Si la especie contiene altos niveles de carbohidratos, la biomasa residual puede ser fermentada para producir bioetanol (Halim et al., 2012).
Por otro lado, en los últimos años se ha conceptualizado el tér mino “sistemas de doble propósito” para describir al conjunto de procesos encaminados en la reincorporación de nutrientes presentes en efluentes residuales en la producción de biomasa (Olguín, 2012; Robles-Pliego et al., 2015). La biomasa generada, de acuerdo con sus características, puede ser empleada para la obtención de productos de alto valor agregado y biocombustibles, principalmente, en el contexto de una biorrefinería.
Específicamente, en el desarrollo de sistemas de doble propósito en el contexto de biorrefinerías de microalgas, se puede optar por un diseño modular en donde se establezcan procesos y operaciones uni tarias bien definidas (Figura 5). Para lo anterior, deben considerarse módulos de biorremediación y obtención de biogás, como los módulos de pretratamiento de efluentes residuales y el de digestión anaerobia de residuos con elevada carga orgánica; un módulo por etapas de cul tivos monoalgales o en consorcios microalga-bacteria; un módulo de recuperación de biomasa mediante métodos mecánicos, así como asis tidos por procesos fisicoquímicos, como la inducción de floculación; y el módulo de obtención de productos de interés, en donde se llevan a cabo procesos de extracción o reacción y purificación.
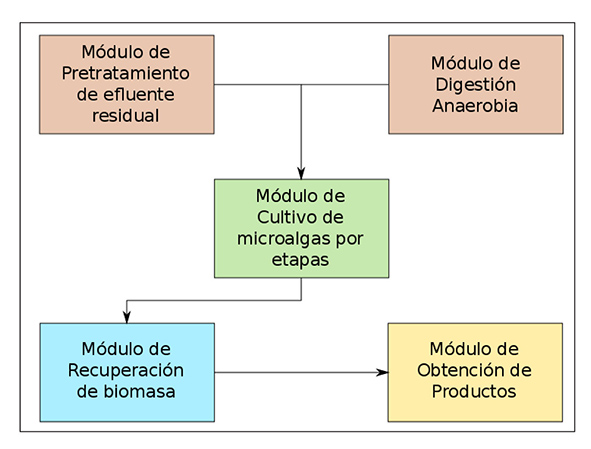
Figura 5 Diagrama modular de operación de sistemas de doble propósito para el cultivo de microalgas.
En lo que respecta al módulo de cultivo de microalgas por etapas, en la Figura 6 se muestra un ejemplo de las etapas en el cultivo de microalgas. La primera etapa consiste en la preservación de cepas, la producción y propagación del inóculo en fotobiorreactores. La producción del inóculo se realiza bajo condiciones controladas de crecimiento en medios de cultivo minerales sintéticos y estériles, lo que significa bajos volúmenes de producción. Por su parte, la propagación del inóculo se puede realizar en condiciones asépticas o en medios no estériles y con medios sintéticos o alternativos, como diluciones de efluentes residuales con agua corriente. Sin embargo, esto último lleva consigo la posible contaminación del inóculo y la competencia por nutrientes con otros microorganismos presentes en los medios alternativos no estériles (Olguín et al., 2013).
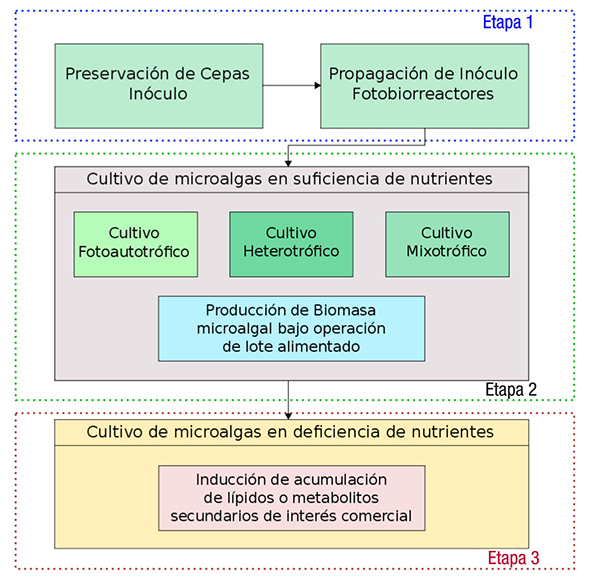
La segunda y tercera etapa del módulo se enfocan en la producción a mayor escala de las cepas o consorcios de microalgas de interés. El objetivo de la segunda etapa es obtener cultivos con alta densidad celular, por lo regular superiores a 1 g/L. En esta etapa se implementan cultivos fotoautotróficos, heterotróficos y mixotróficos, que se distin guen de acuerdo a la adición de su fuente de carbono y de su modo de operación (Lowrey et al., 2015).
En los cultivos fotoautotróficos la fuente de carbono es inorgánica, regularmente CO2, proporcionada mediante una mezcla aire-CO2, en donde la concentración de CO2 regularmente no rebasa el 10% (v/v) y la energía lumínica para las funciones fotosintéticas de las microalgas es suplementada por luz natural o artificial. En el caso de luz natural el fotoperiodo estará definido por la posición geográfica y estación del año, mientras que en caso de luz artificial los cultivos se pueden operar inclusive bajo luz continua.
En cultivos heterotróficos la fuente de carbono es orgánica, como glucosa, glicerol, algún ácido graso de bajo peso molecular (AGV) o en algunos casos se han utilizado residuos industriales ricos en glucosa como la melaza (Bumbak et al., 2011, Liu et al., 2013). Estos cultivos se realizan en fermentadores de gran capacidad bajo condiciones contro ladas de pH, temperatura, agitación y sin fuente de energía luminosa. La operatividad de estos procesos resulta costosa; sin embargo, las mayores densidades celulares en el cultivo de microalgas se alcanzan en este tipo de cultivos y en condiciones asépticas, por lo que son am pliamente utilizados para la obtención de nutracéuticos y productos de consumo humano (Bumbak et al., 2011).
En los cultivos mixotróficos se tienen ambas fuentes de carbono, orgánica e inorgánica, además de una fuente de luz, de manera que la microalga puede aprovechar ambas fuentes de carbono para su creci miento (Lowrey et al., 2015). En sistemas de doble propósito la fuente orgánica de estos cultivos regularmente proviene de carbono de efluen tes con elevada carga orgánica (digestato) y el carbono inorgánico de mezcla de gases de combustión y aire (Olguín et al., 2015a; 2015b). En algunos casos también se ha optado por sales de carbonato como fuente inorgánica de carbono (Murray et al., 2012; Olguín et al., 2015b).
Ya que en la segunda etapa del módulo de cultivo de microalgas el principal objetivo es maximizar la producción de biomasa, en la Figura 6 se sugiere que los cultivos sean operados como lote alimentado, de esta manera se ha observado que se obtienen altas densidades celu lares incluso en cultivos en continuo (Cheirsilp & Torpee, 2012; Ji et al., 2015). Bajo operación de lote alimentado los nutrientes que son rápidamente consumidos son repuestos periódicamente, de manera que la densidad no disminuya por la falta de nutrientes. En cultivos mixotróficos, donde los nutrientes provienen de efluentes residuales, la operación por lote alimentado puede no ser la más apropiada, ya que agregar periódicamente efluente residual al cultivo incrementa la turbidez y limita el paso de luz inhibiendo los procesos fotosintéticos de las microalgas. Ante esto, para la operación de lote alimentado en estos cultivos se puede optar por agregar sales minerales de bajo costo únicamente de los macronutrientes más importantes, como NH4HCO3, que cumple la doble función de ser fuente tanto de nitrógeno como de carbono (Olguín et al., 2015b).
El objetivo de la tercera etapa es la inducción de acumulación de productos de interés. Estos pueden ser lípidos o metabolitos secunda rios, como pigmentos o carotenoides, que ante condiciones de estrés la microalga utiliza como mecanismos de defensa (Hu et al., 2008; Le moine & Schoefs, 2010; Mulders et al., 2014). En el caso de los lípidos, se considera que ante estrés nutricional o fotoinhibitorio, las microalgas acumulan estos en organelos que sirven como depósitos de reserva de energía (Wijffels & Barbosa, 2010; Klok et al., 2013). La falta o limita ción de nitrógeno en los medios de cultivo de microalgas se considera como el principal estrés nutricional involucrado en la acumulación de lípidos y de metabolitos secundarios (Gouveia & Oliveira, 2009).
La etapa de estrés puede realizarse en los mismos fotobiorreac tores de la segunda etapa sin la adición de nutrientes y alargando el tiempo de residencia, hasta obtener la máxima acumulación de meta bolitos de interés sin que se presente una disminución significativa de su densidad celular. Durante la operación de esta etapa no se aconseja la dilución del medio de cultivo para disminuir la concentración de nu trientes, ya que esto a su vez diluye la densidad celular y dificulta aún más la recuperación de biomasa.
Con un enfoque sustentable de sistemas de doble propósito, la Figura 7 muestra el diagrama de bloques para obtención de productos de alto valor agregado y biocombustibles a partir del cultivo de microalgas en dos procesos interconectados en el concepto de una biorrefinería. En el Proceso I, para la obtención de pigmentos y nutracéuticos, el cul tivo supone condiciones de crecimiento controladas y medios de cultivo estériles. Mientras que en el Proceso II, el cultivo de las microalgas o consorcio microalgas-bacterias se realiza en condiciones exteriores a grandes volúmenes y bajos costos de operación.
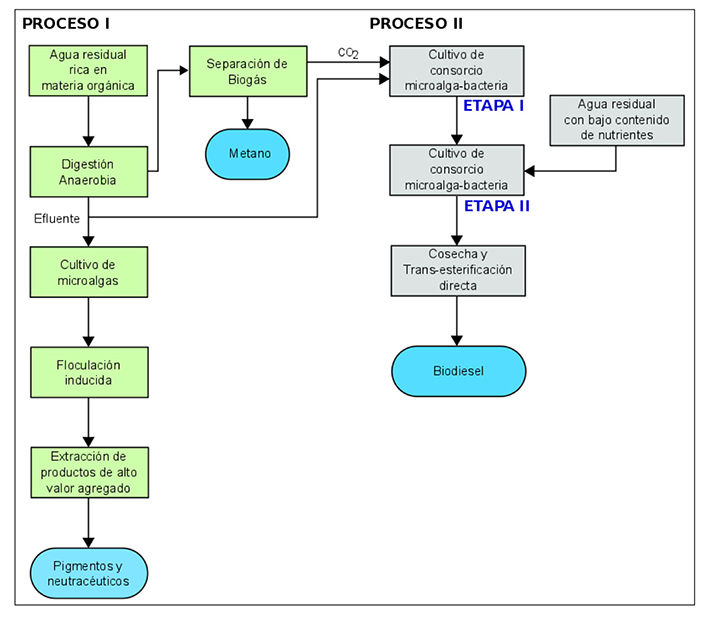
Figura 7 Diagrama de bloques para el cultivo de microalgas en un sistema de doble propósito aplicado al concepto de biorrefinería
Aun cuando se tiene toda esta información sobre la producción de biodiésel y la obtención de compuestos de alto valor agregado a partir de la biomasa de microalgas, existe mucho campo fértil para realizar investigación sobre el mejoramiento de métodos de cultivo, la identificación y obtención de nuevos productos, mejoramiento de procesos de cosecha y de extracción de compuestos, análisis de ciclo de vida y estudios de viabilidad económica, entre muchos otros (Freire et al., 2011).
Conclusiones
La necesidad de desarrollar combustibles renovables y ecológicos es cada vez más evidente. El uso de microalgas para la obtención de biodiésel representa una alternativa viable por su alto contenido lipídico y su perfil de ácidos grasos, sin embargo, hace falta el desarrollo de tecnologías que reduzcan el costo de producción. La transesterificación en la actualidad resulta ser el método más barato para la producción de biodiésel. Sin embargo, actualmente no se han logrado precios competitivos frente a los producidos a partir de recursos fósiles. Los principales factores que incrementan los precios de obtención de biodiésel son los métodos de cultivo, la extracción de lípidos (downstream) y los procesos de bioconversión.
El uso de campos eléctricos, extracción por presurización cíclica, microondas y sonicación se perfilan como métodos novedosos para la recuperación de lípidos intracelulares. Actualmente, el proceso de transesterificación puede realizarse mediante el uso de lipasas comer ciales o mediante la catálisis directa, utilizando microorganismos como levaduras y hongos filamentosos. Los sistemas de cultivo de doble propósito son un proceso novedoso que podría disminuir los costos, debido a que permite incrementar la biomasa reincorporando los nutrientes presentes en efluentes residuales. En este trabajo, además, se propone un diseño modular para sistemas de doble propósito y se describen detalladamente diferentes etapas de este diseño, principalmente enfocadas en el módulo de cultivo de microalgas que es la parte nodal de esta investigación.

