











 nova página do texto(beta)
nova página do texto(beta) Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO 
 Permalink
PermalinkINTRODUCCIÓN
Las lagunas costeras son importantes ecosistemas marinos por su alta producción primaria y relevancia ecológica. Algunos procesos que re gulan y definen la productividad de estos sistemas son el ciclo y flujo de nutrientes, la descomposición y transformación de materia orgánica y la remoción de contaminantes (Brito et al., 2012). El conocimiento de los balances de nutrientes (entradas y salidas) y otras variables hidroló gicas como oxígeno disuelto, clorofila a, entre otras, en los cuerpos de agua lagunares, permiten hacer inferencias acerca de su biota y su es tado trófico. Existen diferentes aproximaciones para estimar los flujos de material disuelto y particulado en los sistemas marinos costeros (De la Lanza-Espino & Rodríguez-Mena 1993; Gordon et al., 1996; Alcánta ra et al., 2012). Las lagunas costeras del oeste de la península de Baja California (México) presentan una variedad de ambientes geomorfoló gicos (Lankford 1977) y procesos oceanográficos, como la actividad de surgencias, que contribuyen a realizar un estudio comparativo (Zaytsev et al., 2003). Debido a su ubicación, su importancia para la economía regional y su estatus de sitio Ramsar, Bahía Magdalena (BM) ha sido objeto de varias investigaciones ecológicas (Funes-Rodríguez et al., 2007) e interdisciplinarias (Bizarro 2008). La calidad del agua de la laguna ha sido estudiada desde varios puntos de vista, incluyendo la dinámica de nutrientes (Álvarez-Borrego et al., 1975, Cervantes-Duarte et al., 2007, 2010, 2013). En este sistema antiestuarino se han rea lizado modelaciones numéricas y trabajo de campo para determinar el patrón de circulación e intercambio de agua entre la laguna y el mar adyacente (Morales-Zárate et al., 2006; Sánchez-Montante et al., 2007, Zaitsev et al., 2007). Estos estudios mostraron que las aguas subsuperficiales oceánicas, relativamente frías y ricas en nutrientes, se presentan en la vecindad inmediata de la boca de BM y son suscep tibles de ser transportadas hacia el interior del sistema por la acción de mareas (Cervantes-Duarte et al., 2012). En consecuencia, las con centraciones de nutrientes en la laguna dependen en gran medida de la calidad de las aguas costeras adyacentes. Hasta ahora los estudios sobre la importancia del intercambio de agua entre la laguna y el mar (Acosta-Ruíz & Lara-Lara, 1978; Guerrero et al., 1988; Robinson et al., 2007) no han estimado el transporte de nutrientes y clorofila a como una medida del proceso responsable de la productividad. De acuerdo con los antecedentes oceanográficos en esta región, suponemos que los nutrientes aportados por el océano durante los procesos de sur gencia y transportados por las corrientes de marea, fertilizan la laguna aumentando la biomasa fitoplanctónica. Con el propósito de analizar el efecto de la marea sobre la distribución de variables hidrológicas y calcular los flujos y el transporte de nutrientes y clorofila a en Bahía Magdalena y el océano adyacente, se realizaron 3 campañas de 24 horas cada una, en un sitio cercano a la boca principal en junio, agosto y noviembre del 2005.
MATERIALES Y MÉTODOS
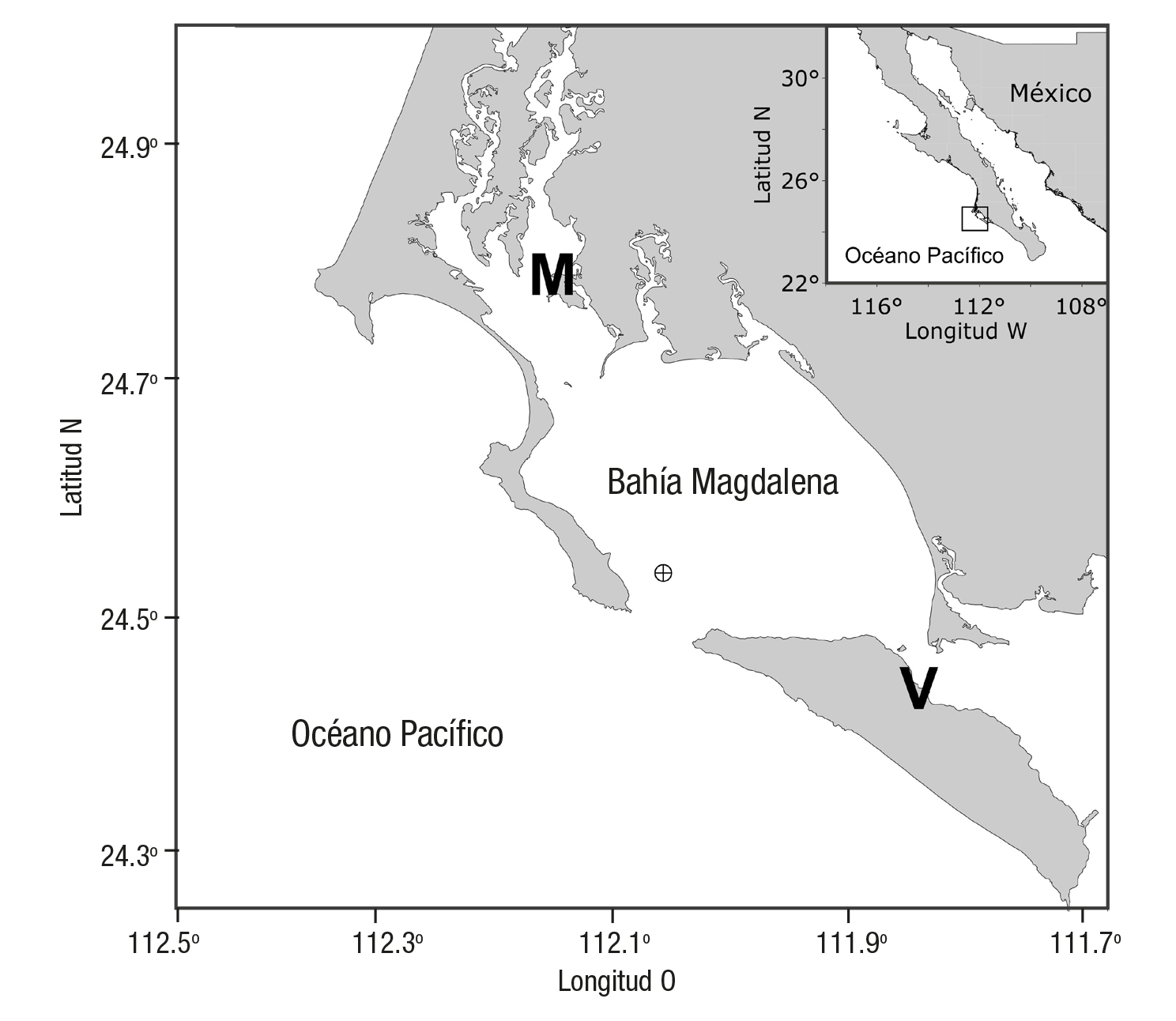
Bahía Magdalena está localizada en el suroeste de la península de Baja California (24° 30’ - 24° 48’ N y 111° 51’ - 112° 09’ O), es el cuerpo de agua costero más extenso y más profundo de la región y forma parte de un complejo sistema lagunar (Funes-Rodríguez et al., 2007). Tiene un área de aproximadamente 565 km2 con un volumen total de agua (marea baja-alta) de 6.8 a 8.1 km3. Las mareas son mixtas semidiurnas la mayor parte del año y su rango varía de 0.7 a 2.3 m (micromareal) (Hayes 1975). La zona interna es (< 10 m) y ocupa alrededor de la mitad de la superficie de la laguna, mientras que la zona más profunda se comunica con el océano a través de una boca de 5.6 km de ancho y 38 m de profundidad (Fig. 1). Durante el año 2005 se realizaron 3 campañas en diferentes épocas del año (2 días de duración): 29-30 de junio (principios de verano boreal), 17-18 de agosto (verano) y 14-15 de noviembre (otoño), a bordo de la embarcación CICIMAR XX en un sitio ubicado a 3 km al noroeste de la parte central de la boca de Bahía Magdalena (24º 57´N y 112º 03´O) (Fig. 1). El propósito de estos muestreos fue determinar las características del agua, del océano y la laguna que se intercambian por ciclo de marea. Cada muestreo se efectuó cada 2 h y durante un lapso de 24 h. Se realizaron lances verti cales de CTD Seabird 19Plus para medir la temperatura (oC) y salinidad (adimensional) en la columna de agua. Cabe destacar que además se tomaron muestras discretas de agua para calcular el oxígeno disuelto (OD), nutrientes (nitrato, nitrito, amonio y fosfato) y clorofila a, a pro fundidades de 0, 15 y 30 m con una botella Niskin, General Oceanic de 5 L. Las muestras para OD fueron colectadas y fijadas en botellas de vidrio de 125 ml, y transportadas en una caja obscura hasta su análisis al día siguiente del muestreo en el laboratorio. Las submuestras para la determinación de sales nutrientes y clorofila a (aproximadamente 1.5 L) fueron tomadas de la botella Niskin y refrigeradas a 4 oC hasta ser filtradas en el laboratorio. Cerca de 12 horas después de su colecta, las muestras para clorofila a (1 L) fueron procesadas con filtros What man GFF de 25 mm de diámetro. El contenido del filtro fue utilizado para extraer la clorofila a de acuerdo con el procedimiento de Venrick y Hayward (1984) y luego cuantificar la concentración con al méto do de Jeffrey & Humphrey (1975) con ayuda de un espectrofotómetro (Spectronic Genesys-2). El filtrado de cada muestra (aproximadamente 100 ml por analito) fue almacenado en un congelador a -50 oC para los análisis de amonio, nitrato, nitrito y fosfato, con base en los métodos de Strickland y Parsons (1972). En este estudio se consideró el nitrógeno inorgánico disuelto (NID) como la suma de nitrito, nitrato y amonio. Las unidades e intervalo de detección para cada variable fueron los siguien tes: clorofila a (0.1 - 10 mg m-3); nitrito (0.01 - 2.5 µM); nitrato (0.05- 25 µM); amonio (0.04 - 5 µM); fosfato (0.03 - 2.5 µM) y oxígeno disuelto (0.07 - 7.0 ml l-1). Los datos de la amplitud de la marea (pronósticos respecto al NMM) y viento (dirección en grados y velocidad en metros por segundo) fueron obtenidos de las páginas: http://predmar.cicese.mx/programa/ y http://meteorologia.semar.gob.mx/, respectivamente. La ubicación de las estaciones del mareógrafo y anemómetro se indi can en la Figura 1. Para determinar si existían diferencias significativas entre las variables medidas en las tres profundidades seleccionadas (0, 15 y 30 m) durante el ciclo de 24 horas, y considerando los supuestos de normalidad (prueba de Levene), se realizó un análisis de varianza de una vía (ANOVA) y una prueba a posteriori de comparaciones múltiples por pares de Tukey HSD (Honestly-Significant-Difference), con un nivel de confianza de 95%. Asimismo, se llevaron a cabo pruebas de aso ciación y relación entre la concentración de clorofila a como indicador de la biomasa fitoplanctónica y el resto de las variables medidas. El transporte de volumen de agua transversal a la boca (gasto) integrado en la vertical y a lo largo de la sección longitudinal (172 x 103 m3 s-1 en flujo y en 162 x 103 m3 s-1 en reflujo), fue estimado a partir de medicio nes in situ y modeladas con el modelo numérico de circulación costera (ECOM, por sus siglas en inglés) (Sánchez-Montante et al., 2007).
RESULTADOS
El 29 y 30 junio de 2005 la marea tuvo una amplitud de 1.01 m y el viento dominante fue del noreste (NE) (73%), con una velocidad prome dio de 2.2 ± 0.8 m s-1. La temperatura en la columna de agua (0, 15 y 30 m) presentó los valores más altos en superficie (18.7 ± 1.5 oC) y los menores en el fondo (17.6 ± 1.5 oC), con una diferencia de hasta 5 oC entre ambas profundidades. El valor máximo se registró a las 19 h (22.4 oC a 0 m) y el mínimo a las 07 h (15.8 oC a 30 m) (Fig. 2a). Los datos no cumplieron el criterio de normalidad (Tabla 1) y la relación con la altura de la marea fue significativa (Tabla 2). La salinidad superficial (34.0 ± 0.2) fue significativamente menor que a 15 y 30 m (34.3 ± 0.0) (Tabla 1), excepto al inicio del muestreo de las 17 a las 21 h (Fig. 2b). La relación con la altura de marea no fue significativa. La densidad de la columna de agua varió entre un mínimo de 24.3 kg m-3 en superficie a un máximo de 25.4 kg m-3 a 30 m de profundidad (Fig. 2c). Se regis traron diferencias significativas entre las profundidades (0, 15 y 30 m) y no se observó relación significativa con la amplitud de la marea (Tabla 2). La saturación de oxígeno (%) durante el muestreo obtuvo valores por arriba de la saturación en la superficie (84-124%) y disminuyeron significativamente a los 15 m (31-72%) y 30 m (24-47%) (Tabla 1 y Fig. 2d). Sólo a una profundidad de 30 m se relacionó significativamente con la amplitud de la marea (Tabla 2). Durante el ciclo de muestreo, los nutrientes (NID y fosfato) mostraron el mismo patrón de distribución y su relación fue alta y significativa (r = 0.90; p < 0.001); ambos incre mentaron su concentración de la superficie al fondo (Tabla 1; Fig. 2e) y no se relacionaron de forma significativa con la altura de marea (Tabla 2). La clorofila a disminuyó significativamente de la superficie (3.7 ± 1.2 mg m-3) al fondo (2.3 ± 0.8 a 15 m y 1.4 ± 0.2 mg m-3 a 30 m) (Tabla 1 y Fig. 2f). La relación con la amplitud de la marea no fue significativa en ninguna de las profundidades (Tabla 2).
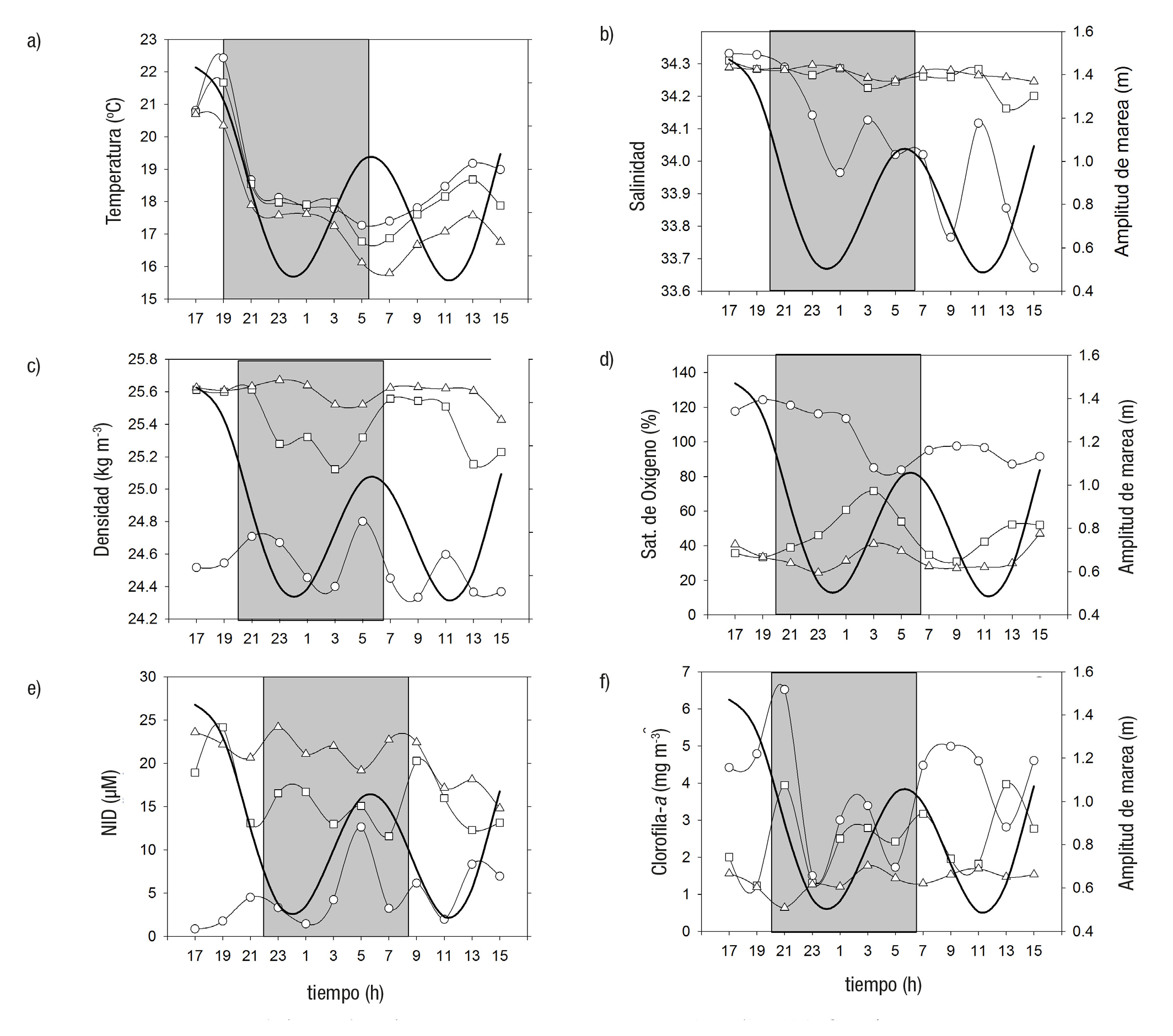
Figuras 2a-f Distribución de las variables medidas en la boca de Bahía Magdalena, México, en junio, a 0 (O), 15 (□) y 30 m (Δ). a) temperatura, b) salinidad, c) den sidad, d) oxígeno, e) nutrientes, f) clorofila (Cl-a).
Tabla 1 Valores promedio y desviación estándar por ciclo de 24 horas en la boca de Bahía Magdalena, B.C.S., México (en negritas se indican dife rencias significativas entre profundidades (p < 0.05).
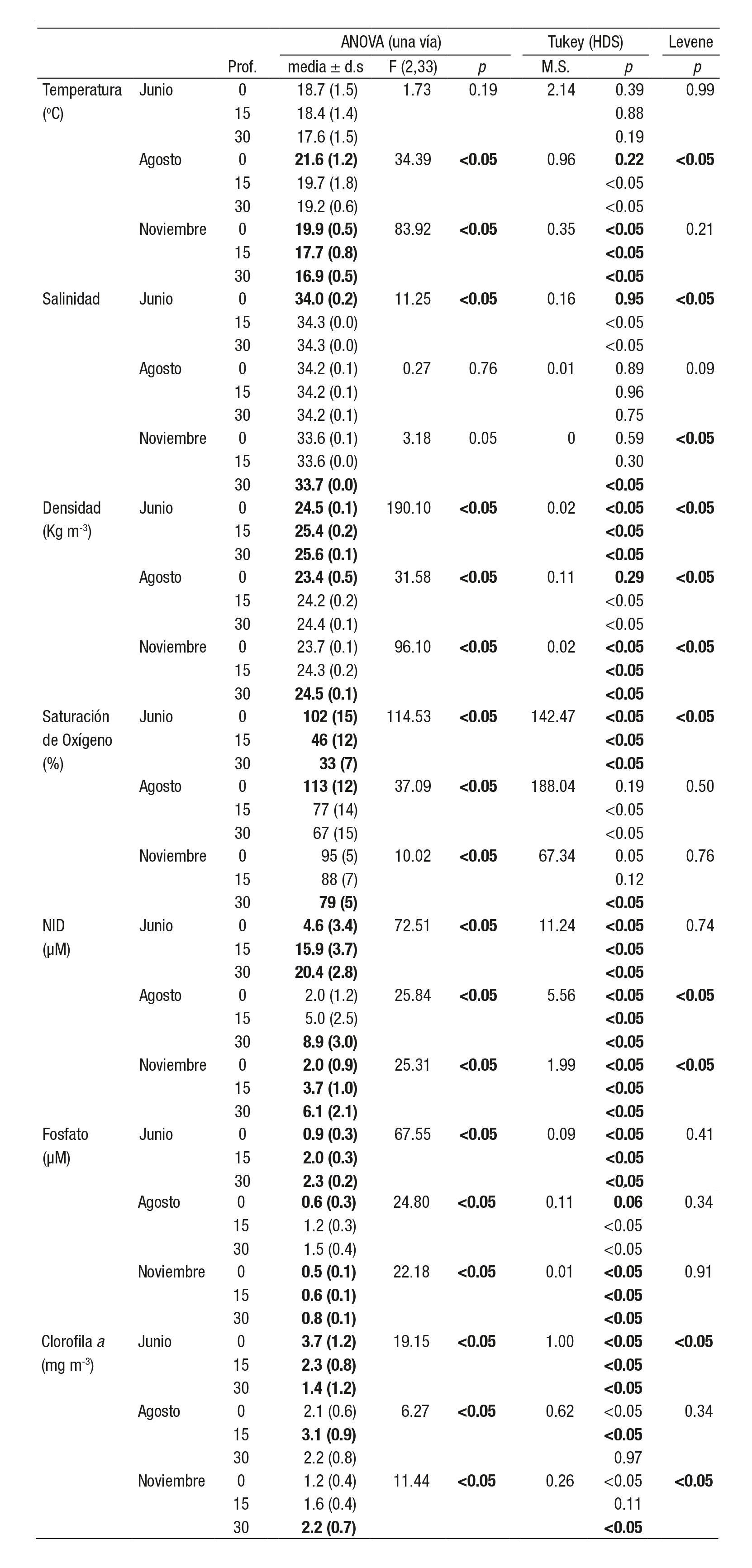
Tabla 2 Coeficiente de correlación de Pearson (r) y valor de significancia p de la amplitud de la marea y la variable medida durante el ciclo de 24 h (boca de bahía Magdalena, B. C. S., México, 2005). En negritas se indican las pruebas significativas.
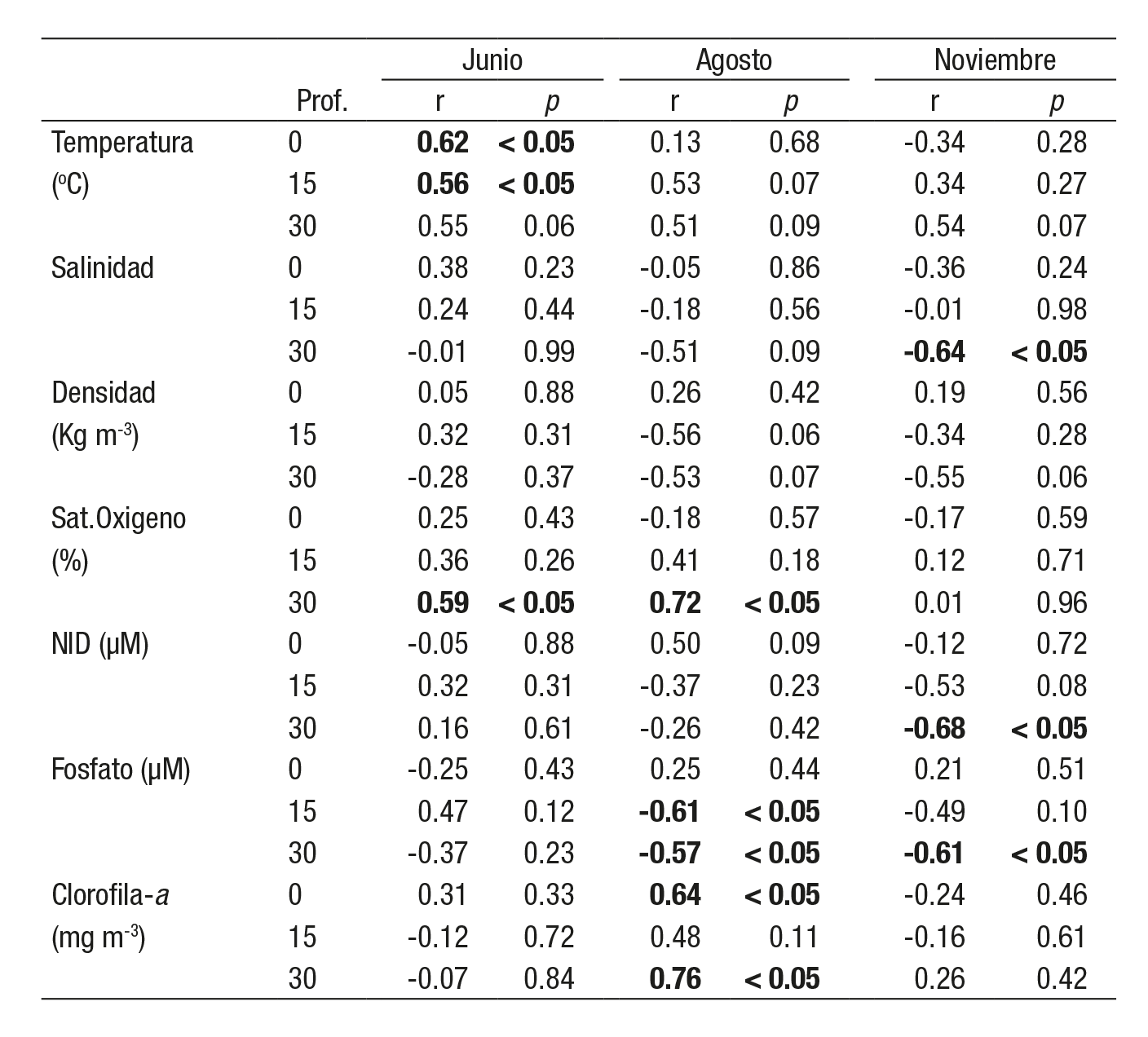
El 17 y 18 de agosto la marea tuvo una amplitud de 2.3 m y el viento dominante fue del suroeste (SO) (55%), con una velocidad de 3.1 ± 0.4 m s-1. La temperatura en la columna de agua varió durante el ciclo de un máximo superficial de 24.3 oC a las 14 h, a un mínimo de 18.3 oC a 30 m de profundidad a las 24 h (Tabla 1, Fig. 3a). La temperatura superficial fue significativamente mayor que a los 15 y 30 m de profundidad. Las relaciones entre la temperatura y la amplitud de la marea no fueron sig nificativas (Tabla 2). La salinidad, en las tres profundidades, desarrolló un patrón similar de variación, las mayores diferencias se observaron en la superficie, con un mínimo de 34.0 a las 06:00 h y un máximo de 34.4 a las 18:00 h (Fig. 3b). La relación con la altura de la marea fue negativa y no significativa (Tabla 2). Durante el ciclo, la densidad aumentó de la superficie al fondo, con un mínimo de 22.4 kg m-3 a las 14 h, a un máxi mo de 24.7 kg m-3, a las 24:00 h (Fig. 3c). La densidad superficial fue significativamente menor a 15 y 30 m y la correlación con la altura de la marea en ninguno de los casos fue significativa. La saturación de oxige no (%) fue alta en la superficie (94-129%) y mínima a 30 m (48-89%). Únicamente a 30 m se registró una relación significativa con la amplitud de la marea (Tabla 2). El NID y fosfato manifestaron el mismo patrón de variación en el ciclo con un incremento de la superficie al fondo (Fig. 3d; Tabla 1). La relación entre ambos nutrientes fue alta y significativa (r = 0.84; p < 0.001). El NID presentó diferencias significativas entre las profundidades (0, 15 y 30 m) y la relación con la altura de la marea no fue significativa. La clorofila a en la columna de agua obtuvo un máximo a 15 m (3.1 ± 0.9 mg M-3) respecto a la superficie (2.1 ± 0.6 mg M-3) y a 30 m (2.2 ± 0.8 mg m-3). La relación con la amplitud de la marea fue significativa a 0 y 30 m (Tabla 2).
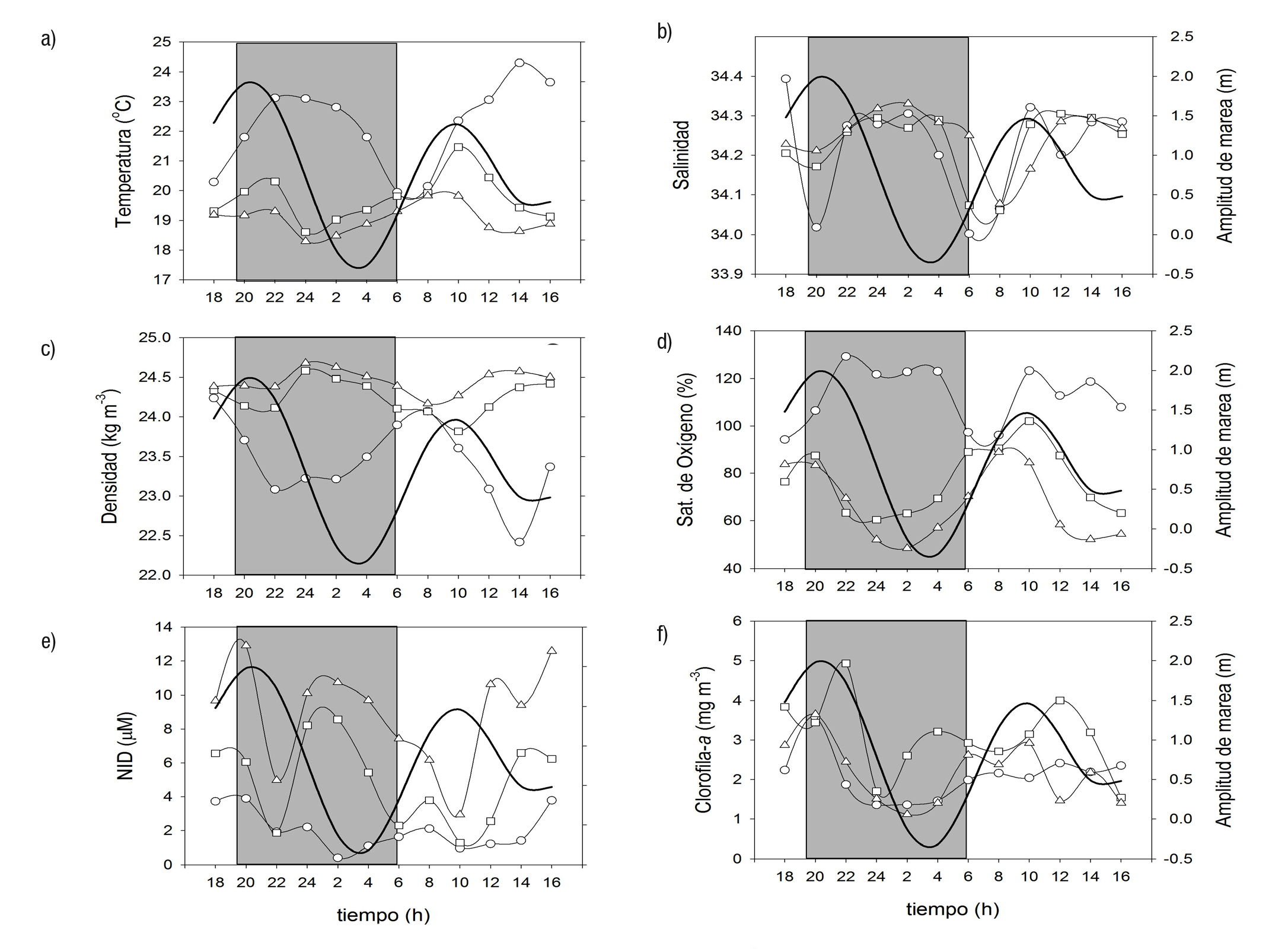
Figuras 3a-f Distribución de las variables medidas en la boca de Bahía Magdalena, México en agosto, a 0 (O), 15 (□) y 30 m (Δ). a) temperatura, b) salinidad, c) densidad, d) oxígeno, e) nutrientes, f) clorofila (Cl-a).
El 14 y 15 de noviembre el viento dominante fue del sur-sureste (SSE) (78%), con una velocidad promedio de 2.3 ± 1.0 m s-1 y una amplitud de marea de 2.4 m. La temperatura en la columna de agua tuvo el valor más alto en la superficie (20.7 oC) a las 16 h y el mínimo (16.3 oC) a las 18 h (Fig. 4a). En promedio, la temperatura disminuyó de la superficie (19.9 ± 0.5 oC) al fondo (17.7 ± 0.8 a 15 m y 16.9 ± 0.5 oC a 30 m). La relación con la amplitud de marea no fue significativa (Tabla 2). La salinidad manifestó una desviación estándar de 0.09 en la superficie en el ciclo y en promedio aumentó de 33.6 (en superficie) a 33.7 (a 30 m) (Tabla 1; Fig. 4b). La salinidad a 30 m fue significativa mente mayor que la calculada a 0 y 15 m (Tabla 1), y se relacionó con la amplitud de marea (Tabla 2). La densidad promedio se incrementó de la superficie al fondo (23.7 ± 0.1; 24.3 ± 0.2 y 24.5 ± 0.1 kg m-3) (Tabla 1; Fig. 4c). No se relacionó de forma significativa con la amplitud de la marea (Tabla 2). La saturación de oxígeno disminuyó de la superficie (95 ± 5 %) al fondo (88 ± 7 a 15 m y 79 ± 5 % a 30 m) (Tabla 1; Fig. 4d). No hubo un vínculo significativo con la amplitud de marea (Tabla 2). Los nutrientes (NID y fosfato) se acrecentaron de la superficie al fondo durante el muestreo (Tabla 1; Fig. 4e) y se relacionaron positiva y significativamente (r = 0.83; p < 0.001). Se pudieron observar en el NID diferencias significativas entre las tres profundidades (Tabla 1) y la concentración a 30 m se relacionó de manera significativa con la amplitud de marea, tanto para NID como fosfato (Tabla 2). La clorofila a aumentó de la superficie (1.2 ± 0.4 mg m-3) al fondo (1.6 ± 0.4 a 15 m y 2.2 ± 0.7 mg m-3 a 30 m) (Tabla 1; Fig. 4e). La concentración a 30 m fue mayor que a 0 y 15 m y no se relacionó de modo significativo con la amplitud de marea (Tabla 2).

DISCUSIÓN
Durante el año 2005 se identificaron en el sitio de estudio tres dife rentes condiciones climático-oceanográficas. La primera se registró en junio (principios de verano) con la influencia del viento dominante del NO cuando se observó agua más fría y con mayor clorofila a en la zona de la boca y exterior de la laguna (Fig. 5a y 5b) debido a las surgencias costeras, las cuales ocurren con mayor intensidad de marzo a junio (Zaytsev et al., 2003; Cervantes et al., 2012; Cervantes et al., 2013). La columna de agua se caracterizó por tener en promedio la menor temperatura (<18 oC) y mayor densidad (hasta 25.59 kg m-3 a 30 m de profundidad), así como los más altos nutrientes (hasta 20.4 µM de NID y 2.3 µM de fosfato a 30 m de profundidad) y clorofila a (3.7 mg m-3 en superficie) durante el periodo de estudio. La temperatura y la amplitud de la marea se relacionaron positiva y significativamente (r = 0.6; p < 0.05), lo que indica que durante este ciclo de marea, especialmente en su máxima amplitud, la salida de agua fue más cálida durante el reflujo (Fig. 2a). La biomasa de fitoplancton (medida por la clorofila a) reveló una relación positiva y significativa con la saturación de oxígeno (r = 0.73), posiblemente como resultado de la fotosíntesis del fitoplancton, con valores arriba del 100% de saturación cuando la clorofila a fue máxima y disminuyeron ambas variables con el incremento de la pro fundidad. En agosto (verano) ocurrió una estratificación en los primeros 10 metros de la columna de agua debido al aumento de temperatura. Esta época es la más cálida y oligotrófica del año (Fig. 5c y 5d). El viento dominante típicamente proviene del SSE con velocidad casi siempre menor (3.1 ± 0.4 m s-1 en este estudio) a la observada cuando la di rección es NO (promedio 5 y hasta 15 m s-1), excepto en temporada de tormentas tropicales (García-Escobar, 1992). En este mes, bajo las con diciones de una mayor amplitud de la marea, se registró el mayor nú mero de relaciones significativas (y de signo contrario) con los nutrien tes y positivo con la clorofila a. La temperatura presentó una relación inversa y significativa con los nutrientes (DIN = -0.74 y fosfato = -0.87), es decir, que el agua superficial más cálida fue pobre en nutrientes, resultado de su utilización y por la estratificación térmica que evita el transporte desde el fondo. La relación de temperatura con la saturación de oxígeno fue positiva (r = 0.90), posiblemente debido a la producción de oxígeno del fitoplancton, el cual tuvo un máximo de clorofila a a 15 m de profundidad. La imagen de clorofila a (Fig. 5d) mostró una dis tribución homogénea en la región con valores alrededor de 2 mg m-3, consistentes con los registrados in situ (2.1 ± 0.6 mg m-3). La relación de la clorofila a con el resto de las variables fue baja y no significativa, excepto con la amplitud de la marea (r = 0.51) de manera similar a lo reportado por Acosta-Ruíz y Lara-Lara (1978), quienes sugieren que Bahía Magdalena aporta altas biomasas de fitoplancton (clorofila a) al océano abierto durante el reflujo. La tercera condición tuvo lugar en noviembre (otoño) donde la salinidad fue la menor del estudio (< 34), y que junto con la temperatura (< 21 oC) (Fig. 5e) corresponde con la del agua de la Corriente de California (Durazo & Baumgartner, 2002; Durazo, 2015), que generalmente es más rápida en invierno-primavera (0.15 a 0.20 m s-1 en primavera), pero que debido a un enfriamiento regional pudo haber favorecido su flujo al sur en esta época del año, como lo indica el cambio de signo del índice multivariado El Niño Osci lación del Sur (ENSO por sus siglas en inglés) con un incremento de la corriente al sur (http://www.esrl.noaa.gov/psd/enso/mei) (Fig. 6). Las relaciones con la amplitud de la marea generalmente fueron negativas (Tabla 2), lo cual evidencia el contraste entre las condiciones ambienta les de flujo y reflujo en la laguna durante un ciclo de marea. Excepto la amplitud de la marea, el resto de las variables presentaron relaciones significativas entre ellas. La temperatura se relacionó inversa y signi ficativamente con la densidad (r = -0.99) y los nutrientes (r = -0.79 y -0.74 para NID y fosfato respectivamente), lo que indica que aguas más frías son más densas y con mayor concentración de nutrientes y viceversa. La clorofila a se relacionó positivamente con los nutrientes (r = 0.47 y 0.40 para NID y fosfato respectivamente), esto es, la corre lación entre la mayor disponibilidad con la mayor biomasa fitoplanc tónica (Barber & Hiscock, 2006). En este ciclo la clorofila a promedio a 30 m fue significativamente mayor que a 0 y 15 m, es decir, que la distribución de clorofila no es homogénea en la columna de agua; su distribución espacial mostró mayor concentración dentro de la laguna posiblemente como resultado del aporte de nutrientes remineralizados (Fig. 5f) (Cervantes-Duarte et al., 2012).
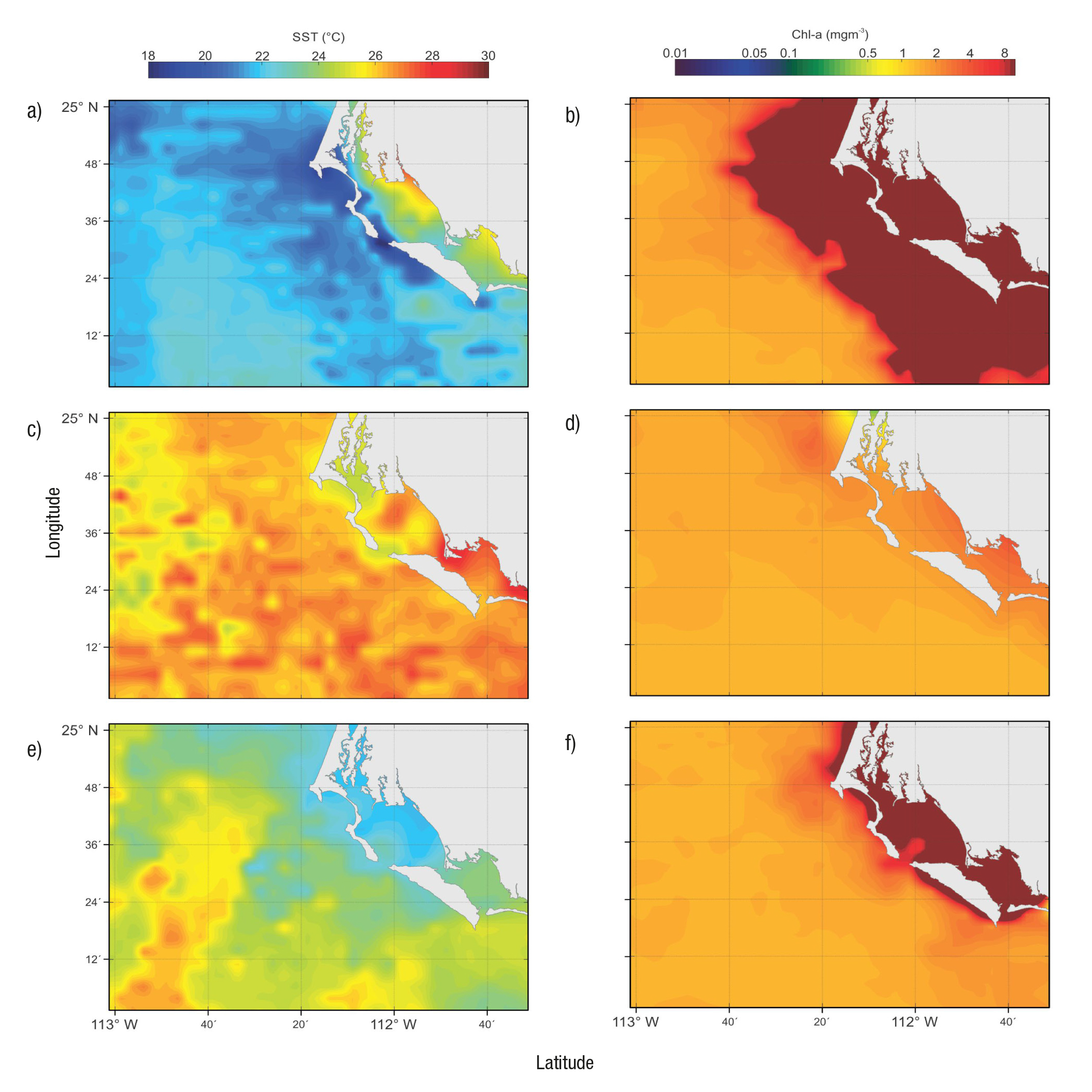
Figuras 5a-f Imágenes de temperatura (oC) y clorofila a (mg M-3) en la región de Bahía Magdalena, México, composiciones promedio de ocho días (incluidos los días de muestreo), resolución de 4 km, plataforma MODIS-Aqua. a-b) junio, c-d) agosto, e-f) noviembre de 2005.
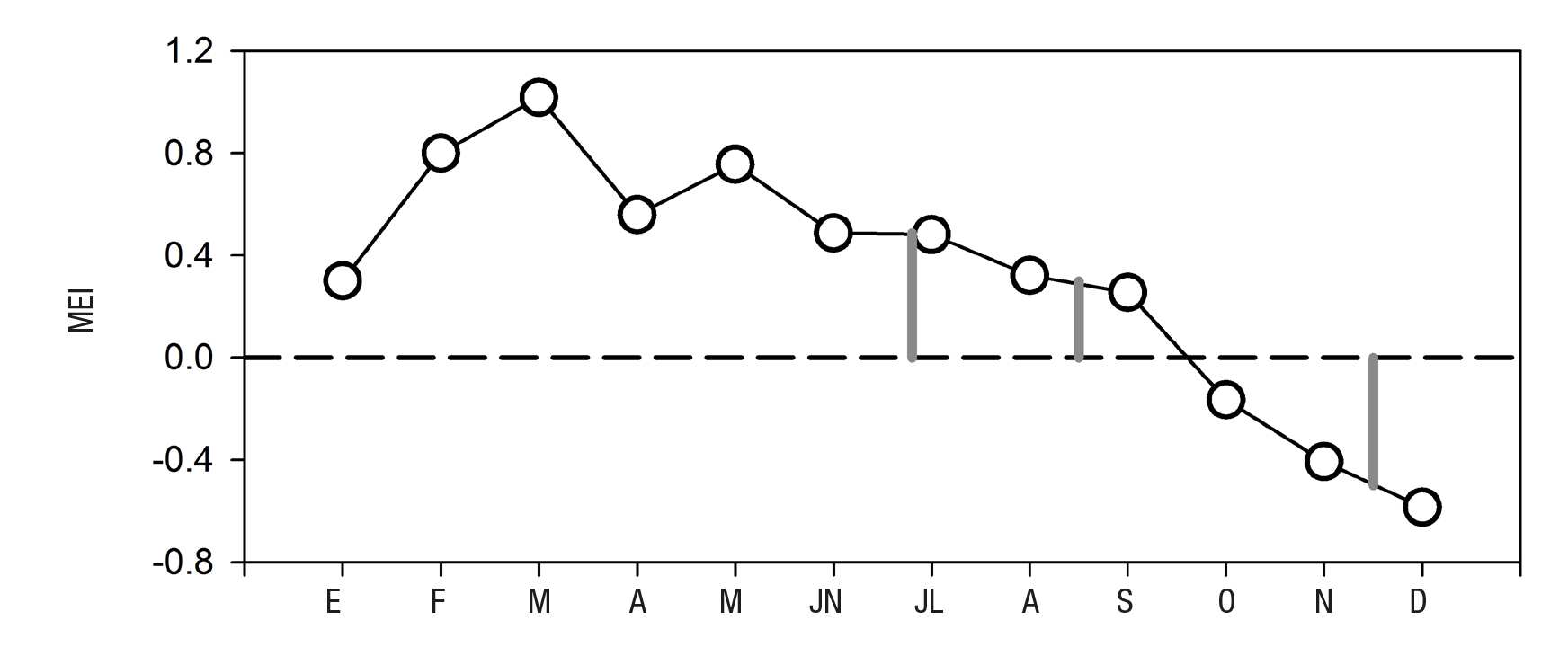
Figura 6 Variación del índice mutivariado El Niño (MEI) en el Pacífico oriental, durante los distintos meses del año 2005.
En promedio, la saturación de oxigeno disminuyó de ≈100 % en la superficie al fondo (33% en junio, 67% en agosto y 79% en noviembre). Los valores mínimos registrados en junio es posible que se deban al origen subsuperficial del agua (aproximadamente 80 m) transportada hacia la superficie por el proceso de surgencia costera (Cervantes-Duarte et al., 2013; Zaitsev et al., 2014). En verano y otoño (agosto y noviembre) los valores de oxígeno debajo de la superficie tienden a disminuir de manera normal por el aumento de temperatura en el año y el aumento en la tasa de respiración de los organismos.
Los nutrientes se incrementaron en general de la superficie al fon do y su concentración fue mayor en junio, esto asociado al abasto por surgencias. El proceso antes mencionado funciona como un sistema de fertilización que incrementa la biomasa de clorofila a en la región. En agosto y noviembre una menor disponibilidad de nutrientes produjo una disminución relativa de la clorofila a en la superficie, con máximos significativos a 15 y 30 m en agosto y noviembre, respectivamente, y de forma regular están relacionados con los procesos de estratificación en la columna de agua (Murillo-Murillo et al., 2013).
Para obtener una primera aproximación del transporte neto de nu trientes a través de la boca en mareas vivas, se utilizó el volumen máxi mo esperado en flujo y en reflujo (Sánchez-Montante et al., 2007) y el sentido de la marea fue determinado por la amplitud de la misma como se observa en el gráfico de la anomalía de densidad a la profundidad de 30 m (Fig. 7). El transporte neto de nutrientes por ciclo de marea para junio (29-30) fue de -143 kg de NID y -4 kg de fosfato, que indican una salida de nutrientes desde la laguna durante el reflujo, los cuales es posible que no fueron asimilados en su totalidad en la laguna debido a su alta disponibilidad. En agosto (17-18) y noviembre (14-15) los trans portes fueron respectivamente de 209 y 176 kg de NID y 30 y 3 kg de fosfato. En éstos últimos, entran nutrientes hacia la laguna durante el flujo de la marea. En términos de nitrógeno, los valores son superiores a los reportados en el estero Agua Dulce Sinaloa, México, de 26.4 kg h-1 amonio y 18.6 kg h-1 nitrato durante un ciclo de marea de 6 h (De la Lanza-Espino & Rodríguez-Mena, 1993), y en la Bocana de Ancao, Portugal, de 3.9 kg de nitrato + nitrito por ciclo de marea. Sin embargo, el transporte de fosfato fue más variable en este estudio que el valor de 16 kg de fosfato reportado por estos autores por ciclo de marea (Alcán tara et al., 2012); en ambos casos determinados en bocas pequeñas y someras con transporte de agua mucho menor, comparado con la boca de Bahía Magdalena que tiene 5.6 km ancho y 40 m de profundidad.
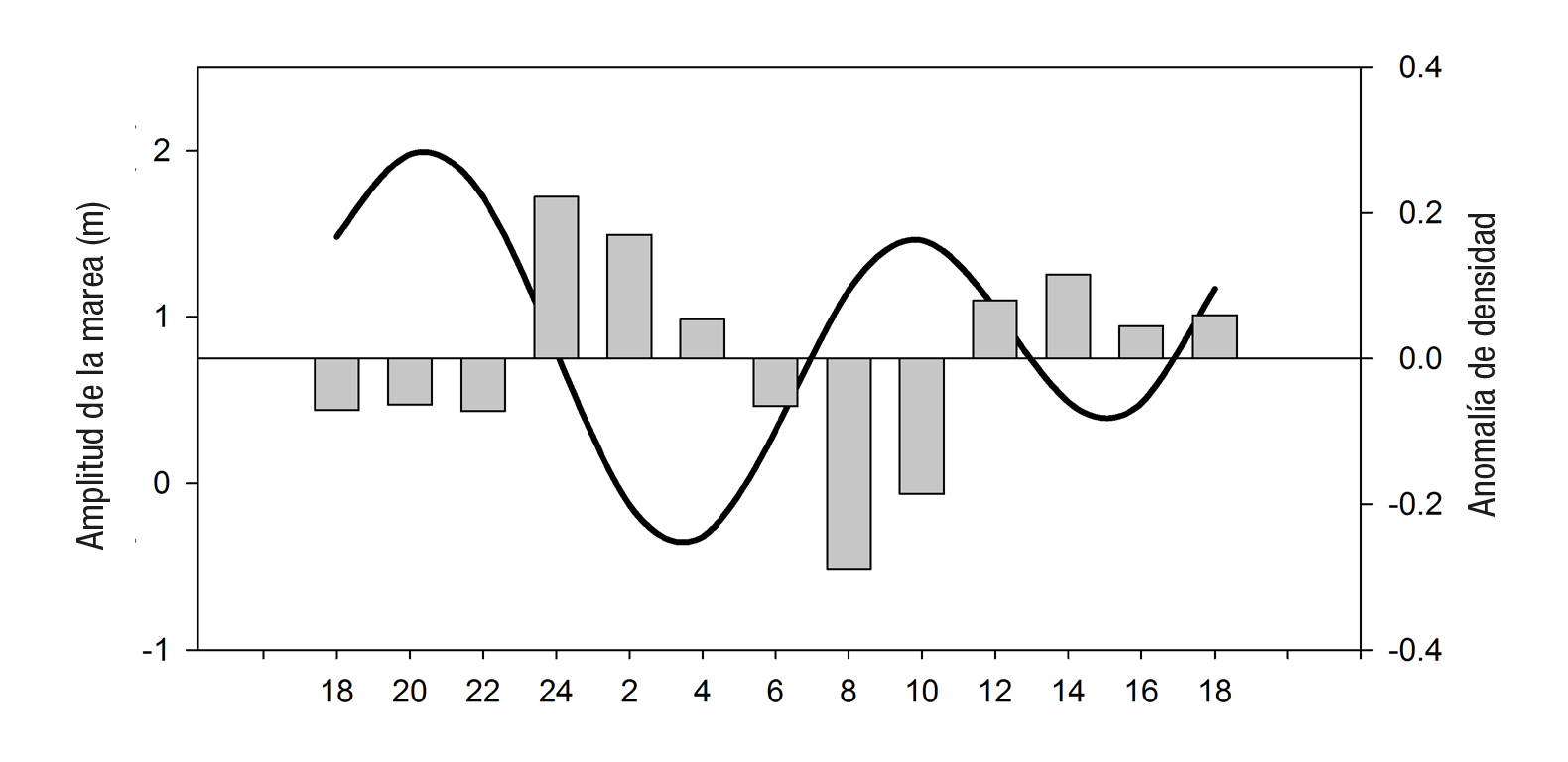
Figura 7 Variación diurna de la amplitud de marea y diferencias de densidad en la capa a 30 m medidos en la boca de Bahía Magdalena, México. Los valores positivos corresponden al reflujo y los negativos al flujo.
A diferencia de otros sistemas lagunares, la costa occidental de Baja California presenta lagunas costeras más o menos profundas con bocas amplias con un transporte de material disuelto y particulado comparativamente mayor, que ayuda a explicar su alta productividad durante el año. El metabolismo neto (autótrofo / heterótrofo) de esta laguna, evaluado por el modelo Land Ocean Interactions in the Coastal Zone (LOICZ) (Gordon et al., 1996), mostró que en el periodo de fe brero a julio (surgencias intensas) tiende a ser autótrofo (fotosíntesis > respiración) y de agosto a enero (surgencias débiles) es heterótrofo (fotosíntesis < respiración) (Cervantes-Duarte, 2016). En las lagunas costeras localizadas a lo largo de la costa del noroeste de la península de Baja California, México, el océano es la fuente externa más impor tante de nutrientes y de carbono orgánico e inorgánico (Ribas-Ribas et al., 2011), por lo que sus lagunas actúan como sumidero de nutrientes, similar a lo reportado para Bahía Magdalena, donde el NID es sumi dero todo el año y el FID es fuente en el periodo de agosto a enero (Cervantes-Duarte, 2016). Los resultados en la escala diurna difieren de los reportados en la escala estacional. En principio porque los primeros consideran a la marea como la principal fuente de variación, mientras que en la escala estacional los procesos climático-oceanográficos son más relevantes. Este estudio debe considerarse como una primera aproximación del transporte diurno de nutrientes y clorofila a, donde dependiendo de los cambios de la marea (viva y muerta) y la época del año, el balance de nutrientes definirá en gran medida la productividad del ecosistema.

