











 nova página do texto(beta)
nova página do texto(beta) Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO 
 Permalink
PermalinkIntroducción
Los lagos tropicales de alta montaña se caracterizan por ubicarse por arriba de la cota de vegetación arbórea (timberline = TL). La altitud a la que se observa la TL depende de la latitud, y se encuentra a mayor altitud en latitudes más bajas (Hutchinson, 1975; Sommaruga, 2001; Kernan et al. 2002). En la zona tropical se localiza por arriba de los 3 000 m s. n. m. (Margalef, 1983; Burgis & Morris, 1987). Este tipo de lagos se encuentra en islas de clima frío dentro de las zonas tropicales. La mayor parte de los lagos de alta montaña tropicales se localiza en la zona de los Andes, y en menor cantidad, en el este de África (Löffler, 1964; Payne, 1986). Este tipo de lagos presenta analogías ambientales con los lagos nórdicos de latitudes altas. La similitud principal entre estos dos tipos de lagos es la temperatura, la cual disminuye conforme incrementan la latitud y la altitud (Thomasson, 1956; Lewis, 1983; Steinitz-Kannan et al., 1983; Burgis & Morris, 1987; Lewis, 1987 y 1996).
Los macroinvertebrados bentónicos (MIB), son una comunidad importante debido, entre otros aspectos, a su papel en la remineralización de la materia orgánica (Ivanov, 2000; Simčič & Brancelj, 2002). En los lagos de alta montaña, las variaciones en tiempo y espacio de los MIB están determinadas por los parámetros físicos (temperatura, transparencia, textura sedimentaria), químicos (conductividad, pH, oxígeno disuelto) y biológicos (alimento, cobertura vegetal, interacciones), que se presentan en todos los cuerpos acuáticos (Newrkla & Wijegoonawardana, 1987). Sin embargo, es frecuente que dentro del mismo cuerpo acuático se presenten condiciones diferentes en la zona profunda y la litoral. El efecto del viento y el oleaje, la penetración de la luz, el calentamiento gradual del agua, el tipo de sustrato y la presencia de vegetación sumergida son sólo algunas de las variables que marcan la diferencia entre ambas zonas (Brinkhurst, 1974; Ivanov, 2000).
Estas variables inducen a las especies de la comunidad bentónica a desarrollar estrategias adaptativas para dominar o influir sobre otras especies, como entrar en un estado de latencia hasta el restablecimiento de condiciones fisiológicamente más favorables para un desarrollo mejor (Wetzel & Likens, 2000). Los MIB presentan adaptaciones que les permiten resistir condiciones extremas generalmente presentes en los lagos de alta montaña (como pH ácido, temperatura baja y exposición elevada a rayos UV), con lo cual evitan la depredación y competencia de otras especies no resistentes a dichas condiciones (Kobuszewski & Perry, 1994; Hider et al., 1999), por lo que su abundancia es mayor que en condiciones normales.
La abundancia o riqueza específica de los MIB en lagos tropicales frecuentemente son menores a las encontradas en los lagos templados (Lewis, 1996). Las especies bentónicas que han sido reportadas para los lagos de alta montaña son más similares a las especies halladas en lagos de latitudes templadas que a las presentes en lagos de latitudes tropicales de menor altitud (Burgis & Morris, 1987).
Además, los lagos de alta montaña son considerados muy sensibles a las perturbaciones humanas indirectas, particularmente a los efectos de la acidificación. Sin embargo, para poder reconocer las perturbaciones antropogénicas, primero es necesario conocer la variabilidad natural de las comunidades.
En México se conocen sólo dos lagos de alta montaña: El Sol y La Luna, en el cráter del volcán Nevado de Toluca, en el Estado de México. Dadas las condiciones ambientales extremas en estos lagos, se espera que la composición taxonómica de los macroinvertebrados bentónicos (MIB) sea reducida y que, para el caso de El Sol, las condiciones ambientales de la zona profunda y litoral no modifiquen la estructura de la comunidad de los macroinvertebrados bentónicos (MIB), dado lo somero del lago (<15 m). Por lo anterior, el objetivo del presente trabajo fue evaluar la distribución espacial y temporal de la comunidad de los MIB en los lagos La Luna y El Sol, en el centro de México. Asimismo, se analizó la relación del comportamiento temporal de los parámetros fisicoquímicos de cada uno de los lagos y cómo afectan la composición taxonómica y la distribución de los macroinvertebrados bentónicos (MIB).
Materiales y métodos
Área de estudio. Como se mencionó anteriormente, en México se conocen sólo dos lagos de alta montaña: La Luna y El Sol, en el cráter del Nevado de Toluca, en el Estado de México. Son lagos-cráter tropicales de alta montaña que se encuentran dentro de la cuenca hidrológica prioritaria Nevado de Toluca (Arriaga et al., 1998), localizada entre los 19° 06” 36’ N y 18° 48” 00’ N y los 99° 54” 36’ O y 99° 28” 48’ O, a una altitud de 4 240 m s. n. m. El clima de la zona va de frío a semifrío húmedo, con una temperatura media mensual que oscila entre 2 y 12 °C. Presenta una precipitación anual de entre 1 200 y 2 000 mm y una evaporación potencial promedio anual de 990 mm (García, 1988a).
La profundidad máxima de La Luna es de 10 m (profundidad media de 5 m), con una superficie de 31 083 m2 (largo 227 m, ancho 209 m), y la profundidad máxima de El Sol es de 15 m (profundidad media de 6 m), con una superficie de 237 321 m2 (largo 795 m, ancho 482 m) (Alcocer et al., 2004).
Ambos lagos tienen aguas transparentes, un pH ácido (4.9 5.6) y una capacidad de amortiguación reducida. La temperatura promedio del agua oscila entre los 9 y 11 °C en La Luna, y en El Sol, entre los 7 y 13 °C. El contenido de oxígeno disuelto es alto (6.0 7.8 mg L-1) en ambos lagos, y su conductividad, baja (18 24 μS cm-1). Presentan un contenido bajo de materia orgánica disuelta y en suspensión (Caballero, 1996; Armienta et al., 2000).
Trabajo de campo. El muestreo se realizó mensualmente durante un ciclo anual (de abril de 2000 a marzo de 2001). Las estaciones de muestro se eligieron con base en un muestreo prospectivo, el cual consistió en tomar partes de sedimento de ambos lagos para determinar la presencia de MIB. En La Luna se estableció una sola estación de muestreo a los 10 m (Luna), ya que los sedimentos de la zona litoral estaban muy compactados, lo que impide el establecimiento de invertebrados bentónicos. En El Sol se establecieron dos estaciones profundas (Sol1 = 10 m y Sol2 = 12 m) y dos en la zona litoral, una en sustrato desnudo (sin vegetación) (Sol3 = 0.5 m) y la otra con vegetación enraizada sumergida (Sol3v = 0.5 m) (Fig. 1).
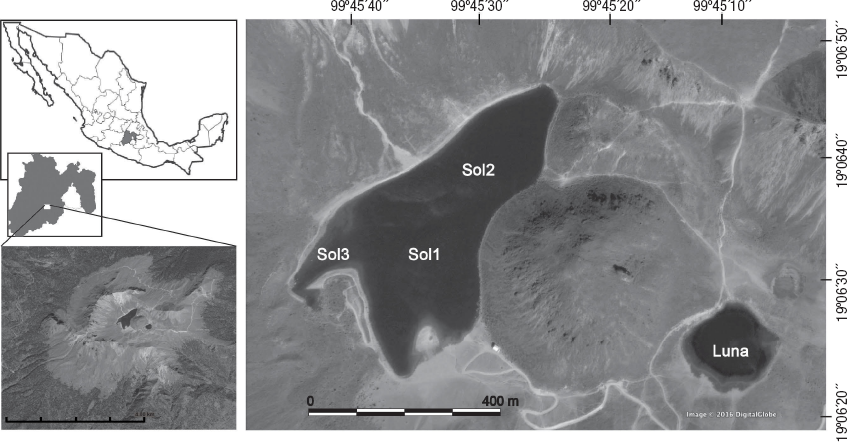
Figura 1 Lagos El Sol y La Luna en el cráter del Volcán Nevado de Toluca, México, mostrando las cuatro estaciones de muestreo.
Los variables ambientales que se midieron in situ cerca del sedimento fueron temperatura, pH, concentración y porcentaje de saturación de oxígeno disuelto y conductividad específica (K25), con ayuda de una sonda multiparamétrica de calidad del agua y un sistema de captura y almacenaje de información marca Hydrolab modelos DS4 y SVR4 respectivamente. Asimismo, se obtuvieron muestras de sedimento para el análisis del tamaño de grano, el contenido de materia orgánica, la concentración de clorofila a y la biomasa vegetal.
En las estaciones profundas la obtención de los MIB se realizó con una draga Ekman (0.0225 m2 de área) (Weber, 1973; Wetzel & Likens, 2000), mientras que en las estaciones de la zona litoral se tomaron con un nucleador de acrílico (0.0019 m2 de área). Los sedimentos se tamizaron por medio de una malla de 0.42 mm para retener al macrobentos (Greenberg et al., 1985; Wetzel & Likens, 2000), posteriormente, se fijaron con alcohol a 96%, adicionado con rosa de Bengala como colorante vital (Greenberg et al., 1985).
Trabajo de laboratorio. El tamaño de grano se determinó por el método de tamizado en seco (60 °C) para las arenas, y de pipeteo en húmedo para limos y arcillas (Folk, 1969). Para determinar el porcentaje de materia orgánica sedimentaria se utilizó la técnica de pérdida de peso por combustión (LOI) (550 °C) (Greenberg et al., 1985). La biomasa vegetal se obtuvo con el peso seco de la vegetación encontrada en cada uno de los núcleos de donde se separaron los MIB. Finalmente, la medición de la concentración de clorofila a del sedimento se hizo siguiendo la técnica 445.0 de la EPA, con un fluorómetro digital Turner Designs modelo 10-AU (Arar & Collins, 1997). Para transformar la concentración de μg de clorofila a L-1 a μg de clorofila a cm-2 se tomó la concentración integrada de un núcleo de sedimento de un cm3. En el laboratorio se llevó a cabo la separación de los organismos y se identificaron en primera instancia con base en las claves generales de Thorp y Covich (1991), Edmondson (1959) y Pennak (1953). Posteriormente, se emplearon claves específicas para los oligoquetos (Stimpson et al., 1982; Hiltunen & Klemm, 1980; Brinkhurst & Marchese, 1992) y quironómidos (Mason, 1973; Wiederholm, 1983). Todos los organismos fueron conservados en alcohol glicerinado a 70%.
Análisis de la información. Para determinar la presencia de estacionalidad durante el ciclo de muestreo, se aplicó un análisis de conglomerados (AC) por el método de Ward, con distancias euclidianas para la zona profunda de La Luna y para la zona somera y profunda de El Sol, con el fin de agrupar los meses similares en cada zona con base en las características ambientales del agua y sedimento (Gauch, 1982). Asimismo, se realizó un análisis de componentes principales (ACP) con el fin de identificar los parámetros que más influencia tienen sobre el ordenamiento (Gauch, 1982; García, 1988b).
La densidad de los organismos se calculó por metro cuadrado, multiplicando la abundancia de cada especie por un factor de conversión (área de la draga = 0.0225 cm2 y área del nucleador = 0.0018 cm2). Para los análisis estadísticos, las densidades se transformaron a logaritmos [log10 (n+1)] para cumplir con las suposiciones de normalidad y homocedasticidad de los datos (Elliot, 1983). Para reconocer si la densidad de los MIB era diferente entre sí, se realizó una prueba t-Student para las estaciones de la zona litoral de El Sol, y un análisis de varianza (ANOVA) de una vía para las estaciones profundas (Luna, Sol1 y Sol2) (Dytham, 2011; Elliot, 1983). Para determinar si las estaciones profundas eran diferentes, se aplicó la prueba post hoc de Tukey. Se realizaron análisis de correlación canónica (ACC) entre las variables ambientales y las biológicas para la zona profunda de La Luna, para la zona profunda de El Sol y para la zona somera de El Sol. Lo anterior, con el fin de reconocer qué variables ambientales se asociaban directamente con la variación temporal de los MIB (Garson, 2015; Jeffers, 1978). Todas las pruebas se elaboraron con ayuda del paquete estadístico SPSS v10.
Resultados
Parámetros ambientales. La temperatura promedio anual, máxima y mínima de las tres estaciones profundas (Luna, Sol1 y Sol2) fueron muy similares. La temperatura promedio anual de la zona litoral (Sol3 y Sol3v) fue más elevada (≈1.5 °C), y se observó la mayor diferencia en las máximas. El pH promedio anual fue ácido para ambos lagos, y menor en La Luna, donde se encontraron valores < 5 durante casi todo el año. El promedio de oxígeno disuelto fue muy similar en ambos lagos; se encontró alrededor de la saturación durante la mayor parte del año. La conductividad fue ligeramente más elevada en El Sol que en La Luna. En cuanto al sedimento, tanto la textura como el contenido de materia orgánica fueron mayores en la zona profunda de ambos lagos que en la zona litoral de El Sol. La clorofila a de la zona profunda de El Sol fue casi el doble de la encontrada en la zona litoral del mismo lago y de la zona profunda de La Luna (Tabla 1).
Tabla 1 Características ambientales del agua del fondo y del sedimento de las estaciones profundas del lago El Sol (Sol1 y Sol2) y someras (Sol3 y Sol3v) del lago La Luna del Parque Nacional “Nevado de Toluca”, en la porción central de México.
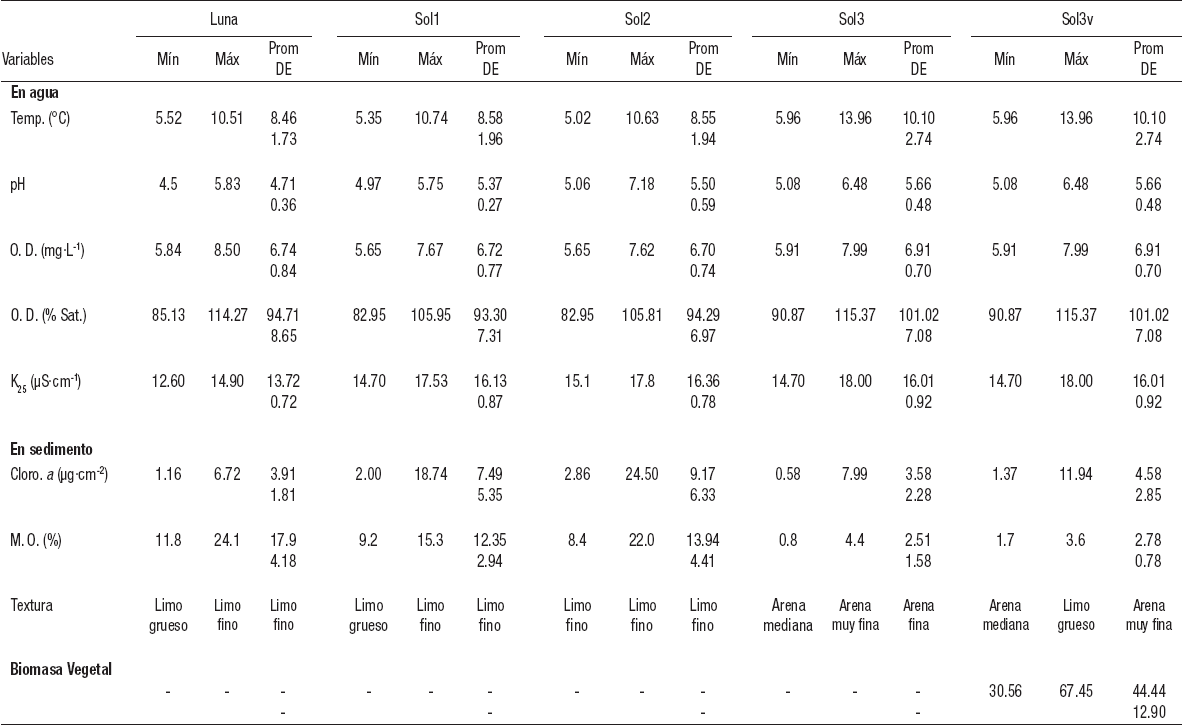
Prom = Promedio, DE = Desviación estándar. Máx = Valor máximo. Mín = Valor mínimo. Temp. = Temperatura. O.D. = Oxígeno disuelto. K25 = Conductividad estandarizada a 25°C. Cloro. a = Clorofila a. M.O. = Materia orgánica.
De manera general, en los dendrogramas de similitud obtenidos del AC de las variables ambientales de las tres zonas se forman dos épocas (cálida y fría) con una línea de corte de la escala de similitud a 25 (Figs. 2a, 2b y 2c). En los ACP, los primeros dos componentes explican el 100% de la varianza. El aporte mayor se da en el primer componente, tanto para la zona profunda de La Luna (99.4%) como para la zona profunda (99.8%) y somera (99.7%) de El Sol. El oxígeno disuelto fue la variable que mayor peso tuvo en el primer componente de la zona profunda para ambos lagos, y de la zona litoral para el caso de El Sol. Para la zona profunda de La Luna fue la materia orgánica la que mayor peso tuvo en el componte dos, mientras que para la zona profunda de El Sol fue la clorofila a sedimentaria. La temperatura fue la variable que mayor peso tuvo en el componente dos para la zona somera de El Sol.
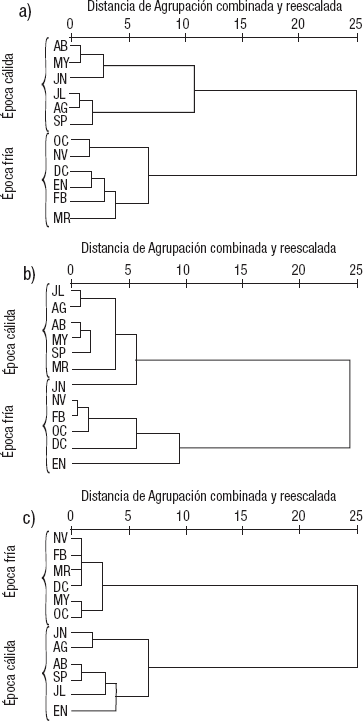
Figuras 2a-c Dendrogramas de similitud (Método de Ward) de los parámetros ambientales del lago La Luna (a) y la zona profunda (b) y somera (c) del lago El Sol del volcán Nevado de Toluca, en la porción central de México.
De acuerdo con lo anterior, la época cálida presentó un menor porcentaje de oxígeno disuelto y una mayor cantidad de materia orgánica. Además, tuvo una temperatura más elevada y una concentración de sólidos disueltos (K25) más baja. La época fría se caracterizó por tener un porcentaje de oxígeno disuelto elevado y una cantidad de materia orgánica más baja, así como una temperatura menor y una conductividad mayor.
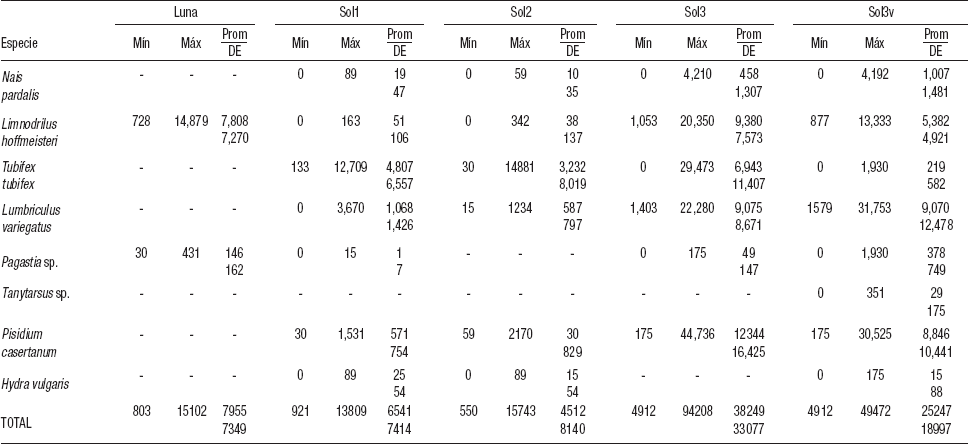
Parámetros biológicos. La riqueza específica en la zona profunda de La Luna fue de dos especies, un oligoqueto (Limnodrilus hoffmeisteri Claparède, 1862) y un quironómido (Pagastia sp.). En la zona profunda de El Sol (Sol1 y Sol2), se encontraron cuatro especies de oligoquetos (L. hoffmeisteri, Nais pardalis Piguet, 1906, Tubifex tubifex Muller, 1774 y Lumbriculus variegatus Müller, 1774), una especie de bivalvo (Pisidium casertanum Poli, 1791) y una de hidrozoario (Hydra vulgaris Pallas, 1766). Adicionalmente, en Sol1 se registró una especie de quironómido (Pagastia sp.). En la zona somera de El Sol (Sol3 y Sol3v), se encontraron las cuatro especies de oligoquetos (N. pardalis, L. hoffmeisteri, T. tubifex y L. variegatus), la del bivalvo (P. casertanum) y la del quironómido (Pagastia sp.). En Sol3v se encontró, además, el hidrozoario (H. vulgaris) y otra especie de quironómido (Tanytarsus sp.) (Tabla 2).
Tabla 2 Densidad (org×m2) de los macroinvertebrados bentónicos del lago La Luna y las estaciones profundas (Sol1 y Sol2) y someras (Sol3 y Sol3v) del lago El Sol del Parque Nacional “Nevado de Toluca”, en la porción central de México. Prom = Promedio. DE = Desviación estándar. Máx = Valor promedio máximo. Mín = Valor promedio mínimo.
De acuerdo con el ANOVA, no se encontraron diferencias significativas entre las densidades promedio anuales de las comunidades de MIB de la zona profunda de ambos cuerpos de agua (p = 0.103; n = 36; F = 2.328). Asimismo, con la prueba t no se encontró evidencia significativa (p = 0.267; n = 36; t = 1.539) para decir que la densidad promedio anual de las poblaciones de los MIB de la zona litoral de El Sol (Sol3 y Sol3v) fuera diferente. A pesar de que no se puede comparar estadísticamente, gráficamente se aprecia que en El Sol las densidades promedio de las estaciones de la zona litoral (Sol3 y Sol3v) son mucho mayores a las encontradas en la zona profunda (Sol1 y Sol2) (Tabla 2, Fig. 4 [Fig. 3]).
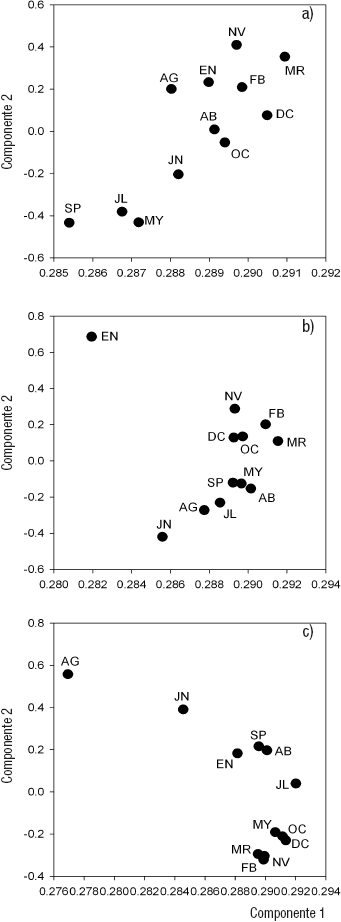
Figuras 3a-c Análisis de Compontes Principales de los parámetros ambientales de el lago La Luna (a) y de la zona profunda (b) y somera (c) de el lago El Sol del volcán Nevado de Toluca, en la porción central de México.
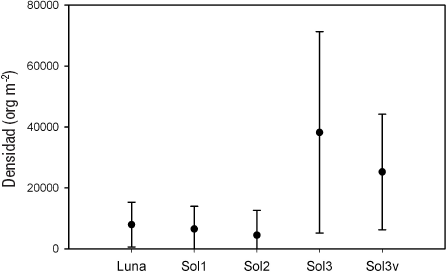
Figura 4 Densidad promedio anual de los macroinvertebrados bentónicos de las estaciones de los lagos tropicales de alta montaña El Sol y La Luna del volcán Nevado de Toluca, en la porción central de México.
La densidad promedio anual de los MIB en La Luna fue de 7 955 ± 7 349 org m-2, la densidad máxima (15 102 ± 13 789 org m-2) se encontró en mayo, mientras que la mínima (803 ± 1 013 org m-2) se presentó en enero (Fig. 5). La especie que presentó la densidad mayor anual fue L. hoffmeisteri (7 808 ± 7 270 org m-2), que es 52 veces superior a la de Pagastia sp. (146 ± 162 org m-2) (Tabla 3). La densidad mayor de L. hoffmeisteri se presentó en mayo (14 879 ± 13 776 org m-2), mientras que la densidad menor se encontró en enero (728 ± 1 071 org m-2 ) (Fig. 5).
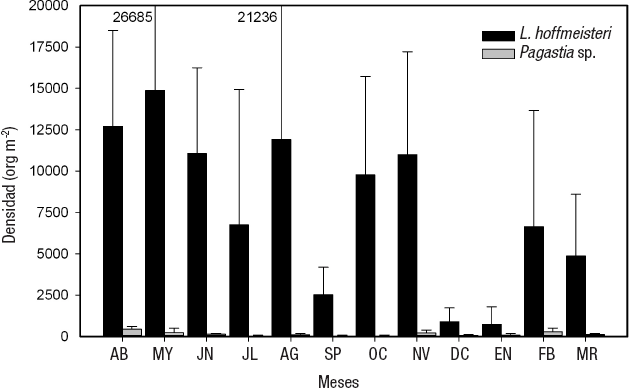
Figura 5 Variación temporal de las densidades de las especies de macroinvertebrados bentónicos de la zona profunda del lago La Luna (Luna) del volcán Nevado de Toluca, en la porción central de México. (AB=abril. MY=mayo. JN=junio. JL=julio. AG=agosto. SP=septiembre. OC=octubre. NV=noviembre. DC=diciembre. EN=enero. FB=febrero. MR=marzo.)
En la zona profunda de El Sol, las densidades anuales promedio fueron de 6541±7414 para Sol1, y 4512±8140 orgm-2 paraSol2.
En Sol1 y Sol2 la densidad menor se encontró en diciembre (921 ± 287 y 550 ± 686 org m-2, respectivamente), y su valor máximo, en noviembre (Sol1 = 13 809 ± 5 517 org m-2, Sol2 = 15 743 ± 24 org m-2). La especie que presentó la densidad mayor promedio anual en la zona profunda de El Sol fue T. Tubifex, con 4 807 ± 6 557 y 3 232 ± 8 019 org m-2 para Sol1 y Sol2, respectivamente (Tabla 2). Durante noviembre, T. tubifex obtuvo su densidad mayor (Sol1 = 12 709 ± 13 862 org m-2, Sol2 = 14 881 ± 24 811 org m-2), y en diciembre, la densidad menor (Sol1 = 133 ± 224 org m-2, Sol2 = 30 ± 51 org m-2) (Fig. 6a-b.).
Figuras 6a-b Variación temporal de la densidad de las especies de los macroinvertebrados bentónicos de la zona profunda del lago El Sol (a) Sol1. b) Sol2). Nevado de Toluca, en la porción central de México. (AB=abril. MY=mayo. JN=junio. JL=julio. AG=agosto. SP=septiembre. OC=octubre. NV=noviembre. DC=diciembre. EN=Eenero. FB=Febrero. MR=marzo.)
Para Sol1, L. variegatus fue la segunda especie con mayor densidad promedio anual (1 068 ± 1 426 org m-2), con un valor máximo en julio (3 670 ± 3 218 org m-2), y con el mínimo en enero, que no se encontraron organismos. La tercera especie en densidad fue P. casertanum (571 ± 754 org m-2), que obtuvo valores de densidad máxima en octubre (1 531 ± 1 708 org m-2), y la mínima en abril (30 ± 50 org m-2) (Tabla 2, Fig. 6a).
Para Sol2, las especies que le siguieron en densidad promedio anual a T. tubifex fueron L. variegatus y P. casertanum (587 ± 797 y 630 ± 829 org m-2) (Tabla 2). L. variegatus alcanzó la densidad mayor en junio (1 234 ± 2 022 org m-2), y la menor, en diciembre (15 ± 26 org m-2). La densidad mayor para P. casertanum se encontró en febrero (2 170 ± 1 803 org m-2), y la menor, en septiembre (59 ± 51 org m-2) (Tabla 2, Fig. 6b).
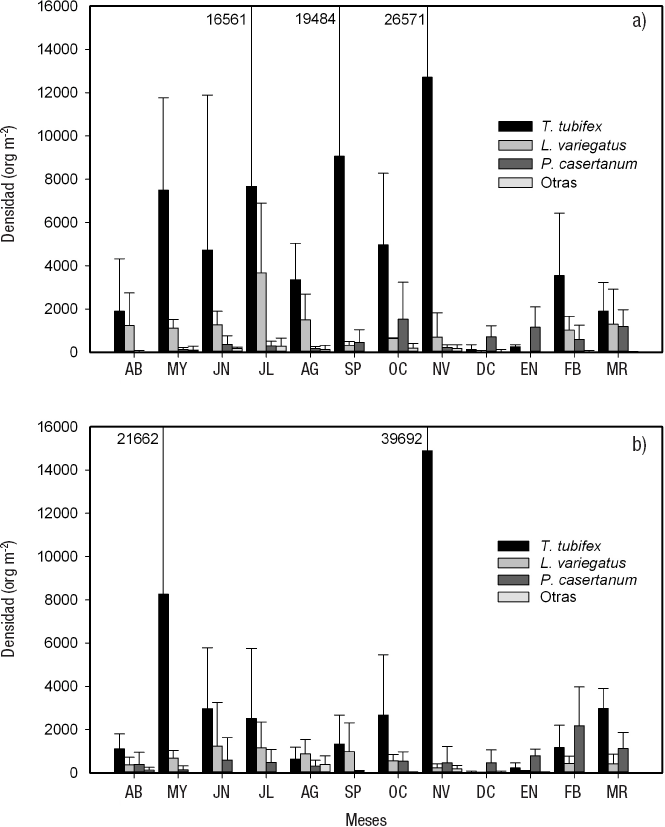
La densidad promedio anual para la zona litoral fue de 38 249 ± 33 077 org m-2 para la parte sin vegetación (Sol3), y de 25 247 ± 18 997 org m-2 para la que presentó vegetación (Sol3v) (Tabla 2). En enero se observó la densidad menor para Sol3 (4 912 ± 1 096 org m-2) y Sol3v (4 912 ± 1 608 org m-2), mientras que su densidad mayor se presentó en mayo (94 208 ± 62 954 org m-2) para Sol3, y en septiembre (49 472 ± 42 212 org m-2) para Sol3v.
En Sol3, P. casertanum presentó la densidad promedio anual mayor (12 344 ± 16 425 org m-2). Esta especie registró valores de densidad máxima en mayo, con 44 736 ± 30 453 org m-2 y mínima en octubre, con 175 ± 304 org m-2. Tanto L. hoffmeisteri como L. variegatus presentaron densidades promedio anuales similares (9 380 ± 7 573 y 9 075 ± 8 671 org m-2, respectivamente). Ambas especies tuvieron densidad menor en enero (L. hoffmeisteri 1 053 ± 526 org m-2, L. variegatus 1 403 ± 1 993 org m-2), mientras que la mayor se obtuvo en agosto (20 350 ± 11 119 org m-2) para L. hoffmeisteri, y en septiembre (22 280 ± 16 242 org m-2) para L. variegatus. La tercera especie en densidad fue T. tubifex, con un promedio anual de 6 943 ± 11 407 org m-2; su valor máximo se observó en junio (29 473 ± 22 105 org m-2) y estuvo ausente en octubre y noviembre (Fig. 7a).
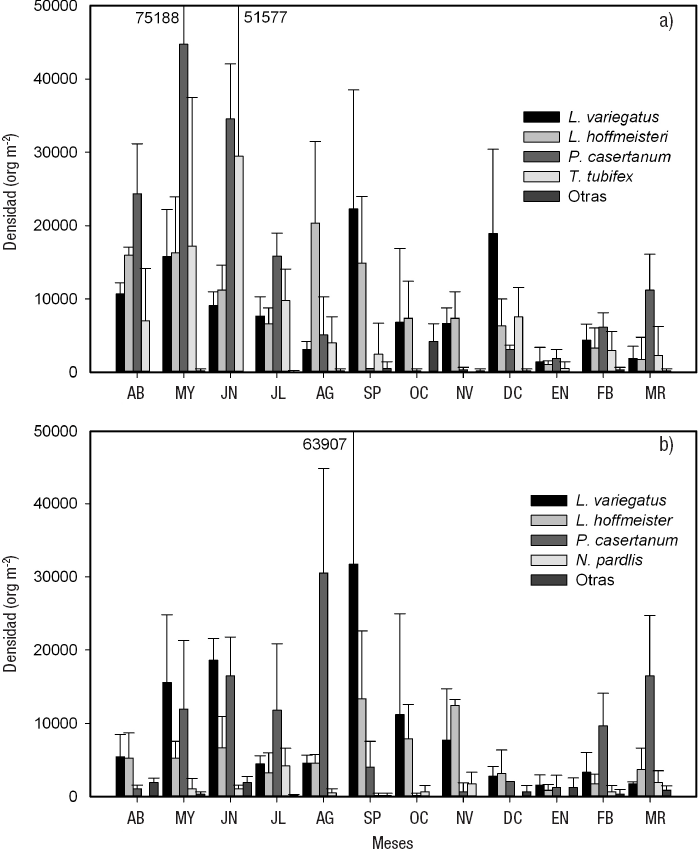
Figuras 7a-b Variación temporal de la densidad de las especies de los macroinvertebrados bentónicos de la zona somera desnuda a) Sol3, y con vegetación b) Sol3v del lago El Sol en el volcán Nevado de Toluca, en la porción central de México. (AB=abril. MY=mayo. JN=junio. JL=julio. AG=agosto. SP=septiembre. OC=octubre. NV=noviembre. DC=diciembre. EN=enero. FB=febrero. MR=marzo.)
En Sol3v, las especies que tuvieron una densidad promedio anual mayor fueron L. variegatus y P. casertanum (9 070 ± 12 478 y 8 846 ± 10 441 org m-2, respectivamente). L. variegatus presentó su densidad máxima (31 753 ± 32 153 org m-2) en septiembre, y la mínima (1 579 ± 1 392 org m-2) en enero; mientras que para P. casertanum se observó densidad mayor en agosto (30 525 ± 14 239 org m-2), y en octubre, la menor (175 ± 304 org m-2). La tercera especie en densidad promedio anual fue L. hoffmeisteri (5 382 ± 4 921 org m-2), cuyo valor máximo se encontró en septiembre, y el mínimo, en enero (13 333 ± 9 271 y 877 ± 804 org m-2, respectivamente) (Fig. 7b).
Relación entre los parámetros ambientales y los MIB. Para la zona profunda de La Luna, la primera dimensión del ACC explica el 64.8% de la suma de los eigenvalores y muestra una correlación canónica significativa (R = 0.64, p = 0.028) entre las variables biológicas y ambientales. Para el primer eje se observó que L. hoffmeisteri tuvo una relación positiva con la temperatura, y negativa con la concentración de clorofila a del sedimento (Fig. 8a). La segunda dimensión explica el 35.2% de la suma de los eigenvalores; sin embargo, no se encontró una correlación significativa (R = 0.52, p = 0.135) entre las variables biológicas y ambientales.
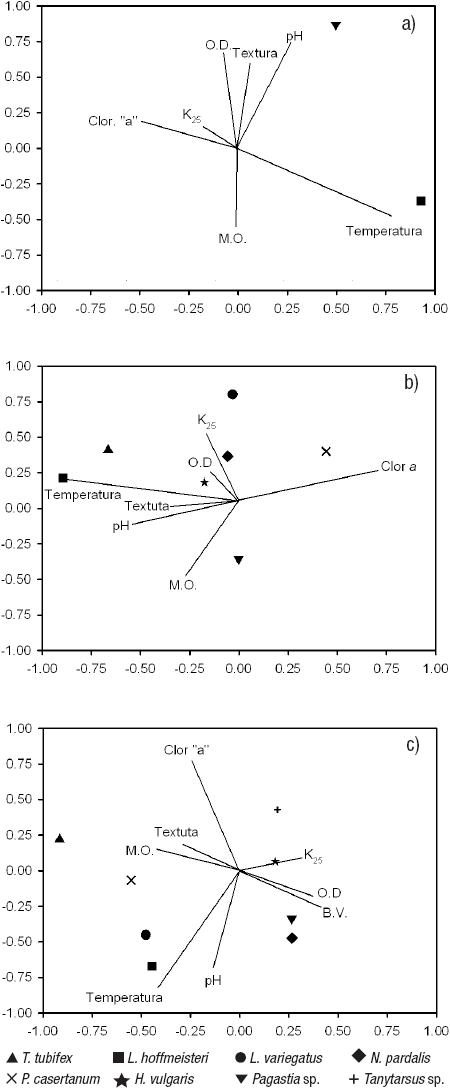
Figuras 8a-c Ordenación de las especies de macroinvertebrados bentónicos con las variables ambientales obtenido del ACC, a) La Luna. b) El Sol, Zona profunda. c) El Sol, Zona somera. Volcán Nevado de Toluca, en la porción central de México.
En la primera dimensión del ACC de la zona profunda de El Sol se explica el 45.3% de la suma de los eigenvalores y se encontró una correlación canónica significativa (R = 0.78, p = 0.000), lo que significa que las variables biológicas y las ambientales tienen una fuerte correlación. En el primer eje se observó una correlación positiva de las dos especies de oligoquetos dominantes de la zona profunda de El Sol (T. tubifex y L. variegatus) con la temperatura y el pH. Estas mismas especies tuvieron una correlación negativa con la concentración de clorofila a del sedimento. Por otra parte, P. casertanum mostró correlaciones opuestas a la de los oligoquetos para las mismas variables. En las restantes dimensiones las correlaciones canónicas no fueron significativas (p > 0.05) (Fig. 8b).
Las dos primeras dimensiones del ACC realizado para las variables ambientales y biológicas de la zona somera desnuda explican el 45.3% y 33.7% de la suma de los eigenvalores, respectivamente. Las dos primeras correlaciones canónicas fueron significativas (R1 = 0.78, p = 0.000 y R2 = 0.73, p = 0.000). Para el primer eje se encontró una correlación positiva de la temperatura, la materia orgánica y la textura con los oligoquetos dominantes de esta zona (T. tubifex, L. hoffmeisteri, L. variegatus) y con el bivalvo P. casertanum. Estas mismas especies mostraron una correlación negativa con la biomasa vegetal. En el segundo eje se observó que la temperatura y el pH tuvieron una correlación positiva con L. hoffmeisteri, L. variegatus y N. pardalis, y una negativa con Tanytarsus sp. Asimismo, la clorofila a del sedimento mostró una correlación negativa con L. hoffmeisteri, L. variegatus y N. pardalis, y positiva con Tanytarsus sp. (Fig. 8c).
Discusión
De los cuatro grupos taxonómicos encontrados en este estudio, los oligoquetos, los quironómidos y los bivalvos han sido reportados en distintos lagos de alta montaña de diferentes regiones del mundo (Bretschko, 1975; Vranovský et al., 1994; Rieradevall et al., 2000; Raddum & Fjellheim, 2002). En la mayoría de los trabajos anteriormente citados, el grupo que presenta mayor número de especies es el de los oligoquetos, seguido del de los quironómidos, tal como sucede en La Luna y El Sol. De igual forma, los bivalvos son un grupo que muestra un número reducido de especies en este tipo de ambientes.
La riqueza específica de ambos lagos muestra un número de especies por abajo del reportado para otros lagos de alta montaña, en los cuales se han reconocido entre 13 y 35 especies (Bretschko, 1975; Vranovský et al., 1994; Johnson et al., 1996; Rieradevall et al., 1999 y 2000; Raddum & Fjellheim, 2002). Esto probablemente se debe al pH ácido en ambos lagos, particularmente en La Luna, lo que ha sido observado por otros autores en lagos de alta montaña (como Schindler et al., 1985; Schindler, 1987; Raddum & Fjellheim, 2002). Larsen et al. (1996) encontraron que las especies presentes disminuyen cuando el pH es más ácido. Otra posible causa es que La Luna y El Sol se encuentran a una elevada altitud (4 200 m.s.n.m.), y la riqueza específica disminuye conforme aumenta la altitud (Dumnicka & Galas, 2002), lo cual se asocia a que los organismos están expuestos a una cantidad de radiación ultravioleta más elevada, lo que puede reducir su establecimiento y desarrollo (Kinzie III et al., 1998; Vinebrooke & Leavitt, 1999; Rhode et al., 2001).
La cantidad mayor de especies encontradas en el lago de El Sol con respecto al de La Luna se puede atribuir a la introducción de especies exóticas de peces (como Oncorhynchus mykiss Walbaum, 1792) durante el siglo pasado, tal como ha sido registrado en lagos de Canadá, Estados Unidos y Bolivia (p. ej., Modenutti et al., 1998; Matthews & Knapp, 1999; Simon & Townsend, 2003), donde se ha encontrado que al introducir truchas aparecieron especies bentónicas que no habían sido encontradas en estudios realizados anteriormente.
El número de especies en ambas estaciones de la zona profunda del lago El Sol fue similar, ya que éstas presentaron condiciones ambientales similares tanto en el agua como en el sedimento. No obstante, lo encontrado en la zona somera es contradictorio con lo que se ha reportado en otros estudios de lagos de alta montaña, donde existe un número mayor de especies en la zona con vegetación debido a que ésta presenta una gran cantidad de microhábitats y sirve como zona de alimentación y refugio contra la depredación y el arrastre por el oleaje y las corrientes (Barbieri et al., 2001; Weatherhead & James, 2001). La semejanza entre ambas estaciones litorales se puede atribuir principalmente a que en la zona desnuda tal vez no existe una fuerte presión de depredación por parte de vertebrados e invertebrados acuáticos, así como a que esta zona, que presenta forma de gancho, está protegida de la acción directa del oleaje, lo que favorece el establecimiento de más especies (Strayer & Findlay, 2010).
Tampoco se encontró diferencia significativa entre el número de especies de la zona litoral y la profunda, lo cual también ha sido observado en otros lagos con condiciones ambientales similares (Kornijów, 1988; Vranovský et al., 1994; Ivanov, 2000). No obstante, en otros lagos de alta montaña se han visto diferencias en el número de especies entre la zona profunda y la somera, lo cual se ha atribuido a que las condiciones del agua y del sedimento son diferentes en ambas zonas por presentar profundidades mayores (Rieradevall et al., 1999 y 2000).
La densidad total de los MIB tanto de La Luna como de las cuatro estaciones de El Sol fue más baja en los meses fríos (diciembre y enero); en el resto del año no se observó patrón alguno. Al analizar el comportamiento de las especies dominantes en cada estación de El Sol, se hallaron patrones diferentes entre sí.
La variación temporal de las especies de oligoquetos (L. hoffmeisteri, T. tubifex y L. variegatus) en ambos lagos se asocia principalmente a los cambios en la temperatura y la concentración de clorofila a del sedimento. En ambos lagos, las densidades menores se encontraron en los meses más fríos, y las mayores, en los de temporada cálida. Esta variación de la densidad de oligoquetos en relación con la temperatura ha sido observada por otros autores tanto en condiciones naturales como de laboratorio (p. ej., Kennedy, 1966; Lafont ,1987; Brinkhurst & Marchese, 1992; Dumnicka & Galas, 2002). Por otro lado, los valores máximos de clorofila a del sedimento se encontraron en los meses fríos para la zona profunda de La Luna y El Sol, así como para la zona somera sin vegetación. La relación inversa entre la densidad de los oligoquetos y la concentración de clorofila a sedimentaria es probablemente resultado de la disminución del consumo de células de fitoplancton por los oligoquetos durante los meses fríos, lo que permite su acumulación en el sedimento (expresada como clorofila a) (Brinkhurst, 1964 y 1975; Kennedy, 1966; Brinkhurst, 1974; Bretschko, 1975; Brinkhurst & Marchese, 1992; Juget & Lafont, 1994).
Para El Sol se obtuvo una correlación significativa en el ACC entre tres de los oligoquetos y el pH. La densidad de éstos disminuye en El Sol cuando el pH es menor a 5.3 (T. tubifex en Sol1, Sol2 y Sol3, L. hoffmeisteri en Sol3v y L. variegatus en Sol1, Sol2 y Sol3v). Tanto Vranovský et al. (1994) como Dumnicka y Galas (2002) registraron la ausencia de tubifícidos en los lagos de alta montaña de la zona del Tatra cuando el pH presenta valores por debajo de 5.2. Aunque en este caso no se encontró correlación entre el lumbricúlido y el pH, Leppänen y Kukkonen (1998) han observado que L. variegatus tiene un mejor desarrollo en un pH alrededor de 7.
Otra variable que resultó correlacionada con T. tubifex, L. hoffmeisteri y L. variegatus en la zona somera de El Sol fue la textura del sedimento, pues se encontraron las mayores densidades en los sedimentos finos. En otros estudios (Kennedy, 1965 y 1966; Lafont, 1984 y 1987; Grigelis, 1984; Brinkhurst & Marchese, 1992; Juget & Lafont, 1994), se ha observado que tanto T. tubifex como L. hoffmeisteri aumentan su densidad en diferentes tipos de lagos cuando se presenta un aumento en el porcentaje de los sedimentos finos. L. variegatus prefiere los sedimentos gruesos (arenas), donde tiene un mejor desarrollo (Grigelis, 1984; Leppänen & Kukkonen, 1998), ya que presenta densidad mayor en este tipo de textura.
El bivalvo P. casertanum tuvo sus densidades menores en diferentes meses para cada estación en el lago de El Sol; no obstante, esta especie presentó durante el muestreo una correlación negativa con la temperatura y el pH para la zona profunda y la somera de El Sol, y positiva con la temperatura para la estación sin vegetación. La correlación inversa entre la temperatura y el pH con P. casertanum en el lago de El Sol se debe principalmente a que esta especie es característica de ambientes fríos y de pH ácidos, por lo tanto, se ve favorecida bajo estas circunstancias. Es importante destacar que tanto P. casertanum como los oligoquetos dominantes se alimentan principalmente de detrito, por lo cual puede existir una competencia por el alimento entre estos dos grupos, en la cual los oligoquetos tienen mayor ventaja por su movilidad (Bretschko, 1975; Lopez & Holopainen, 1987; Brinkhurst & Marchese, 1992; Lestochava, 1994). Lo anterior se refleja cuando hay una disminución de las densidades de los oligoquetos, lo que permite la acumulación de la clorofila a en el sedimento, al disminuir y, por lo tanto, P. casertanum incrementa su densidad, esto explica la correlación positiva entre esta especie y la concentración de clorofila a del sedimento.
En la zona somera sin vegetación del lago El Sol, el comportamiento de P. casertanum con respecto a la temperatura es contrario al de las otras tres estaciones, cuyos valores mínimos de densidad coinciden con las temperaturas mínimas. Sin embargo, probablemente la relación no es directa y sean otras variables las que realmente estén provocando este comportamiento. La materia orgánica y la concentración de clorofila a sedimentarias tuvieron un descenso considerable en octubre y noviembre, por lo que la disminución del alimento conllevó un descenso de P. casertanum.
En conclusión, en los lagos de El Sol y La Luna se encontraron ocho especies de MIB, de las cuales cuatro fueron oligoquetos (Limnodrilus hoffmeisteri, Tubifex tubifex, Lumbriculus variegatus y Nais pardalis), un bivalvo (Pisidium casertanum) y un hidrozoario (Hydra vulgaris), así como dos quironómidos (Pagastia sp. y Tanytarsus sp.). Comparativamente, la riqueza específica resultó más baja, mientras que la densidad es similar a la encontrada en otros lagos de alta montaña tropicales y templados. Muy probablemente la menor riqueza taxonómica derive del efecto de las elevadas dosis de radiación UV, producto de su elevada altitud. Las especies dominantes, tanto en la zona profunda como en la somera (con y sin vegetación) de El Sol, fueron L. variegatus y P. casertanum. La especie dominante en la zona profunda y en la somera desnuda de El Sol fue T. tubifex, mientras que L. hoffmeisteri fue dominante en las estaciones de la parte somera. En La Luna, la especie dominante fue L. hoffmeisteri. La variación temporal de la densidad de los MIB presentó una correlación positiva con la temperatura y el pH. Los menores valores de densidad se presentaron en la época fría de secas, cuando la temperatura y el pH son más bajos.

