











 nueva página del texto (beta)
nueva página del texto (beta) Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroducción
La zona litoral de los lagos es un ecotono que conforma un compartimiento autónomo con todos los niveles tróficos, gran diversidad de nichos ecológicos y de redes tróficas (vía herbivoría y de detritos) y consecuentemente, mayor diversidad (Roldán & Ramírez, 2008). La interfaz tierra-agua es un hábitat dinámico que controla o influye en el intercambio de organismos, nutrientes, materia y energía dentro y a través de los ecosistemas (Larmola et al., 2006). Son zonas de alta variabilidad temporal debido a las fluctuaciones en el nivel de las aguas, la energía del oleaje, la actividad biológica y las condiciones climáticas que contribuyen a la dinámica ecológica característica de la zona litoral (Strayer & Findlay, 2010).
Debido a esta yuxtaposición de los ecosistemas terrestres y acuáticos, la zona litoral presenta un elevado contraste y heterogeneidad ambiental. Los ritmos diarios y estacionales (heterogeneidad temporal), que pulsan las actividades y estructuran el ecosistema, así como sus escalas (desde segundos hasta décadas), tienden a incrementar el número de especies y a reducir las fluctuaciones en el uso de la energía, lo que aporta cierto grado de estabilidad o constancia (Lampert & Sommer, 2007). Debido a la heterogeneidad ambiental que presenta la zona litoral, el estudio de los macroinvertebrados bentónicos (MIB) ha tomado una mayor importancia como indicadores del estado ecológico de los lagos (Shreiber & Brauns, 2010).
A pesar de lo anterior, se conoce muy poco sobre los MIB que habitan en los cuerpos acuáticos epicontinentales mexicanos, en particular sobre algunos grupos difíciles como, por ejemplo, de los oligoquetos (Peralta et al., 2002) y crustáceos (Hernández et al., 2010). La mayoría de los escasos trabajos publicados respecto a los MIB son de naturaleza estrictamente taxonómica y pocos (p.ej., Alonso-Eguía Lis et al., 2011; Rico-Sánchez et al., 2014) se enfocan en desenmascarar las variables ambientales y biológicas que influyen en su distribución y dinámica ecológica.
Como se mencionó, la zona litoral de los lagos presenta una elevada heterogeneidad ambiental lo que conlleva una diversidad de hábitats susceptibles de ser ocupados por una rica y diversa fauna bentónica. El lago Alchichica, Puebla, presenta un anillo discontinuo de tufa (i.e., estromatolitos) que corre paralelo a la costa. Esta estructura carbonatada al mismo tiempo que el descenso paulatino del nivel del lago ha propiciado que se presenten distintos grados de aislamiento o separación entre las zonas litoral y limnética (i.e., pelágica), de esta manera hay áreas que se encuentran en contacto directo mientras que otras, en el extremo opuesto, están aisladas.
Lo anterior, representa la oportunidad única de estudiar en el mismo sitio y al mismo tiempo un proceso que en la naturaleza tomaría mucho tiempo, ya que el resultado es la coexistencia de una serie de hábitats equivalentes a las distintas etapas que seguiría el proceso natural de desecación de un cuerpo acuático a lo largo del tiempo. De esta manera, la zona litoral del lago Alchichica representa un abanico de características ambientales y biológicas asociado a los distintos hábitats.
Con base en lo anterior, los objetivos del presente estudio fueron: a) caracterizar la heterogeneidad ambiental, b) reconocer las distintas comunidades de MIB presentes como reflejo de esta heterogeneidad, c) identificar las variables ambientales que mayor influencia tienen para definir a las comunidades de MIB de la zona litoral del lago Alchichica y d) describir el efecto de la desecación sobre la estructura comunitaria de los MIB.
Materiales y métodos
Área de estudio. En la porción poblana de la Cuenca Oriental (19° 24’ N, 97° 24’ 0 y 2,345 m s. n. m.) se localiza el lago maar Alchichica. El clima es templado-semiseco con lluvias en verano y temperatura media de 12.9 °C (García, 2004). La tasa de evaporación anual es de 1,690 mm y la de precipitación inferior a los 500 mm (promedio de 1966 a 2002 del Servicio Meteorológico Nacional, Adame et al., 2008).
Alchichica es uno de los lagos más hondos de México, con una profundidad máxima de 62 m y una profundidad media de 41 m, con un área superficial de 2.3 km2. La cuenca del lago alberga 94 214 080 m3 de agua salina (sólidos totales disueltos 7.2 ± 0.1 g L-1) y básica (pH 8.7 9.2), dominada por iones de sodio y cloro (Vilaclara et al., 1993; Filonov et al., 2006). Alchichica es un lago monomíctico cálido caracterizado por un periodo de circulación invernal corto (enero a marzo) y un largo periodo de estratificación (Alcocer et al., 2000).
El lago es oligotrófico, con una concentración de clorofila a anual promedio < 5 μg L-1 y concentraciones de nutrientes (nitrógeno total 4.14 ± 0.69 μM y fósforo total 0.21 ± 0.04 μM) típicamente bajas en la capa de mezcla (Oliva et al., 2001; Ramos-Higuera et al., 2008).
Muestreo. El muestreo se llevó a cabo en marzo de 2007. Las estaciones de muestreo seleccionadas se ubicaron en la periferia del lago, en la zona litoral. Como criterio de muestreo se consideró la ubicación de los sitios de colecta a distancias regulares de ≈ 500 m, lo que dio como resultado once localidades denominadas de aquí en adelante ALC-1 a ALC-11 (Fig. 1, Tabla 1). En cada localidad se midieron variables ambientales del agua y del sedimento de forma adicional al muestreo de MIB. El intervalo de profundidad en el que se tomaron las muestras fue de 5 a 80 cm.
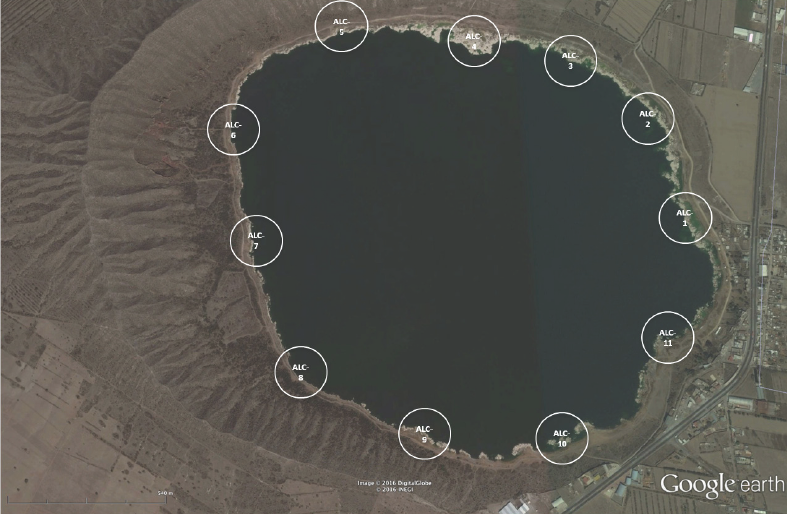
Figura 1 Ubicación de las 11 estaciones muestreadas de la zona litoral del lago Alchichica, Puebla, México.
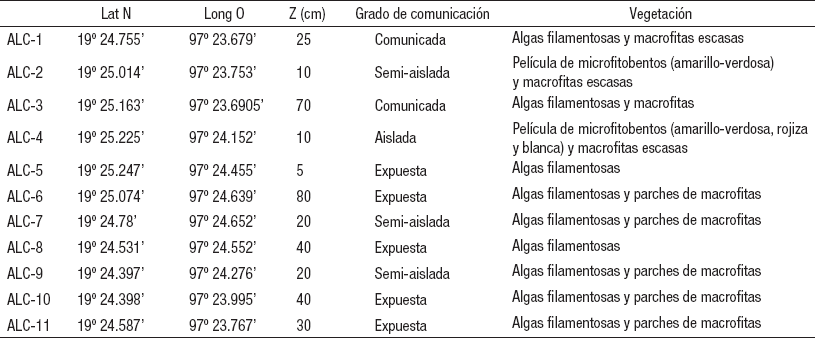
Tabla 1 Ubicación de las estaciones de muestreo en la zona litoral del lago Alchichica, Puebla, México.
Z = profundidad
Datos ambientales. En cada estación se midieron in situ factores ambientales del agua (temperatura, oxígeno disuelto, pH, potencial redox y conductividad eléctrica), mediante el uso de una sonda Hydrolab DS4. Adicionalmente, se obtuvieron muestras de agua en cada estación para analizar en el laboratorio la concentración de clorofila a (método 445.0 e la EPA, Arar & Collins, 1997), así como nutrientes (N-NH4, N-NO2, N-NO3, fósforo soluble reactivo -PSR- y sílice soluble reactivo -SiSR-), estos últimos siguiendo los métodos estándar (Strickland & Parsons, 1972) adaptados por Grasshoff et al. (1983) a los circuitos indicados por Kirkwood (1994) para un sistema autoanalizador de flujo segmentado Skalar Sanplus.
Asimismo, se tomaron muestras de 0.0005 m2 y 0.0225 m2 de sedimento para determinar parámetros ambientales (tamaño de grano, concentración de clorofila a y carbono y nitrógeno, elementales en el primer caso, y biomasa vegetal macrofítica en el segundo), las cuales se procesaron en el laboratorio. El tamaño de grano del sedimento se obtuvo por la técnica de tamizado en seco para las arenas y de pipeteo en húmedo para los limos y arcillas (Folk, 1969). Para la biomasa vegetal macrofítica, compuesta en todos los casos por Ruppia marítima Linnaeus, en cada muestra se separó, lavó y secó en un horno a una temperatura de 60°C hasta obtener peso constante; este valor se utilizó como medida de la cantidad de biomasa de vegetación presente. La concentración de clorofila a en sedimento se midió fluorométricamente siguiendo el método 445.0 de la EPA (Arar & Collins, 1997). La cantidad y calidad de la materia orgánica se obtuvo a través de los análisis de C y N elementales, que se midieron con ayuda de un analizador elemental Carlo Erba modelo NC2100.
Macroinvertebrados bentónicos. La colecta de organismos se realizó mediante una draga Ekman (0.0225 m2), con una profundidad de penetración en el sedimento de entre 5 y 10 cm. Los sedimentos colectados fueron cribados por un tamiz de 0.50 mm de apertura de malla con la finalidad de retener los MIB. Una vez separados los organismos, se fijaron y tiñeron utilizando etanol al 70% mezclado con rosa de bengala (US Geological Survey, 2012). En cada estación se tomaron tres repeticiones. Los organismos se separaron e identificaron mediante el uso de un microscopio estereoscópico hasta el nivel taxonómico más bajo posible con ayuda de claves generales y especializadas (Barnard & Barnard, 1983; Edmondson, 1959; Epler, 2001; Merrit et al., 2008; Pennak, 1978). Una vez que los organismos fueron debidamente separados e identificados, se contabilizaron.
Análisis de la información. Los valores de los parámetros ambientales fueron agrupados de acuerdo con su procedencia en: a) columna de agua (11 localidades) y b) sedimento (10 localidades, ya que la ALC-8 no presentó sedimentos); asimismo, se conjuntó la información de ambas procedencias -agua y sedimentoen un sólo conjunto general (10 localidades). Para los factores ambientales se realizó su normalización con base en su media estadística y su desviación estándar. Una vez normalizados los datos se procedió al cálculo de la distancias euclideanas con unión simple entre las estaciones de muestreo, generándose matrices de distancia que permitieron el análisis de jerarquías para producir dendrogramas, los cuales clasificaron las variables ambientales por estación de muestreo.
A partir de las mediciones ambientales normalizadas se realizaron análisis de componentes principales (ACP), utilizando los factores ambientales medidos: a) en la columna de agua de las 11 localidades, b) en el sedimento de las 10 localidades y c) en la columna de agua y sedimento de las 10 localidades.
Para el caso de la información biológica, los datos de cada muestreo fueron estandarizados dividiendo el valor de las densidades específicas en la muestra entre el total del muestreo, expresándose su valor como un porcentaje del total. Para reducir el peso y efecto de las especies dominantes en los análisis, se sustituyeron los valores de la densidad por el logaritmo natural de 1 + el valor de la densidad de cada especie [Ln (x+1)].
Una vez estandarizados y transformados los valores de densidad de los organismos se procedió a realizar una matriz de similitud de Bray-Curtis, a partir de la cual se realizó un análisis de jerarquías en unión simple generando el dendrograma. Con los valores de densidad una vez estandarizados y transformados, se realizó un ACP, utilizando todos los valores de las estaciones ALC-1 a ALC-11, excepto ALC-8 porque no presentó datos.
Por último, se realizó el cálculo de la distancias euclideanas con unión simple entre las estaciones de la zona litoral del lago Alchichica con base en variables ambientales (agua y sedimento) y biológicas (densidad). Asimismo, se efectuó el ACP correspondiente empleando tanto las variables ambientales de agua y sedimento así como las densidades de los organismos; se consideraron los componentes que explicaron el 95% de la varianza total.
Adicionalmente, se corrió un análisis de redundancia con la librería vegan 2.3 en la plataforma R Studio 0.99. Para su ejecución se utilizaron las matrices de datos obtenidas por la estandarización y transformación de la densidad de los organismos y la matriz de información ambiental (la agrupación de columna de agua y sedimento) previamente normalizada.
La categorización de las especies se llevó a cabo con base en la ampliamente utilizada escala de abundancia DAFOR (Dominante > 40%, Abundante 10-40%, Frecuente 2-10%, Ocasional 1-2%, Rara <1%). De igual manera, se catalogaron con ayuda de un diagrama bivariado de abundancia versus frecuencia, comúnmente denominado diagrama o prueba de asociación de Olmstead-Tukey (Zar, 1974; Sokal & Rohlf, 1985).
Para el análisis ecológico de la estructura comunitaria de los MIB, los datos de composición taxonómica permitieron establecer los cambios en la riqueza de especies (S). Por otro lado, con fundamento en la presencia y abundancia de las especies por localidad se calculó la diversidad (H’), la diversidad máxima (Hmax) y la equidad (J’) con el software BiodiverPro.6.
Resultados
Descripción de las estaciones. Las estaciones variaron en condiciones de exposición y aislamiento con el resto del lago y el grado de conectividad hidrológica, la cantidad de materia orgánica, presencia de estromatolitos, tipo de sustrato y vegetación (Tabla 1). La ALC-1 está conectada, con vegetación reducida y presencia abundante de algas filamentosas. La ALC-2 está semi-aislada del lago con una conexión reducida; es somera y con circulación de agua reducida; no posee macrofitas pero sí una película amarillo-verdosa de algas y bacterias. La ALC-3 es una bahía semi-cerrada pero bien conectada a través de un canal; tiene buena circulación y presencia de vegetación y algas filamentosas. La ALC-4 está aislada y no presenta conexión con el lago; es muy somera y la circulación reducida; no presenta vegetación sino una película “multicolor” (amarillo, verdosa, rojiza e incluso blanquecina) de algas y bacterias. La ALC-5 está expuesta al lago y bordeada en ambos extremos por estromatolitos; presenta escasas macrofitas y algas filamentosas. La ALC-6 está expuesta con presencia de rocas en la costa y una plataforma con gravas; hay pequeños parches de vegetación y algas filamentosas. La ALC-7 es una bahía con comunicación intermedia con el lago a través de los estromatolitos en ambos flancos; presenta vegetación y algas filamentosas. La ALC-8, como se mencionó antes, es una plataforma rocosa, de elevada pendiente y expuesta a fuertes corrientes por lo que no presenta sedimento y está totalmente desprovista de vegetación y algas. La ALC-9 presenta una conexión reducida con el lago con presencia de estromatolitos en los extremos, las corrientes son inducidas por el viento; presenta tanto vegetación como algas filamentosas en buena cantidad. La ALC-10 es una costa expuesta bordeada por vegetación y con abundantes algas filamentosas. Finalmente, la ALC-11 está expuesta y presenta “islas” de estromatolitos y talud con abundantes algas filamentosas y escasa vegetación. En resumen, de la mayor exposición/apertura a la zona limnética al mayor grado de aislamiento, las estaciones se ordenaron de la siguiente manera: ALC-8 → ALC-1 → ALC-5/ALC-6/ALC-10/ALC11 → ALC-3 ALC-7/ALC-9 → ALC-2 → ALC-4.
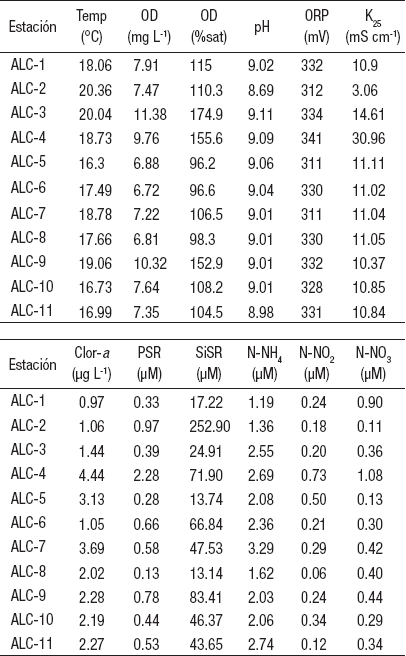
Caracterización ambiental de la zona litoral. Las variables ambientales de la columna de agua de la zona litoral (Tabla 2) mostraron baja variación en el pH (9.03 ± 0.04), potencial redox (327 ± 9 mV), temperatura (18.2 ± 1.3°C) y oxígeno disuelto (17.9 ± 1.2 mg L-1); sin embargo, difirieron en conductividad (11.31 ± 1.26 mS cm-1), concentración de clorofila a (2.12 ± 0.9 μg L-1) y concentración de nutrientes (Tabla 2). El análisis de agrupamiento de distancias euclideanas con unión simple a un nivel de corte de 4 (Fig. 2A) confirmó la gran similitud entre las estaciones en donde la que más se separa es ALC-4 debido a que presenta el agua más salada (30.96 mS cm-1), así como los valores más elevados de PSR (2.28 μM) y clorofila a (4.44 μg L-1), lo que indica su elevado grado de aislamiento y por ende los efectos de la evaporación/ concentración. ALC-4 puede ser considerada el extremo de aislamiento.
Tabla 2 Características ambientales del agua de la zona litoral del lago Alchichica, Puebla, México. (Temp = Temperatura, OD = Oxígeno disuelto, ORP = Potencial de óxido-reducción, K25 = Conductividad estandarizada a 25°C, Clor-a = Clorofila a, PSR = Fósforo soluble reactivo, SiSR = Sílice soluble reactivo, N-NH4 = Nitrógeno como amoniaco, N-NO2 = Nitrógeno como nitritos, N-NO3 = Nitrógeno como nitratos).
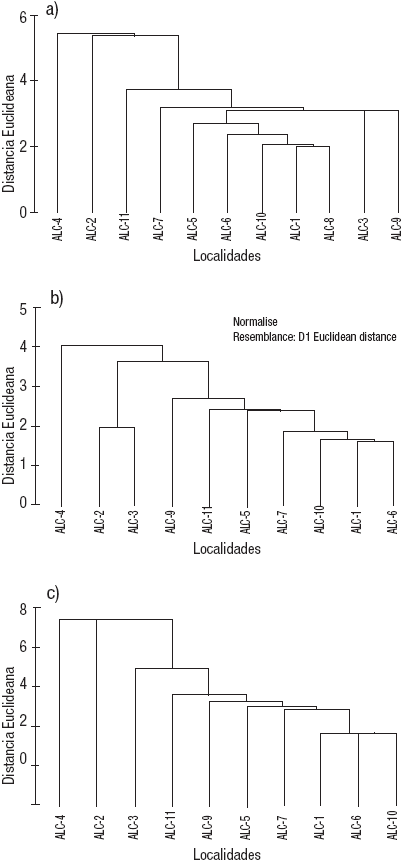
Figuras 2a-c Dendrograma de las distancias euclideanas de las estaciones de muestreo en la zona litoral del lago Alchichica, Puebla, México, con base en: a) variables del agua, b) variables del sedimento, c) combinado (agua y sedimento).
La segunda más disímil después de ALC-4 es ALC-2, ya que es la única que presenta agua dulce (3.06 mS cm-1), el menor pH (8.69) y la mayor concentración de SiSR (252.90 μM), sugiriendo la presencia de un manantial. Con base en lo anterior, ALC-2 no puede ser considerada, en sentido estricto, como parte de la secuencia de desecación de la zona litoral ya que, como se mencionó, recibe agua de un manantial.
El resto de las estaciones son hiposalinas (11.31 ± 1.26 mS cm-1), con mayor pH (9.03 ± 0.04) y menores concentraciones de SiSR (39.65 ± 24.68 μM), PSR (0.46 ± 0.20 μM) y clorofila a (2.12 ± 0.9 μg L-1).
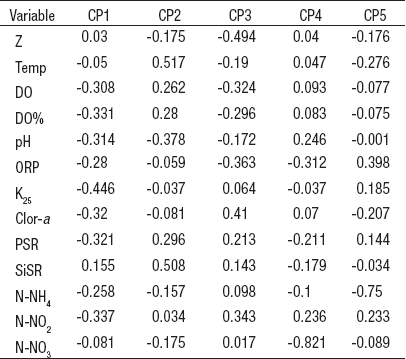
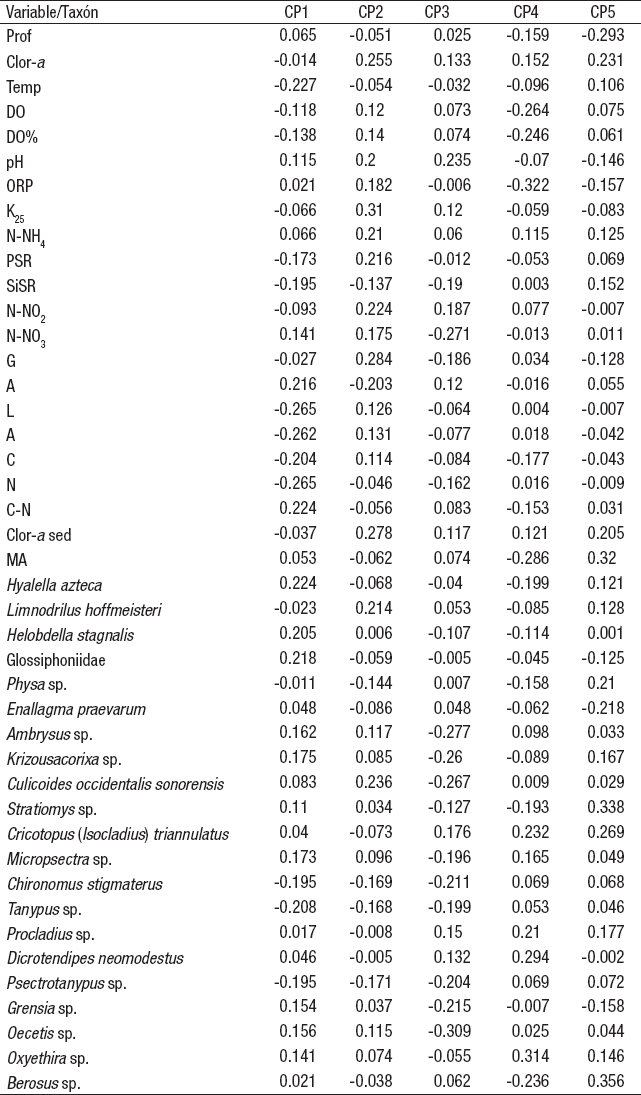
El ACP de las variables ambientales de la columna de agua de la zona litoral del lago Alchichica (Tabla 3) indicó que la conductividad es la variable de mayor relevancia (-0.446) seguida de los N-NO2 (-0.337) y del porcentaje de saturación de oxígeno disuelto (0.331) en el primer componente principal, que explica el 35.7% de la varianza total; por otro lado, la temperatura (0.517) y el SiSR (0.508) lo fueron en el segundo componente principal con el 23.0% de la varianza total.
Tabla 3 Vectores eigen del Análisis de Componentes Principales aplicado a las características ambientales del agua de la zona litoral del lago Alchichica, Puebla, México. (CP = componentes principale, Z = profundidad, Temp = temperatura, OD = oxígeno disuelto, DO% = porcentaje de saturación de OD, ORP = potencial de óxido-reducción, K25 = conductividad estandarizada a 25°C, Clor-a = clorofila a, PSR = fósforo soluble reactivo, SiSR = sílice soluble reactivo, N-NH4 = nitrógeno como amoniaco, N-NO2 = nitrógeno como nitritos, N-NO3 = nitrógeno como nitratos).
Por otra parte, las características del sedimento de la zona litoral variaron ampliamente entre las estaciones (Tabla 4). ALC-8 está constituida por una plataforma rocosa de gran pendiente que, al estar totalmente expuesta a la acción del oleaje y las corrientes, impide que se acumule sedimento; por esta razón, no se incluyó en este análisis ni en los posteriores. Por otro lado, el análisis de agrupamiento de distancias euclideanas con unión simple a un nivel de corte de 4 (Fig. 2B) mostró a la ALC-4 como la estación más diferente debido a su mayor concentración de clorofila a sedimentaria (40.42 μg cm-2), su menor concentración de biomasa vegetal macrofítica (1.3 g PS m-2) y su textura sedimentaria con un mayor contenido de gravas (19%), limos (32.8%) y arcillas (2.6%) y menor de arenas (45.7%). En contraparte, las demás estaciones mostraron menor concentración de clorofila a sedimentaria (15.7 ± 7.74 μg cm-2), su mayor concentración de biomasa vegetal macrofítica (114.0 ± 178.7 g PS m-2) y su textura sedimentaria con menor contenido de gravas (5.5 ± 4.45%), limos (10.1 ± 9.94%) y arcillas (0.8 ± 0.66%) y mayor de arenas (83.7 ± 11.43%). Se observó la separación de un grupo formado por ALC-2 y ALC-3 con base en su contenido de arenas, que posteriormente se une al resto de las estaciones para finalmente enlazarse con la ALC-4.
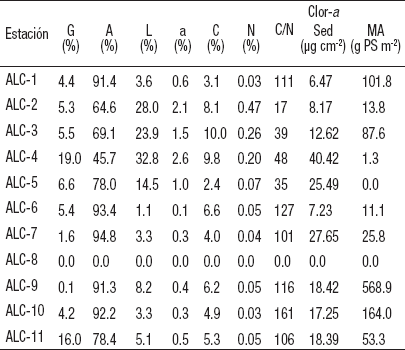
Tabla 4 Características ambientales del sedimento de la zona litoral del lago Alchichica, Puebla, México. (G = gravas, A = arenas, L = limos, a = arcillas, C = carbono, N = nitrógeno, Clor-a sed = clorofila a sedimentaria, MA = macrofitas acuáticas, PS = peso seco).
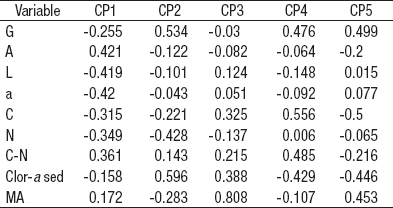
El ACP de las características del sedimento de la zona litoral del lago Alchichica (Tabla 5) develó la relevancia de la textura sedimentaria (arenas 0.421, limos -0.419 y arcillas 0.420) en el primer componente principal que explica el 59.8% de la varianza total, así como la clorofila a sedimentaria (0.596) y las gravas (0.534) en el segundo componente principal que explica el 16.8% de la varianza total.
Tabla 5 Vectores eigen del Análisis de Componentes Principales aplicado a las características ambientales del sedimento de la zona litoral del lago Alchichica, Puebla, México. (CP = componente principal, G = gravas, A = arenas, L = limos, a = arcillas, C = carbono elemental, N = nitrógeno elemental, Clor-a sed = clorofila a sedimentaria, MA = macrofitas acuáticas).
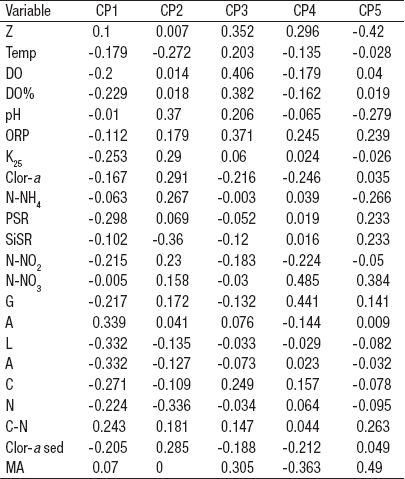
Finalmente, en el análisis de agrupamiento de distancias euclideanas con unión simple a un nivel de corte de 6 en el que se conjuntan las características tanto del agua como del sedimento, las estaciones más disímiles del conjunto son, una vez más, la ALC-4 seguida de la ALC-2 (Fig. 2C). El ACP de las características tanto del agua como del sedimento de la zona litoral del lago Alchichica (Tabla 6) mostró la relevancia de la textura sedimentaria (arenas 0.339, limos -0.332 y arcillas -0.332) en el primer componente principal que explica el 35.7% de la varianza total, además del SiSR (-0.36) y el N elemental (-0.336) en el segundo componente principal que explica el 23% de la varianza total.
Tabla 6 Vectores eigen del Análisis de Componentes Principales aplicado a las características ambientales del agua y sedimento de la zona litoral del lago Alchichica, Puebla, México. (Las abreviaturas son las mismas que para las Tablas 3 y 5).
Caracterización biológica de la zona litoral. Se colectaron un total de 22,637 organismos de MIB en las estaciones ubicadas en la zona litoral del lago Alchichica que correspondieron a 21 taxones (Tabla 7).
Tabla 7 Listado taxonómico de los macroinvertebrados bentónicos registrados en la zona litoral del lago Alchichica, Puebla, México.
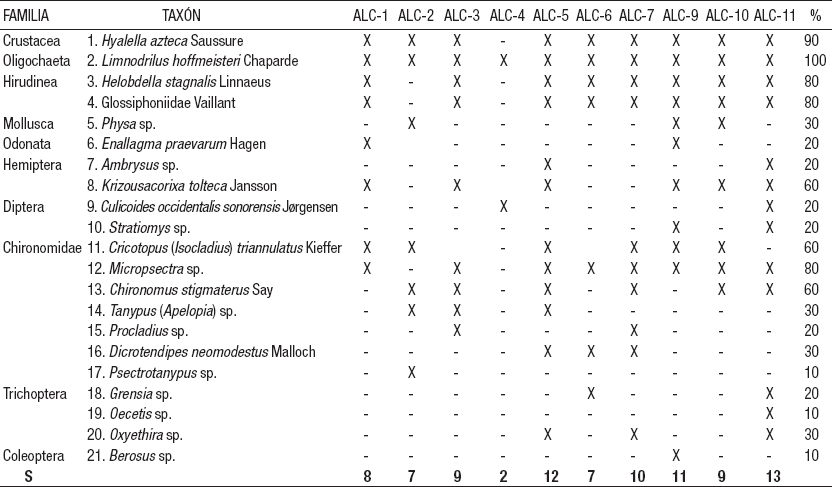
S = Riqueza taxonómica, (X) = Presencia, (-) = Ausencia, % = Frecuencia de presencia.
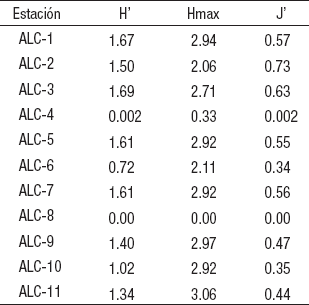
Como se mencionó, la estación ALC-8 no presentó organismos debido a la falta de sustrato adecuado, por lo anterior, de aquí en adelante no se considerará en los resultados ni en la discusión. La riqueza taxonómica varió en un rango de 2 en la ALC-4 hasta 13 en la ALC-11, con un promedio de 9 ± 3 taxones (Fig. 3A). La densidad promedio de los MIB fluctuó de 7,437 ± 5,724 org m-2 (ALC-3) a 70,252 ± 14,407 org m-2 (ALC-10), con una media global de 33,536 ± 20,463 org m-2 (Fig. 3B).
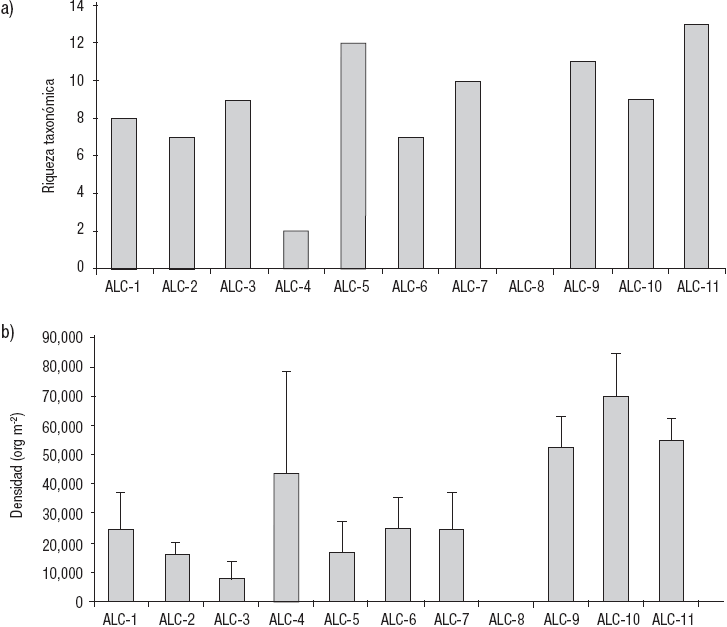
Figuras 3a-b Variación de: (a) la riqueza taxonómica y (b) la densidad promedio (± 1 desviación estándar) de las comunidades de MIB de las estaciones de muestreo en la zona litoral del lago Alchichica, Puebla, México.
La única especie que está presente en todas las localidades (100%) es Limnodrilus hoffmeisteri Claparède; con el 90% de frecuencia se encuentra Hyalella azteca Suassure, con el 80% de frecuencia se encuentran Helobdella stagnalis Linnaeus, Glossiphoniidae y Micropsectra sp. Kieffer, mientras que con el 60% están Krizousacorixa tolteca Jansson, Cricotopus (Isocladius) triannulatus Kieffer y Chironomus stigmaterus Say. El resto de los taxones presenta una frecuencia ≤ 30 %.
Con respecto a la abundancia, los principales taxones son el oligoqueto L. hoffmeisteri (45.49 %) y el anfípodo H. azteca (42.85 %) y, muy por debajo, están los quironómidos Micropsectra sp. (7.55%), Tanypus (Apelopia) sp. Meigen (2.05 %) y Ch. stigmaterus (0.51%). Los demás taxones contribuyeron con abundancias reducidas. Por su contribución en cada estación, los taxones con más abundancia son los mismos cinco, los dos primeros muy por encima de los tres últimos (Fig. 4).
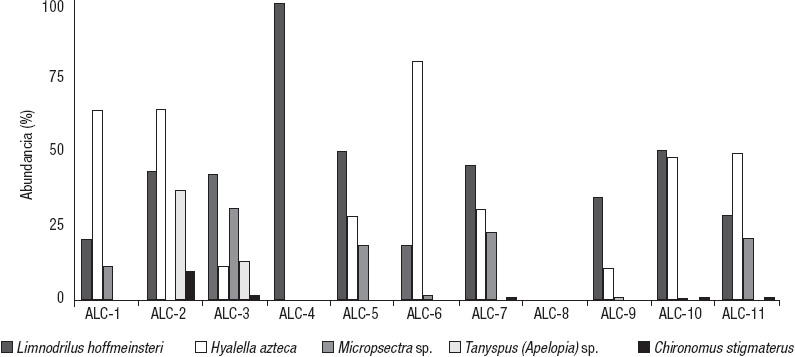
Figura 4 Contribución porcentual a la abundancia de los cinco taxones de macroinvertebrados bentónicos acuáticos más abundantes de MIB de las estaciones de la zona litoral del lago Alchichica, Puebla, México.
Las especies dominantes variaron en las distintas estaciones (Fig. 4). L. hoffmeisteri es dominante en ALC-2, ALC-3, ALC-4, ALC-5, ALC7 y ALC-10 y abundante en ALC-1, ALC-6, ALC-9 y ALC-11. Por otro lado, H. azteca es dominante en ALC-1, ALC-6, ALC-9, ALC-10 y ALC-11 y abundante en ALC-2, ALC-3, ALC-5 y ALC-7. Micropsectra es abundante en ALC-1, ALC-3, ALC-5, ALC-7 y ALC-11, mientras que Tanypus (Apelopia) en ALC-2 y ALC-3. Como frecuentes se encuentran Enallagma praevarum Hagen en ALC-1 y Ch. stigmaterus en ALC-2. Las ocasionales son H. stagnalis en ALC-3 y Glossiphoniidae en ALC-1 y ALC-5. El resto de las especies son catalogadas como componentes raros, por sus valores bajos de abundancia y frecuencia.
Con base en su abundancia y frecuencia (Tabla 8), L. hoffmeisteri y H. azteca se catalogan como dominantes (elevada -por encima de la mediaabundancia y frecuencia), Micropsectra, Ch. stigmaterus, H. stagnalis, Glossiphoniidae, K. tolteca y C. (I.) triannulatus como ocasionales (baja -por debajo de la mediaabundancia y elevada frecuencia) y el resto de los taxones son raros (baja abundancia y baja frecuencia).
Tabla 8 Clasificación de los macroinvertebrados bentónicos registrados en la zona litoral del lago Alchichica, Puebla, México.
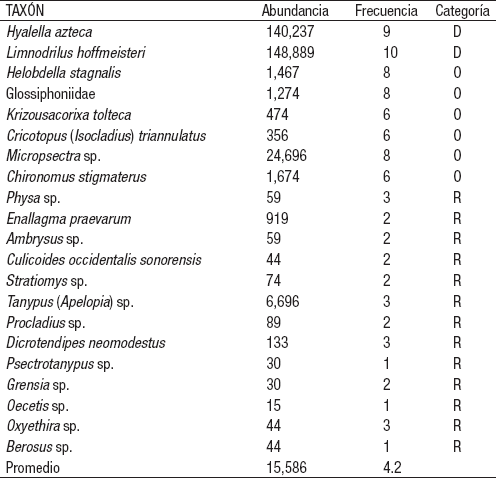
Categoría de los taxa de acuerdo a su abundancia y frecuencia. D = Dominante, = O = Ocasional, R = Rara.
Las propiedades emergentes de las comunidades de MIB que habitan en la zona litoral de Alchichica se presentan en la Tabla 9. La variación en la contribución proporcional de las especies en cada localidad, es decir la equidad, definió en gran medida la variabilidad en la estructura ecológica observada en la zona litoral del lago. La diversidad de Shannon varió de 0.002 en ALC-4 a 1.69 en ALC-3 con un promedio de 1.26 ± 0.54. Por su parte, la diversidad máxima de Shannon fluctuó en un rango de 0.33 en ALC-4 a 3.06 en ALC-11 con un promedio de 2.49 ± 0.84. Finalmente, la equitatividad de Shannon va de 0.002 en ALC-4 a 0.73 en ALC-2 con un promedio de 0.46 ± 0.20. Los valores más bajos de H’, Hmax y J’ se registraron en ALC-4 y ALC-6 al presentar riquezas taxonómicas reducidas y una contribución numérica elevada por un solo taxón. La diversidad máxima se presenta asociada a la riqueza taxonómica (13) más elevada de la estación ALC-11.
Tabla 9 Valores de los índices de diversidad y equidad de Shannon (Log Base 2) de las comunidades de MIB de la zona litoral del lago Alchichica, Puebla, México.
H’ = Diversidad, Hmax = Diversidad máxima, J’ = Equidad
Se observó una tendencia gráfica a disminuir en S, H’, Hmax y J’ de las estaciones más expuestas o comunicadas a las más aisladas (Fig. 5A). Sin embargo, este comportamiento no se apreció para la densidad (Fig. 5B). Cabe recordar que el extremo de exposición al oleaje y corrientes impide la acumulación de sedimento y con ello el desarrollo de una comunidad de MIB como aconteció en la ALC-8.
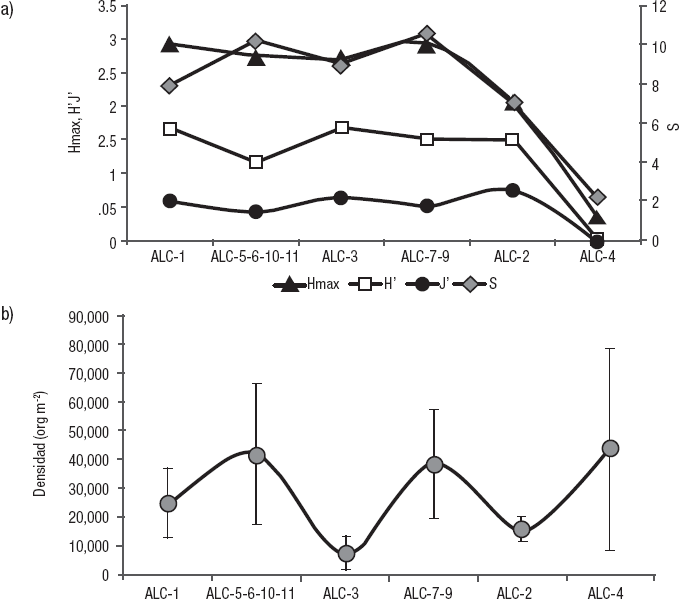
Figuras 5a-b Variación de a) Diversidad máxima (H’max), diversidad (H’), equitatividad (J’) y riqueza taxonómica (S). b) Densidad de las estaciones de la zona litoral del lago Alchichica, Puebla, México, ordenadas de acuerdo al grado de aislamiento de la zona pelágica (i.e., de ALC-1: totalmente comunicada a ALC-4: totalmente aislada).
El dendrograma del índice de similitud de Bray-Curtis aplicado a las estaciones de la zona litoral del lago Alchichica con base en la composición y densidad de las comunidades de MIB (Fig. 6A) muestra la formación de dos grupos con una similitud de aproximadamente el 80%: el primero agrupa las estaciones ALC-9, ALC-10 y ALC-6, mientras que el segundo se conforma por las estaciones ALC-5, ALC-7, ALC11 y ALC-1. Posteriormente y con un grado de similitud decreciente se enlazan a estos dos grupos ALC-3, ALC-2 y finalmente, ALC-4. Una vez más y en concordancia con las características ambientales, las estaciones más disímiles son ALC-4 y ALC-2. El ACP de la densidad de los macroinvertebrados bentónicos registrados en la zona litoral del lago Alchichica (Tabla 10) identificó que los taxones con mayor peso son Micropsectra sp. (-0.601), H. azteca (-0.525) y Tanypus sp. (0.477) en el primer componente principal que explica el 42.7% de la varianza total y los mismos taxones [Micropsectra sp. (0.735), H. azteca (-0.173) y Tanypus sp. (0.597) en el segundo componente (31%)].
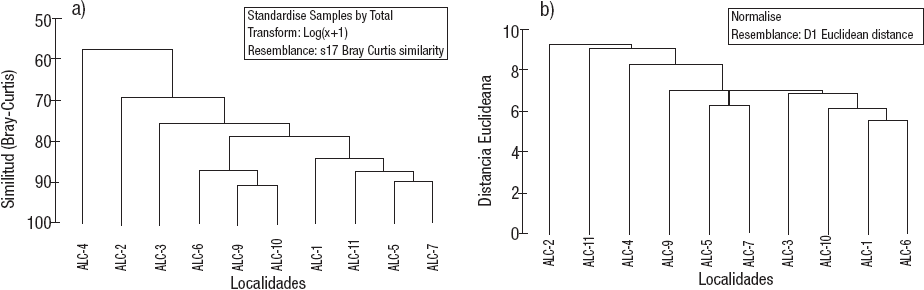
Figuras 6a-b a) Dendrograma del índice de similitud de Bray-Curtis de las estaciones de muestreo en la zona litoral del lago Alchichica con base en la densidad de organismos. b) Dendrograma de las distancias euclideanas de las estaciones de la zona litoral del lago Alchichica, Puebla, México, con base en variables ambientales (agua y sedimento) y biológicas (densidad).
Tabla 10 Vectores eigen del Análisis de Componentes Principales aplicado a la densidad de los macroinvertebrados bentónicos registrados en la zona litoral del lago Alchichica, Puebla, México.
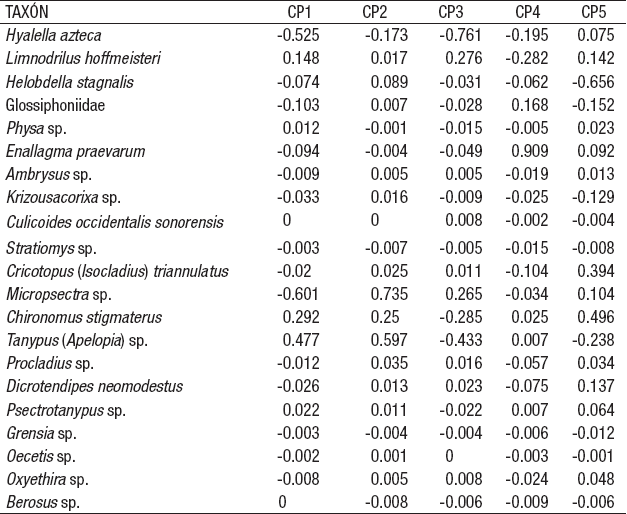
El dendrograma producto del análisis de la matriz de la variables ambientales y biológicas (Fig. 6B) mostró a ALC-2 como la estación más diferente, seguido de ALC-11 y enseguida ALC-4. El resto de las estaciones conforman un conjunto más o menos homogéneo. El ACP de las variables ambientales a las características ambientales del agua y sedimento, así como de los macroinvertebrados bentónicos registrados en la zona litoral del lago Alchichica (Tabla 11), indicó que la variable con mayor peso fue la textura sedimentaria (limos -0.265, arcillas -0.262 y arenas 0.216) y el N elemental (-0.265), de igual manera H. azteca (0.223) en el primer componente principal que explica el 26.7% de la varianza total. En el segundo componente se encontraron las arenas (-0.203), la conductividad (0.31) y la clorofila a sedimentaria (0.278) y en agua (0.255) que explica un 19.2%.
Tabla 11 Vectores eigen del Análisis de Componentes Principales aplicado a las variables ambientales a las características ambientales del agua y sedimento así como de los macroinvertebrados bentónicos registrados en la zona litoral del lago Alchichica, Puebla, México. (Las abreviaturas son las mismas que para las Tablas 3 y 5).
El análisis de redundancia efectuado con los valores de densidad de los organismos y con las variables ambientales de agua y sedimento (Fig. 7) identificó como variables ambientales de mayor peso a la textura (arenas, limos y arcillas) y la concentración de clorofila a en agua y sedimento, el N-NO2 y el PSR en agua, también el N elemental y la proporción C/N. Por la parte biológica, destacaron H. azteca, L. hoffmeisteri, Micropsectra sp., Chironomus sp. y Tanypus sp. como los taxones de mayor relevancia. H. azteca se relacionó con las arenas y la proporción C/N (las mayores densidades de H. azteca se encontraron en las estaciones con sedimentos francamente arenosos y proporciones C/N elevadas, particularmente la ALC-6), mientras que L. hoffmeisteri lo hizo con la clorofila a sedimentaria, los nitritos y el PSR (los valores más elevados de densidad de L. hoffmeisteri se encontraron en las estaciones con concentraciones más elevadas de clorofila a sedimentaria, nitritos y PSR, particularmente en la ALC-4). Por otro lado, Micropsectra sp. se asoció con ALC-7 y ALC-11 donde presentó las mayores densidades, mientras que Chironomus sp. y Tanypus sp. lo hicieron con ALC-2 y ALC-3 donde alcanzaron sus mayores densidades.
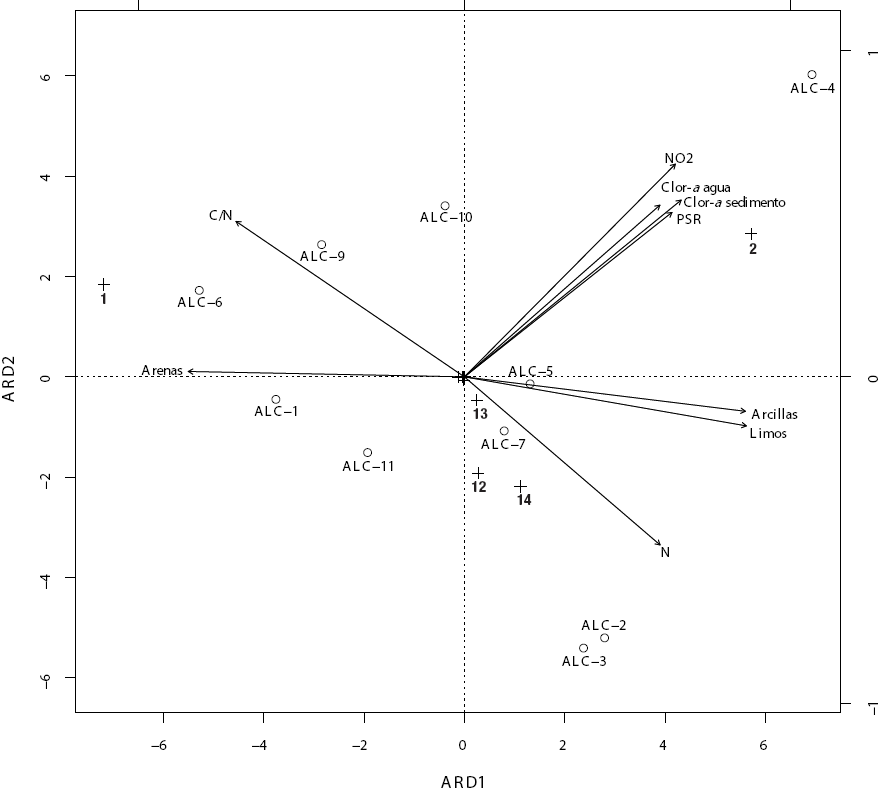
Figura 7 Triplot del análisis de redundancia (ARD) de las estaciones de la zona litoral del lago Alchichica, Puebla, México, (ALC-1 a ALC-11, excepto ALC-8) con base en las variables que resultaron significativas tanto ambientales (agua y sedimento) como biológicas (densidades de los distintos taxones). (N = nitrógeno elemental, C/N = relación carbono:nitrógeno, NO2 = nitritos, Clor-a agua = clorofila a en agua, Clor-a sed = clorofila a en sedimento, PSR = fósforo soluble reactivo). (1 = Hyalella azteca , 2 = Limnodrilus hoffmeisteri , 12 = Micropsectra sp., 13 = Chironomus stigmaterus, 14 = Tanypus (Apelopia) sp.
Discusión
Con respecto a la composición de la comunidad de MIB del lago Alchichica, existe una mayor similitud en la composición de especies entre Alchichica y los lagos salinos de Norte y Sudamérica, que con los lagos salinos de Asia y Australia que está empobrecida en insectos (Geddes et al., 1981). Los lagos Lenore, Killarney, Rabbit y Arthur en las praderas canadienses (Hammer et al., 1990) tienen salinidades y taxa similares a Alchichica como quironómidos y anfípodos. Otros lagos en Saskatchewan y Alberta, Canadá (Timms et al., 1986), también presentan un número elevado de especies en común con Alchichica. Alrededor del 50% de los taxa reportados por Galat et al. (1981) para el lago Pirámide, Nevada, están presentes en Alchichica. Lo mismo sucede en los lagos hiposalinos de Death Valley, California, estudiados por Colburn (1988) y los lagos de la región de Sud Lipez, Bolivia, identificados por Dejoux (1993).
Por otro lado, el lago Bullenmeri, Camperdown, Victoria, Australia (Timms, 1981) a pesar de que tiene una salinidad similar a la de Alchichica con 8 g L-1, su biota es muy diferente; su riqueza taxonómica es de tan sólo 12 especies y las dominantes son dos quironómidos con abundantes anfípodos pero no presenta oligoquetos. Algunos géneros presentes en los lagos del Paroo, New South Wales (Timms, 1993), también fueron registrados en Alchichica, particularmente los quironómidos. De forma similar, Williams et al. (1990) reportan el macrobentos de 79 lagos australianos de salinidades diversas; diversos géneros se encontraron en este estudio incluyendo los oligoquetos, en particular insectos. El lago Shala, Etiopía, es similar ambiental y en origen a Alchichica, sin embargo, biológicamente son diferentes en el taxón dominante, ya que en Shala son los ostrácodos, seguidos por oligoquetos y quironómidos (Tudodancea & Harrison, 1988), mientras que en Alchichica hay ostrácodos, sin embargo, son componentes principalmente de la meiofauna (Hernández et al., 2010). Los lagos salinos de Asia Central (p.ej., Balkhash en Kazakhstan, Issyk-kul en Kirghizia y Chany en Novosibirsk) tienen elevada riqueza taxonómica (Aladin & Plotnikov, 1993), pero con pocos taxa en común con Alchichica como quironómidos y oligoquetos.
De acuerdo al análisis de agrupamiento usando el índice de similitud de Bray-Curtis (Fig. 6A) de las estaciones de la zona litoral del lago Alchichica con base en la composición y densidad de las comunidades de MIB mostró que las estaciones más diferentes del resto, ALC-4 y ALC-2, coinciden en estar casi o totalmente aisladas del cuerpo principal del lago. Adicionalmente, son las estaciones con los valores extremos de conductividad, la ALC-2 con aguas dulce (K25 = 3.06 mS cm-1) mientras que la ALC-4 con las aguas más salinas (K25 = 30.96 mS cm-1); ésta última es la que presentó la menor riqueza taxonómica con tan sólo 2 taxones. La ALC-2 también muestra una baja riqueza taxonómica con 7 taxones, pero similar a otras como ALC-6 también con 7 y ALC-1 con 8. El resto de las estaciones son similares entre sí y se enlazan primero (85% de similitud), antes de hacerlo con las estaciones ALC-2 y ALC-4 con un nivel de similitud menor (75%).
El análisis de redundancia (Fig. 7) muestra que la textura sedimentaria es una variable relevante y que se asocia a la densidad de los taxones dominantes; en este caso, las arenas favorecen a H. azteca, mientras que los limos y las arcillas a L. hoffmeisteri. Se ha observado que los tubifícidos, en particular L. hoffmeisteri, aumentan su densidad cuando se presenta un aumento en el porcentaje de los sedimentos finos (Brinkhurst & Marchese, 1992, Juget & Lafont, 1994). Por otro lado, la vegetación acuática se menciona en la literatura como una variable importante para los MIB. Weatherhead & James (2001) y Urban (2004) demostraron una fuerte relación entre la vegetación macrofítica y los MIB. Cheruvelil et al. (2000) y Ohtaka et al. (2011) mencionan que la complejidad estructural proporcionada por las macrofitas favorece la densidad de MIB al aumentar las zonas de refugio contra los depredadores y al servir de alimento y de sustrato para los MIB. Schramm y Jirka (1989) mostraron la presencia de un mayor número de taxa y abundancia de invertebrados bentónicos en zonas litorales vegetadas comparadas con las desnudas; por otro lado, Machena y Kautsky (1988) mencionan que la distribución de la fauna bentónica sigue muy de cerca la de la vegetación litoral sumergida. Sin embargo, en el presente estudio no se encontró una relación clara entre la vegetación y la densidad u otros atributos ecológicos (S, H’, Hmax, J’) de los MIB; de hecho, la vegetación no apareció como atributo importante en el análisis de redundancia (Fig. 7).
De lo anterior, destaca que una característica importante que parece explicar el agrupamiento de las estaciones con base en las características ambientales y la composición y abundancia de MIB (Fig. 6B) es el grado de comunicación de la zona litoral con el cuerpo lacustre principal. En términos generales, se observa una tendencia gráfica -pero no avalada estadísticamenteque conforme disminuye la comunicación con el cuerpo principal hay una disminución de las características ecológicas (S, H’, Hmax, J’); no sucede así con la densidad en donde las mayores densidades se alternaron entre sitios más comunicados y más aislados. Lo anterior apoya que en las estaciones más comunicadas dominan sedimentos más gruesos (arenas) y con ellos la presencia de H. azteca, mientras que en las estaciones más aisladas predominan los sedimentos más finos (limos y arcillas) y con ellos la presencia de L. hoffmeisteri. Por otro lado, se observa que cuando el grado de exposición es excesivo, la energía de la corrientes y el oleaje impide la acumulación de sedimento y vegetación, lo cual evita el desarrollo de una comunidad de MIB tal y como se encontró en la estación ALC-8.
En conclusión la zona litoral del lago Alchichica estuvo caracterizada por 21 taxones de MIB entre los cuales dominaron por su abundancia y frecuencia el oligoqueto Limnodrilus hoffmeisteri y el anfípodo Hyalella azteca. Las estaciones variaron en su riqueza taxonómica (213 spp.), diversidad (0.002-1.69), diversidad máxima (0.33-3.06), equidad (0.002-0.73) y densidad (7,437-69,881 ind m-2). Se aprecia una diferencia entre las estaciones totalmente o casi aisladas del cuerpo principal del lago que muestran un mayor grado de comunicación con la zona limnética. La estructura comunitaria del ecosistema litoral se simplifica conforme se incrementa el aislamiento. Lo anterior se aprecia gráfica, mas no estadísticamente, en un decremento en la riqueza taxonómica, la diversidad, la diversidad máxima y la equidad -no así en la densidadconforme se incrementa el aislamiento. El tamaño de grano resulta ser una variable relevante para los taxones, en especial para definir la especie dominante; las arenas se vinculan a la presencia de H. azteca y su dominancia en las estaciones mejor comunicadas con la zona limnética; mientras que los limos y arcillas que predominan en las estaciones más aisladas, favorecen a L. hoffmeisteri. De esta manera la zona litoral del lago Alchichica representa un abanico de características ambientales y biológicas asociado a los distintos hábitats generados por el distinto grado de aislamiento de la zona limnética. Conforme se aísla la zona litoral, la salinidad se incrementa, los sedimentos se tornan finos -constituidos por limos y arcillas y en respuesta a lo anterior, la comunidad de MIB se va simplificando y predomina L. hoffmeisteri.

