











 nova página do texto(beta)
nova página do texto(beta) Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO 
 Permalink
PermalinkINTRODUCTION
The toad Incilius valliceps (Wiegmann, 1833) is common on Cozumel Island, and their distribution ranges widely throughout the states of Oaxaca, Chiapas, and the Yucatan Peninsula in Mexico (Ochoa-Ochoa et al., 2006). Although this toad is not yet considered threatened or endangered (Rojas-Canales & Ríos-Valdez, 2012), the decline of am phibian species, in general, is of great concern, especially in Latin America (Young et al., 2000). Information regarding tadpole food pre ferences is scarce, but they often act as detritivores, herbivores, car nivores, or omnivores (Altig et al., 2007). By examining the gut con tents, we can better understand the dietary needs of tadpoles and thus obtain a complete understanding of the ecological role that they play. The ecological behavior of freshwater zooplankton is affected by a wide variety of biotic and abiotic factors. Predation is known to in fluence the behavior of zooplankton and produces effects that can be noticed in the population size, the production of ephippial (res ting) eggs, and the distribution of zooplankton throughout the lim netic zone (Lampert & Sommer, 2007). The effect of fish, copepod, freshwater shrimp, and insect larva predation on communities of zo oplankton has been well documented (O’Brien, 1979; Vijverberg et al., 1990; DeVries & Stein, 1992; Gliwicz & Umaña, 1994; Jawahar-Ali et al., 1996; Fernando, 2002; Romare et al., 2003; Cervantes-Martínez, 2005). However, zooplankton interaction with tadpoles remains lar gely unstudied; only a few studies have investigated this relationship, such as the one performed by Hamilton et al. (2012) that identified the predator-prey relationship that exists between cladocerans (Daphnia pulex Leydig, 1860) and tadpoles (Rana aurora Baird & Girard, 1852) and recognized the importance of cladocerans in the tadpole diet. Given the lack of information related to the feeding habits of tropical ta dpoles and their relationship with zooplankton, we aimed to identify the dietary preferences of I. valliceps tadpoles and the way in which their presence influenced zooplankton richness and abundance in a small artificial pond, where these organisms are present throughout the year.
MATERIALS AND METHODS
Sample collection. We undertook biological collections from one artifi cial pond located on the campus of the University of Quintana Roo (20˚ 29’ 18.3” N and 86° 56’ 22.8” W) on Cozumel Island, Mexico, between June 2011 and January 2012, covering the entire rainy season which favors anuran oviposition (Cedeño-Vázquez et al., 2006). The volume of the pond remained nearly constant (4 m3), had a depth < 1.5 m, and was filled exclusively with rain water.
We collected zooplankton samples monthly using a van Dorn bottle; we filtered 5 L of water through a zooplankton net with a mesh size of 45 µm. We took duplicate samples at a depth of 0.5 m and preserved them according to standard methods for further observation (Elías-Gutiérrez et al., 2008).
We took samples of I. valliceps tadpoles once a month for three months, using a mosquito-netting-like sieve to collect as many tadpo les as possible within 5 minutes. We introduced the net to the center of the system and, using a weight, made it rest on the bottom. After the specified time had passed, we removed the net in a swift upward motion trapping any tadpoles that had rested on the bottom and those found in the water column. We captured between 15 to 30 individuals in the months in which tadpoles were present (July, September, and October), with the exception of July (only six individuals). We preserved all tadpoles following standard methods for the posterior gut content analysis (Dutra & Callisto, 2005).
Laboratory. To evaluate the prey availability in the system, we analyzed the monthly average zooplankton abundance and diversity (S). We es timated the abundance of each zooplankton species using total counts (Lewis, 1979), or the aliquot method for those samples that had ex tremely large quantities of zooplankton (Oscoz et al., 2006). In order to identify zooplankton species, we used taxonomic keys from Koste (1978), Victor (2002), Kutikova (2002), and Elías-Gutiérrez et al. (2008). We calculated the availability of each zooplankton species as the proportion of each species, relative to the total number of individuals pertaining to the zooplankton community. We quantified the diversity of zooplankton species using the Shannon-Wiener Index (Shannon & Weaver, 1949). Shannon-Wiener Index: (H’) = -∑ pi lnpi, where pi is the relative abundance of the species i.
In order to describe tadpole predation on zooplankton, we recorded morphological magnitudes for each tadpole: body length, tail length, and total length (Altig & McDiarmid, 1999). The intestinal contents of the tadpoles were analyzed according to Dutra & Callisto (2005), and the proportional abundance of zooplankton species in the tadpole intestines was estimated by total count.
We calculated the selectivity of zooplankton species by each ta dpole as a monthly average using the Jacobs selectivity index (Jacobs, 1974): D= (r-p) / ((r+p)-2rp)), where r is the proportion of a specific prey species in the gut contents and p is the proportion of the specific prey species in the environment. This index (D) ranges from -1 (nega tive selection), 1 (positive selection), and 0 (no selection) (Lampert & Sommer, 2007).
We performed a simple linear regression in Statgraphics Centurion v. 10.0 to test if tadpole size played a role in the amount of zooplankton consumed. We transformed the data using log (x+1).
RESULTS
Eight zooplankton species were identified as available in the medium: six rotifers:Epiphanes brachionus (Ehenberg,1837), Anuraeopsis fis sa Gosse,1851, Keratella americana Carlin, 1943, Lepadella patella (Müller,1773), Lecane luna (Müller,1776), and an unidentified species of Bdelloidea group, one cladoceran, i.e., Moina wierzejskii (Richard, 1895), and one ostracod, i.e., Cypridopsis vidua (Müller,1776).
In the zooplankton community, the Shannon-Wiener diversity index (H’) ranged from 0.015 to 1.27 (Fig. 1). In August, October, and Dec ember, we confirmed the presence of dominant species (the species of unidentified bdeloideo and A. fissa respectively) and lower values in H’. Meanwhile, the months with higher richness (S) and diversity were June (S= 5, H’=1.27), September (S=5, H’=0.98), and November(S=5, H’= 1) (Fig. 1).
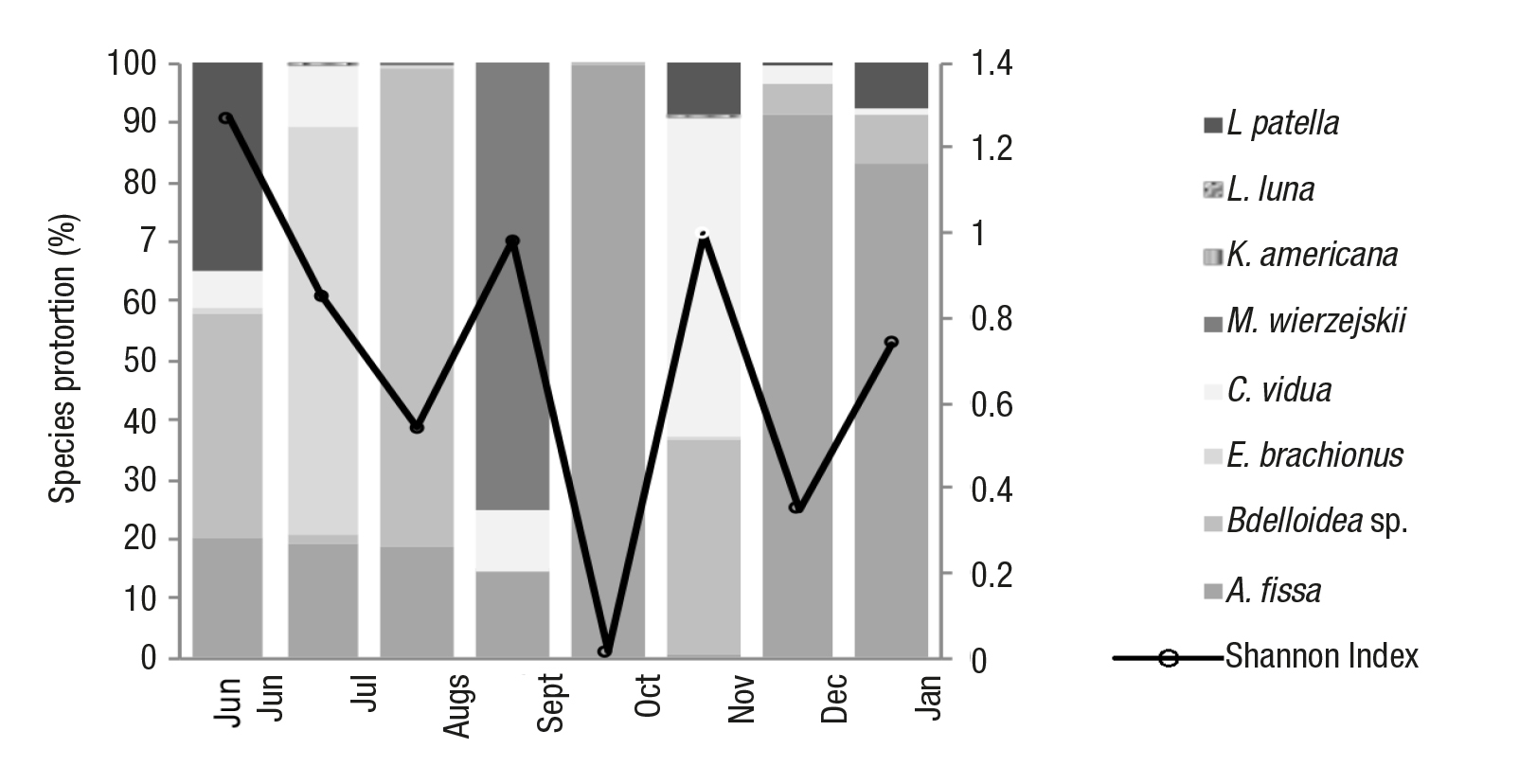
Figure 1 Proportion of every zooplankton species in the environment (for every month (columns) of the study and the corresponding Shannon-Wiener Index values (rows), found in an artificial aquatic pond on Cozumel Island, Mexico.
The availability of zooplankton species in the artificial aquatic system varied throughout the study period. E. brachionus was greatly available in the month of July (68.71%), while A. fissa was available in all months, but especially in October (99.81%), December (91.52%), and January (83.25%). K. americana, L. luna, and L. patella were the species with the least availability throughout the period of study (maxi mum: 0.06%, 0.01%, and 34.78%, respectively). C. vidua was also available throughout the study period in small proportions with the hig hest availability recorded in November (53.43%). M. wierzejskii was only available in August (0.55%) and September (74.95%) (Fig.1).
Only Bdelloidea sp., C. vidua, M. wierzejskii, A. fissa, and E. bra chionus were consumed by I. valliceps, and the species found in higher proportions in the tadpole gut contents were C. vidua (July 83.19%, September 26.54%, and October 36.28%) and. M. wierzejskii (Septem ber 69.63% and October 13.5%) (Fig. 2).
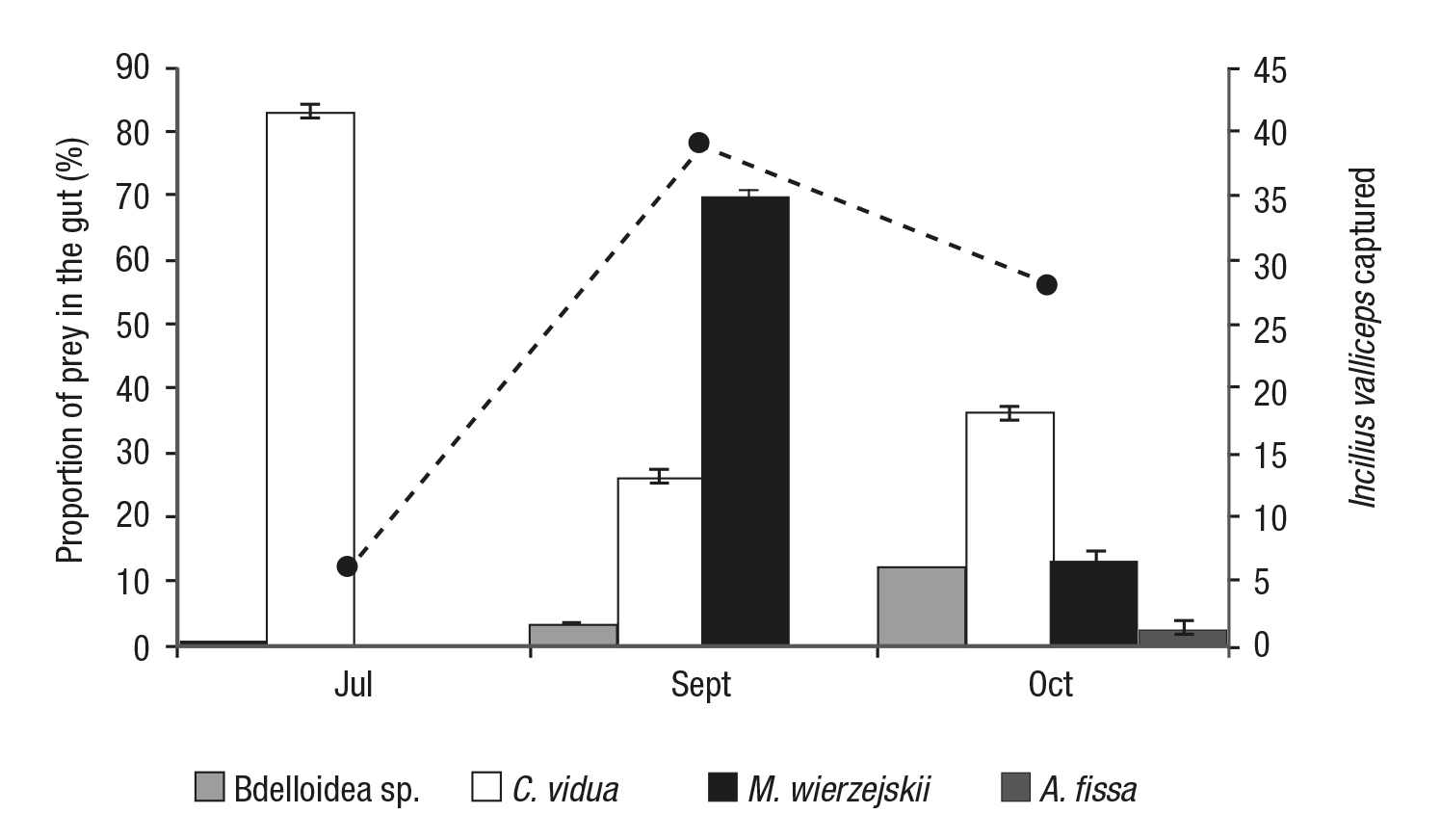
Figure 2 Proportion of each zooplankton prey species present in the tadpole gut contents in each month (columns) and the number of Incilius valliceps tadpoles captured in each month (rows), found in an artificial aquatic pond on Cozumel Island, Mexico.
In July, C. vidua was more abundant in the tadpole guts (83.19%) when the availability was relatively low in the environment (10.64%). While in October, C. vidua was found frequently in the gut contents of the tadpoles (36.28%), but occurred rather rarely in the pond (0.02%). The tadpoles examined in September mainly contained M. wierzejs kii (69.63%), which was also the most abundant zooplankton species in the environment (74.95%). The species of bdeloidea was not very abundant in the environment when tadpoles were present (July: 1.26%, September: 0.38%, and October: 0.08%) and was also only observed in the gut contents in small proportions (July: 0.14%, September: 3.26%, and October: 12.19%) A. fissa was abundant in all three months (July: 19.26%, September: 14.34%, and October: 99.88%), but was absent in all the gut contents with the exception of October (2.75%) (Figs. 1-2).
The Jacobs selectivity index provided insight into the feeding prefe rences of I. valliceps tadpoles in July, September, and October. C. vidua was positively selected; while M. wierzejskii changed from not being actively selected (no selection) in September to being positively selec ted in October; E. brachionus, bdeloidea, and A. fissa were altogether negatively selected (Fig. 3).
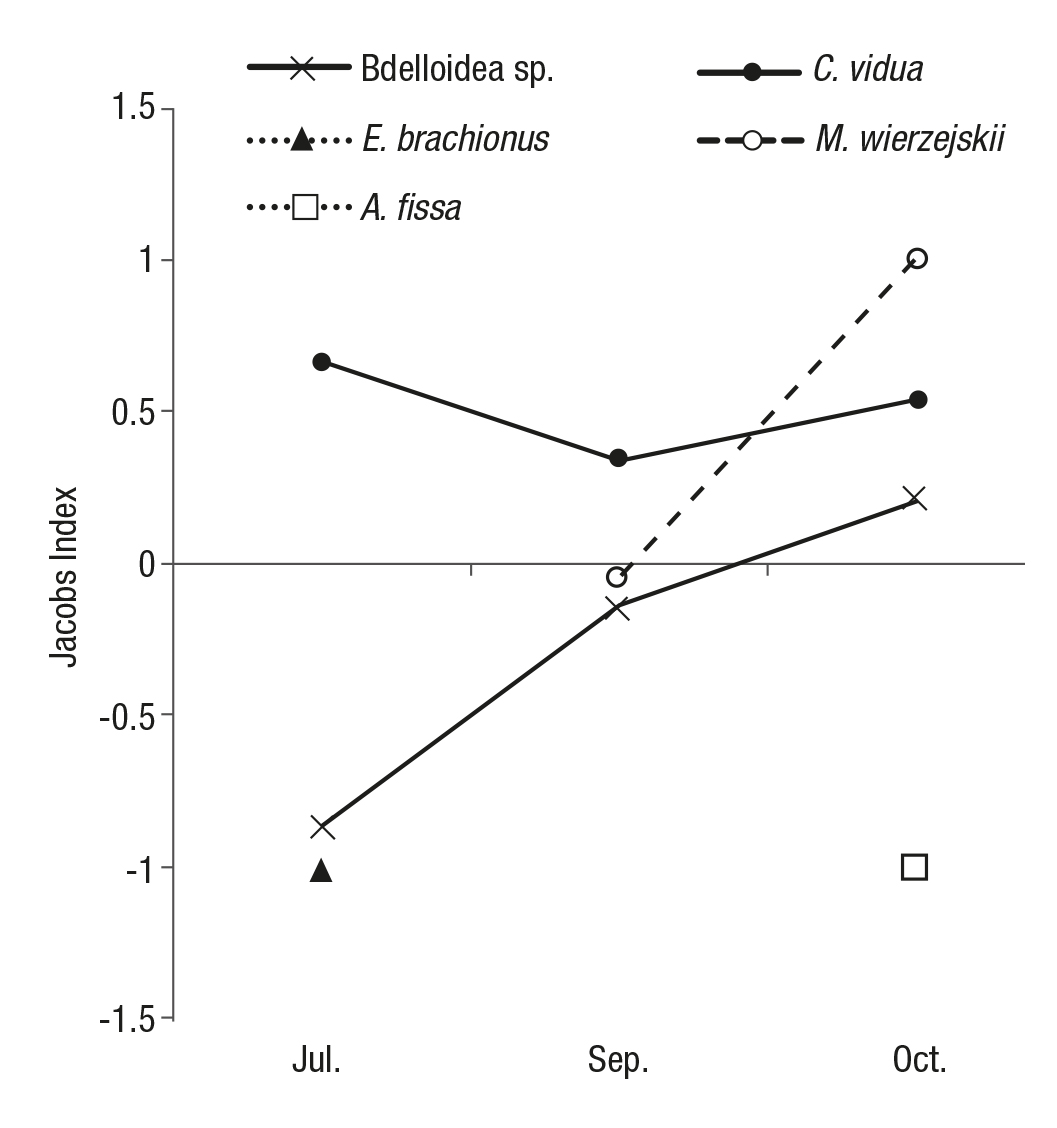
Figure 3 Jacobs selectivity index (monthly average) for every zooplankton spe cies preyed on by the tadpoles each month, found in an artificial aquatic pond on Cozumel Island, Mexico.
The simple linear regression applied revealed the existence of a statistically significant relationship between tadpole size and the num ber of C. vidua (R2 = 0.215, p = 0.00256; Fig. 4a), M. wierzejskii (R2 = 0.1604, p = 0.02021; Fig. 4c), and bdeloidea consumed (R2 = 0.2609, p = 0.0175; Fig. 4b). Larger tadpoles ingested more C. vidua and bdeloi dea and smaller tadpoles consumed slightly more M. wierzejskii.

DISCUSSION
The abundance and presence of tadpoles in aquatic systems is de termined by where adult anurans choose to lay their eggs. Tadpoles are entirely aquatic organisms, meaning that adults must lay their eggs in an aquatic medium where water and resources are abundant, to ensure their development and metamorphosis (Cedeño-Vázquez et al., 2006).
The artificial aquatic system examined in this study contains an es tablished zooplankton community, which explains the relative stability in the number and type of species present. Aquatic systems often reach equilibrium in terms of the number of species present as they age (Do dson et al., 2007; Alfonso et al., 2010). This particular aquatic system is approximately eight years old and, as documented in this study, A. fissa, bdelloidea, and C. vidua are always present (established species). Species richness each month was always between four to six species. This means that a constant and predictable supply of food is available for the tadpoles inhabiting this pond.
The Jacobs selectivity index applied to the gut contents of the ta dpoles showed that the tadpoles of I. valliceps clearly preferred C. vidua throughout the duration of this study. The positive values of the linear regression between the size of the predator and the amount of C. vidua ingested suggest that the tadpoles preferred to consume this ostracod throughout their entire growth and development. The importance of C. vidua to the tadpole diet is a reason to perform experimental tests in a controlled environment.
Furthermore, the relationship between tadpole length and the num ber of M. wierzejskii consumed demonstrates that this species was highly consumed by the young tadpoles, but consumption of cladoce rans diminished as the tadpoles matured and the availability changed. Sousa Filho et al. (2007) have previously reported slight differences in diet, depending on the size and larval stage of tadpoles.
Although ostracods are known for partially swimming and crawling along the bottom of aquatic systems (Cohen et al., 2007), C. vidua is an efficient swimmer that can enter into the limnetic zone, and is relatively large when compared to rotifer species (Victor, 2002), which may cause this species to be easily seen by the tadpoles. As has been recorded in fish (visual predators), larger prey are positively selected in an effort to conserve energy (Lynch, 1979; Owili & Omondi, 2003; Gallardo Alanis et al., 2009). This may explain why tadpoles positively selected the lar ger zooplankton species (C. vidua and M. wierzejskii) in this artificial aquatic system.
Even though the tadpoles did not exhibit a clear preference for bde loidea throughout the study, individuals measuring 4.7 cm consumed a greater quantity of this rotifer species than their counterparts did. Bdelloidea, A. fissa, and E. brachionus were present in the tadpole gut contents, but based on the results of the selectivity index, these species were not preferred. Rotifers are smaller than cladocerans, copepods, or ostracods; as a result, invertebrate predators, rather than vertebrates such as fish or tadpoles (Herzig, 1987), often select them. The fact that these rotifer species were present in the gut contents, available to the tadpoles, but not actively selected by them, suggests that they were incidentally ingested.
K. americana, L. patella, and L. luna were the species with the lowest availability or were not detected in the environment when the tadpoles were present. Rotifers have been previously documented as an important source of protein for tadpoles (Rossa-Feres et al., 2004). Yet they also exhibit special behavior such as entering into, or remai ning in, a state of diapause (Schröder, 2005), or the ability to stay at lower abundances (Walz, 1997) in the presence of predators as stra tegies to avoid predation, ensuring that they maintain permanence in the community.
The sharp decline in the population of M. wierzejskii observed in the months of September and October (Fig. 1) and the production of males and ephippial eggs can be attributed to tadpole predation of this zooplankton species (supported by the gut content analysis). Predation has been known to affect the life history and behavior of tropical clado cerans (Sarma et al., 2005).
In September, M. wierzejskii was abundant in the aquatic system and in the gut contents of the tadpoles. Meanwhile in October, M. wier zejskii was absent in the ecosystem but we continued to find individuals in the gut contents of the tadpoles. In the following months, we did not find any individuals in the gut contents or in the environment.
We also observed M. Wierzejskii with resting eggs in the gut con tents of the tadpoles as well as in the environment in the month of Sep tember. Males pertaining to this cladoceran species were also present in the environment. Males and resting eggs are often only produced when the zooplankton population finds itself living under environmental stress such as lack of nutrients, competition, and predation (Slusarczyk, 1999; Lampert & Sommer, 2007).
Considering the small area of the system, and the fact that only one predator dwells within it (personal observation), we hypothesize that the tadpoles produced a profound diminishing effect on the cladoce ran population. Tadpoles caused them to reproduce sexually, eventually leading to their disappearance from the artificial aquatic system, which has also been recorded in similar studies (Hamilton et al., 2012; Gama-Flores et al., 2013).
Our demonstration in this study regarding tadpole affinity for ma crozooplankton, and the way in which these aquatic vertebrates in fluence zooplankton richness and abundance represents a great advan ce in understanding and defining the trophic status and dietary needs of tadpoles as well as the ecological role that they play within the aquatic systems they inhabit. This knowledge is essential in the conservation of not only I. valliceps, but also of other anuran species and the ecosys tems in which they reside.

