











 nova página do texto(beta)
nova página do texto(beta) Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO 
 Permalink
PermalinkIntroducción
El cauce de los sistemas acuáticos que drenan las microcuencas boscosas, tanto de zonas templadas como tropicales, está compuesto de materia orgánica (i.e., troncos, ramas, hojas) que se origina de la vegetación ribereña (Abelho, 2001; Guevara-Cardona et al., 2006; Allan & Castillo, 2007). Este sustrato se descompone por la actividad física, química y biológica, donde los macroinvertebrados bentónicos juegan un papel importante durante la colonización y fragmentación (Graça, 2001; Callisto et al., 2007; Janke & Trivinho-Strixino, 2007), procesos que contribuyen con la dinámica de nutrientes y energía en estos sistemas dulceacuícolas (Benstead, 1996; Guevara, 2010).
A nivel mundial, estos ecosistemas son afectados por diversas actividades antropogénicas, asociadas con la deforestación y las prácticas agrícolas y ganaderas que reducen la vegetación ribereña, modifican el aporte de hojarasca y alteran la estructura y función de los macroinvertebrados bentónicos (Vörösmarty et al., 2010; Richardson et al., 2012). Los sistemas lóticos colombianos, no son ajenos a esta generalidad; pues se ha observado que los bosques ribereños, vinculados con estos ecosistemas, son modificados para el fomento de actividades de importancia económica, como la ganadería, por ejemplo, una actividad común que se asocia directamente con las cuencas hidrográficas (Márquez, 2003; Rivera-Rondón et al., 2009), y uno de los principales agentes de perturbación que afecta el recurso hídrico (Sadeghian et al., 1999; Defensoría del Pueblo Colombia, 2010), debido a que, en la mayoría de los casos, las áreas de los sistemas productivos se extienden hasta el margen de las quebradas y demás ambientes acuáticos, comprometiendo su integridad física y biológica (Chará, 2003).
El reemplazo del bosque ribereño por praderas u otros usos de suelo tiene un efecto negativo sobre la estructura y función de la fauna bentónica en ecosistemas lóticos, debido a que esta actividad puede generar una alteración significativa en las características físicas, químicas y del hábitat (Belsky et al., 1999; Sweeney & Czapka, 2004) y, a su vez, afectar el aporte y descomposición de hojarasca, considerada la mayor fuente de energía que mantiene la red trófica fluvial (Casado et al., 2015). Durante la última década, se ha considerado al proceso de descomposición de hojarasca como un método de evaluación directo de la integridad funcional de los ríos (Young et al., 2008; Riipinen et al., 2009; Feio et al., 2010), sin embargo, en el trópico, y en particular en la región Andina colombiana, esta información es aún incipiente y poco documentada.
En este sentido, la oferta hídrica en Colombia requiere de estudios que reflejen su importancia y destaquen aspectos ecológicos, como el proceso de colonización de macroinvertebrados acuáticos asociado con la descomposición de hojas, tanto de especies nativas como exóticas. Este material orgánico puede ser utilizado como un sustrato eficiente y de bajo costo, ya que una vez depositado en el curso de agua, constituye un recurso para la colonización y fragmentación por macroinvertebrados (Herranz & González del Tanago, 1986). Además, dicho sustrato, refleja cómo estos organismos utilizan el espacio multidimensional del ambiente lótico (Janke & Trivinho-Strixino, 2007).
Con base en la importancia de abordar aspectos funcionales para comprender la integridad ecológica de los sistemas acuáticos altoandinos, esta investigación tuvo como objetivo evaluar la pérdida de masa seca, las tasas de descomposición y la dinámica de colonización de macroinvertebrados bentónicos en hojas simples y mezcladas (1:1) de Eucalyptus sp. (especie introducida y ampliamente distribuida en la zona ribereña) y Tibouchina lepidota (Bonpl.) Baillon, 1877 (especie nativa dominante en la zona ribereña), en dos quebradas inmersas en matrices ganaderas de la parte alta del Río Chinchiná (Manizales, Caldas, Colombia).
Materiales y métodos
Área de estudio. Dicha área se localiza en el municipio de Manizales, departamento de Caldas, en la cuenca del Río Chinchiná, Colombia. Se seleccionaron dos quebradas, una de las cuales se definió como zona íntegra o estación de Referencia (quebrada La Elvira) y una zona visiblemente intervenida por la actividad ganadera (quebrada Chuscales). La Elvira se ubica a 2755 m.s.n.m., con las siguientes coordenadas 05°03’08.7’’N, 75°24’34’’O. Se consideró como zona de referencia debido al margen amplio de vegetación ribereña (≥ 10m) que posee y a que no presenta impacto antropogénico evidente. Chuscales se ubica a 2445 m.s.n.m., con coordenadas 05°02’45.2’’N, 75°25’18.9’’O; se encuentra visualmente impactada por actividad ganadera de tipo extensivo, con reducción promedio del ancho de la zona ribereña a ≈ 1.8 m.
Variables hidrológicas y fisicoquímicas. Se determinó la velocidad de la corriente, ancho y profundidad del cauce, según la propuesta de Chará (2003) y Sánchez (2004). Se tomaron algunas variables fisicoquímicas in situ como pH, temperatura del agua, oxígeno disuelto y porcentaje de saturación de oxígeno disuelto con un equipo multiparamétrico (LutronTM). Adicionalmente, se tomaron muestras de agua que fueron enviadas a un laboratorio certificado (ACUATEST S.A., Manizales, Colombia) para el análisis de nitrógeno amoniacal, sólidos totales y fosfatos, variables que se asocian con los efectos de la actividad ganadera sobre el recurso hídrico. Las variables hidrológicas y fisicoquímicas registradas para cada una de estas quebradas, se presentan en la Tabla 1.
Tabla 1 Variables hidrológicas y fisicoquímicas registradas en quebradas contrastantes de la cuenca del Río Chinchiná, en Manizales, Colombia, durante marzo a mayo de 2014, según el nivel de precipitación observado.

* Valores que no pudieron ser registrados por diversas restricciones logísticas.
Incubación de hojas, pérdida de masa seca y colonización por macroinvertebrados. Se eligieron dos especies vegetales dominantes en la zona de estudio, una nativa (Tibouchina lepidota (Bonpl.) Baillon conocida como Siete Cueros, con porcentaje de lignina foliar ~ entre 4.8 y 20% [para T. pulchra; Furlan et al., 2004]) y una introducida (Eucalyptus sp., NC: Eucalipto, porcentaje de lignina entre 18% y 22% [Rencoret et al., 2007]). Las hojas se recolectaron de la hojarasca acumulada en el suelo aledaño de cada quebrada. Posteriormente, se transportaron al Jardín Botánico de la Universidad de Caldas, donde fueron secadas a temperatura ambiente durante tres semanas, siguiendo la metodología propuesta por Moretti (2005). Las hojas secas se llevaron al Laboratorio de Ecología de la Universidad de Caldas, donde se pesaron 20 g de cada especie seleccionada y se realizó una mezcla de ambas hojas, con el mismo peso (1:1), con el fin de verificar si el recurso heterogéneo resultaba más atractivo para los macroinvertebrados.
El material se depositó dentro de bolsas plásticas con una dimensión de 25 x 25 cm y con un ojo de malla de 5 mm (malla gruesa) y 2 mm (malla fina). Con el material debidamente sellado, se formaron filas con cuatro bolsas cada una, distribuidas a lo ancho de cada quebrada y se incubaron durante 60 días (del 18 de marzo al 17 de mayo de 2014). Dado que los sitios de muestreo son muy cercanos al laboratorio, no se realizó el protocolo de corrección de la pérdida de masa inicial (e.g., Torres & Ramírez, 2014). En cada quebrada se colocaron 96 bolsas (32 por tratamiento/malla: Eucalipto, Siete Cueros y mixto). De acuerdo con los tiempos predeterminados (+15, 30, 45 y 60 días), se retiraron 24 muestras de cada quebrada (4 réplicas por tratamiento) en la fecha indicada; mismas que fueron puestas en bolsas con cierre hermético y transportadas al Laboratorio de Ecología, donde el contenido fue lavado y separado, por observación directa, sobre un set de tamices (500-250 μm).
El material vegetal separado fue puesto en bolsas de papel previamente rotuladas y llevadas a secado en horno a 75 oC durante 24 h, luego cada una de las muestras fue pesada en una balanza analítica con el fin de determinar la pérdida de material orgánico a través del tiempo. Los organismos separados se identificaron hasta el nivel de género y/o morfotipo bajo estereomicroscopio (10X), con base en las claves y descripciones de Domínguez y Fernández (2009), Prat et al. (2010) y Gutiérrez-López (2012). Después de la identificación, los especímenes fueron asignados a grupos funcionales de alimentación de acuerdo con la propuesta de Cummins et al. (2005), Chará-Serna et al. (2010) y las sugerencias de Ramírez y Gutiérrez-Fonseca (2014). Posteriormente, las muestras fueron incluidas en la Colección Entomológica del Programa de Biología de la Universidad de Caldas CEBUC (Registro Humboldt: No 188).
Análisis de datos. Para estimar el tiempo de degradación de las hojas (pérdida de masa seca), primero se estimó la tasa de descomposición de cada especie vegetal, la quebrada donde se recolectó y el poro de la malla, con el modelo de regresión exponencial negativo descrito por Bärlocher (2005). Las tasas (k, días-1) se calcularon de la regresión entre el ln del porcentaje de degradación de la hojarasca versus el tiempo de incubación. Para establecer diferencias significativas entre las tasas de descomposición de cada muestra, fue necesario un análisis multivariado de la covarianza (MANCOVA) usando el programa estadístico Statgraphics Centurion XVI, donde el tiempo de muestreo (días) se utilizó como covariable (Benfield, 2006). La densidad media (Ind./m2) de macroinvertebrados, que lograron colonizar las bolsas, se determinó a partir de la abundancia de éstos en cada quebrada, tratamiento y tiempos de extracción. Asimismo, para evaluar el efecto de la variación de la abundancia total de macroinvertebrados colonizadores durante cada evento de recolecta, se hizo un análisis multivariado de la varianza (MANOVA), con el programa estadístico Statgraphics Centurion XVI, previa verificación de los supuestos de normalidad y homoscedasticidad. Se utilizaron como factores: los ecosistemas fluviales (2 niveles), la hojarasca (3 niveles), el tamaño de poro (2 niveles= inclusión/exclusión) y el tiempo de muestreo (4 niveles).
Para la caracterización de las quebradas y sus posibles impactos, se calculó el índice biótico BMWP (Biological Monitoring Working Party), modificado para Colombia por Zúñiga y Cardona (2009); el índice de riqueza total EPT, según lo propuesto por Chará (2003); y se determinó la diversidad efectiva o verdadera, de acuerdo con la propuesta de Chao y Jost (2012), mediante el programa INEXT Versión 1.3.0 (Hsieh & Chao, 2013).
Resultados
Pérdida de masa seca (g) y tasa de descomposición de hojas (k, días-1). El patrón general observado entre tratamientos fue: Eucalyptus sp. > mixto > Tibouchina lepidota. La quebrada Chuscales, presentó, comparativamente, la mayor pérdida de masa seca (g) y, concomitantemente, una alta tasa de descomposición (promedio ± EE), principalmente en la malla gruesa. Sin embargo, los tratamientos con Eucalyptus sp. y el mixto, reflejaron comparativamente, los valores más altos en la malla fina de la quebrada La Elvira (malla fina, k = 0.0145 ± 0.0026; malla gruesa, k = 0.0143 ± 0.0019), y en Chuscales (fina, k = 0.0181 ± 0.0053; gruesa, k = 0.0153 ± 0.0046), respectivamente (Fig. 1).
Figuras 1a-d Patrón de descomposición (promedio ± EE) de hojas seleccionadas y valores k (días-1) asociados con dos tipos de bolsa (malla fina, 2 mm; malla gruesa, 5 mm) en quebradas contrastantes de la cuenca del Río Chinchiná, en Manizales, Colombia, durante marzo a mayo de 2014. a-b) Quebrada La Elvira. c-d) Quebrada Chuscales.
Los datos del experimento con Eucalyptus sp. en la quebrada Chuscales, primero a los 45 días (con malla fina) y después a los 60 días (con malla gruesa), no se pudieron incluir en el análisis por la pérdida de las muestras, debido a las fuertes lluvias que se presentaron durante la fase experimental. No obstante, el MANCOVA, mostró diferencias significativas entre los resultados de las muestras, al comparar el promedio de las tasas de descomposición (p< 0.05, Tabla 2). No se observaron diferencias significativas entre las quebradas ni entre el tipo de malla (p> 0.05, Tabla 2).
Tabla 2 MANCOVA aplicado sobre la pérdida de masa seca de hojas de Eucalyptus sp., Tibouchina lepidota y una mezcla de ambas especies en quebradas contrastantes de la cuenca del Río Chinchiná, en Manizales, Colombia, durante marzo a mayo de 2014.

Colonización de macroinvertebrados. Se recolectaron 4002 organismos, pertenecientes a nueve órdenes, veinticinco familias y 41 géneros y/o morfotipos (Tabla 3), donde la familia Chironomidae fue la más abundante, con un 79%, de los cuales el 86% pertenece a la subfamilia Orthocladiinae, 9% a Chironominae, 3% a Podonominae y 1% a Tanypodinae.
Tabla 3 Abundancia total de taxones de macroinvertebrados acuáticos registrados durante el proceso de colonización, presentes en los paquetes de hojas, evaluados en quebradas contrastantes de la cuenca del Río Chinchiná, en Manizales, Colombia, durante marzo a mayo de 2014.
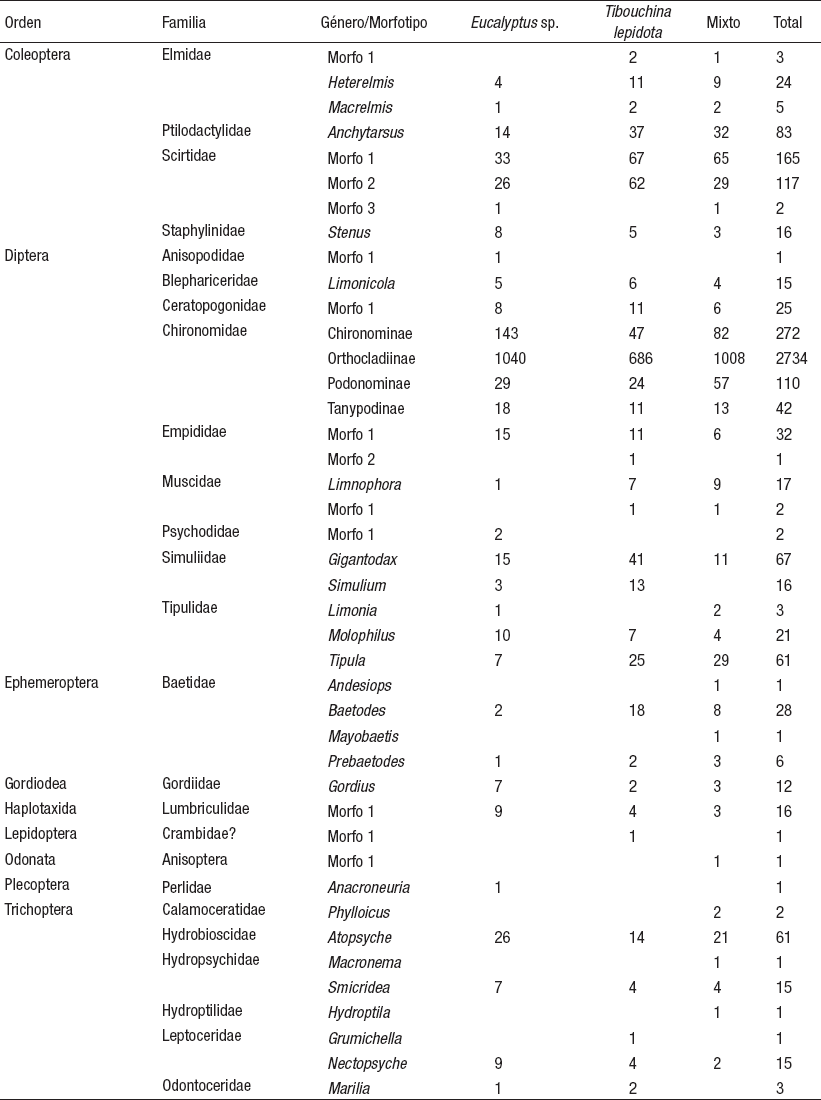
La densidad de macroinvertebrados acuáticos, a partir del número de organismos que colonizaron el área de las bolsas (25 x 25 cm = 625 cm2 = 0.0625 m2), presentó variaciones que dependieron del tipo de muestra y del tiempo que estuvieron expuestas (Fig. 2). El MANOVA, por otro lado, mostró diferencias significativas únicamente en la abundancia total de organismos y entre muestreos (p< 0.05, Tabla 4). La Elvira presentó el valor más alto de abundancia a los 30 días de muestreo, con un registro de 1129 individuos, mientras que Chuscales (ganadería) registró a los 45 días de muestreo, un total de 958 individuos.

Figura 2 Densidad media (± DE) de macroinvertebrados colonizadores de hojas seleccionadas y su combinación, durante el periodo de incubación en quebradas contrastantes de la cuenca del Río Chinchiná, en Manizales, Colombia, entre marzo y mayo de 2014. G= malla gruesa. F= malla fina.
Tabla 4 MANOVA aplicado sobre la abundancia total de macroinvertebrados acuáticos, durante el proceso de colonización sobre los paquetes de hojas en quebradas contrastantes de la cuenca del Río Chinchiná, en Manizales, Colombia, entre marzo a mayo de 2014.

Los organismos asociados con Eucalyptus sp. registraron un total de 1148; con el tratamiento Mixto, 1425 y con T. lepidota, 1129, con una mayor densidad en la quebrada La Elvira (a los 30 días, se cuantificaron 955 organismos), mientras que Chuscales registró el valor más alto a los 45 días de muestreo (852 organismos). El grupo funcional con mayor abundancia durante el periodo evaluado para las dos quebradas, fue el de los colectores, seguido de raspadores y fragmentadores (Fig. 3).

Figura 3 Abundancia total de macroinvertebrados acuáticos separados por grupos funcionales de alimentación, asociados con hojas seleccionadas en quebradas contrastantes de la cuenca del Río Chinchiná, en Manizales, Colombia, durante marzo a mayo de 2014. S.I.= sin información.
Caracterización de las quebradas y posibles impactos. Según el índice BMWP/Col, La Elvira aparece con un puntaje de 134 (aguas muy limpias); Chuscales, por otro lado, posee aguas que son sólo de buena calidad, no alteradas de manera sensible, con un puntaje de 116 (Tabla 5). Al calcular el índice EPT se encontró que en La Elvira la calidad del agua es regular, con un 26%, mientras que para Chuscales, se describe como mala, con 24%. Por su parte, la diversidad verdadera, en La Elvira, presentó mayores valores en cuanto a riqueza (específica), equitatividad (exponencial de Shannon) y dominancia (inverso de Simpson). Las variables fisicoquímicas para las dos quebradas, indicaron que el contenido de nitrógeno amoniacal y fosfatos se encuentran dentro del rango establecido por el Decreto 1594 de 1984 del Ministerio de Agricultura de Colombia, en el que se establecen los usos del agua y la delimitación de sus características para cada uso. El porcentaje de sólidos totales fue mayor en Chuscales (296 mg/L) que en La Elvira (164 mg/L) (Tabla 5).

Discusión
Pérdida de masa seca y tasa de descomposición de hojas. Los resultados del MANCOVA indicaron que las muestras, tal como se prepararon, tienen un efecto significativo sobre el porcentaje de pérdida de masa seca. El tratamiento simple con Eucalyptus sp. registró, en general, una mayor pérdida de masa y tasa de descomposición (k = 0.0162 ± 0.0024), que se puede asociar con un efecto sobre el valor k individual (sensuMoretti et al., 2007), y que puede estar vinculado con la composición química de las hojas. Wood (1974) menciona que la descomposición de las hojas de Eucalyptus ocurre como respuesta a varios factores, entre los que destacan la eliminación y/o el consumo de tejidos por invertebrados, lixiviación y/o degradación bioquímica debido a los microorganismos. Otros autores, como Hernández y Murcia (1992), señalan que los valores de descomposición se relacionan con la cantidad de nutrientes que poseen las hojas en determinado momento, ya que son responsables de la cantidad de organismos descomponedores que podrían encontrarse en ellas.
La pérdida significativamente proporcional de masa seca en el tratamiento mixto (Eucalyptus sp. + T. lepidota) durante el periodo de muestreo, puede estar ligada con la presencia y potencial influencia de los macroinvertebrados durante el proceso de degradación; probablemente la combinación de las hojas sea más atractiva para estos organismos. La alta lixiviación también es un proceso que se debe considerar, puesto que las hojas pueden llegar a perder hasta el 30% de su masa inicial (Anzar et al., 2012).
Por otra parte, Chará et al. (2007) sugieren que la textura de las hojas es otra variable a considerar en la tasa de descomposición debido a que las especies vegetales que presentan una mayor suavidad, registran valores más altos respecto a especies con una dureza superior. Esto coincide con el patrón de las especies evaluadas en la investigación: las hojas de eucalipto son suaves, comparadas con las hojas de T. lepidota, que presentan una textura rugosa. Mathuriau y Chauvet (2002), Abelho et al. (2005) y Rincón y Santelloco (2008) mencionan que la degradación de la hojarasca probablemente esté más influenciada por otros factores como la presencia de hongos y bacterias, variables explicativas en procesos de descomposición de hojarasca en el trópico y las cuales no pudieron ser evaluadas en la presente investigación.
Los resultados de este trabajo, particularmente en Chuscales, coinciden con los estudios realizados por Pettit et al. (2012) en Australia, donde el tratamiento mixto tuvo mayor pérdida de peso seco. Sin embargo, estudios realizados en España, Chile, Argentina y Colombia por diferentes investigadores (e.g., Pozo et al., 1998; Valdovinos, 2001; Albariño & Balseiro, 2002; Eyes-Escalante et al., 2012) hallaron que, al evaluar hojas nativas y exóticas, la mayor pérdida de peso seco se presentó en las nativas, sugiriendo que el reemplazo de estas especies por exóticas, puede ocasionar un fuerte impacto en el flujo de energía (Valdovinos, 2001).
Diferentes trabajos realizados con especies de Eucalyptus en Australia y Portugal (e.g., Campbell et al., 1992; Cortes et al., 1994) evidenciaron tasas de descomposición similares a las registradas en este estudio, con valores de k > 0.01. De acuerdo con Petersen y Cummins (1974), la tasa de descomposición para los tratamientos Eucalyptus sp. y mixto, en ambas quebradas y tipos de malla, se considera rápida (k > 0.01); sin embargo, para el tratamiento con T. lepidota la tasa de descomposición fue más lenta (k < 0.01), excepto en Chuscales, donde la muestra con malla gruesa fue ligeramente superior (k = -0.0102 ± 0.0038).
Colonización por macroinvertebrados. Las diferencias significativas obtenidas a través del MANOVA coinciden con lo investigado por Trama et al. (2009), que sugieren que la abundancia de macroinvertebrados acuáticos tiende a aumentar a través del tiempo y suele ser mayor en los periodos secos que en los lluviosos, similar a lo que ocurrió en esta investigación, donde las mayores densidades se registraron a los 30 y 45 días de muestreo en ambas quebradas, periodo que coincide con la época de menor precipitación (0.0 5.0 mm). A los 60 días se registró un aumento en la precipitación (20.1 60.0 mm; IDEAM, 2014) y una disminución de la abundancia, lo cual puede estar directamente relacionado con la variación del régimen hidrológico. Las precipitaciones juegan un papel fundamental como factores determinantes del ensamblaje de invertebrados bentónicos (Resh et al., 1988; Poff et al., 1997; Lytle & Poff, 2004).
Los estudios de colonización de macroinvertebrados acuáticos en paquetes de hojarasca de ríos neotropicales realizados en Colombia por Mathuriau y Chauvet (2002) y Meza y Rubio (2010); los de Carvalho y Uieda (2004), Nessimian y Henriques-Oliveira (2005), Callisto et al. (2007) y Carvalho et al. (2008) en Brasil, así como los resultados de esta investigación, indican que la familia Chironomidae es la más representativa en este tipo de experimentos, por lo que puede considerarse como eficiente a la hora de procesar las partículas de la hojarasca, gracias a sus adaptaciones morfológicas que les permite recolectar el material orgánico fino resultante de la fragmentación (Kitching, 2000; Ospina et al., 2004).
En este sentido, la mayor densidad de recolectores, correspondiente a quironómidos, sugiere que este grupo es capaz de una mayor adaptación a las diferentes variaciones hidrológicas temporales en cada estación de muestreo, en comparación con la baja densidad de otros taxones. Esto coincide con los resultados encontrados por Guerrero et al. (2004) donde los valores de densidad de macroinvertebrados obtenidos en un estudio en el río Chicamo (Murcia, España) mostraron una clara diferencia entre aquellos alcanzados por los dípteros frente a los demás taxones. Este comportamiento puede atribuirse a la dominancia de los recolectores en los ecosistemas acuáticos tropicales (Cheshire et al., 2005; Moretti et al., 2007; Rivera-Rondón et al., 2009).
Los resultados de este estudio arrojan que la densidad de este grupo funcional representó el 79% del total de los individuos recolectados y coincide con otros estudios en el Neotrópico (Moretti et al., 2007; Rodríguez-Barrios et al., 2007; Rivera-Rondón et al., 2009; Meza & Rubio, 2010), donde también la densidad de este grupo fue superior. Chará-Serna et al. (2010) y Montaño et al. (2012) sostienen que la importancia de la materia orgánica particulada (alimento de organismos recolectores) radica en ser la fuente de alimento de mayor disponibilidad para la entomofauna, asociada con ecosistemas acuáticos altoandinos. Por otra parte, la escasa existencia de recolectores/filtradores probablemente se deba a una mayor presencia de hojarasca, recurso más atractivo para los macroinvertebrados fragmentadores por el tamaño de partícula, respecto a la materia orgánica que estos pueden retener en la columna de agua (Monzón et al., 1991). Los fragmentadores, como Tipula (Diptera) y Anchytarsus (Coleoptera) y los depredadores, como Anacroneuria (Plecoptera), que alcanzan comparativamente tamaños mayores, tampoco fueron muy abundantes durante el periodo evaluado. Por el contrario, los recolectores tendieron a ser cuantiosos, pero de tamaños pequeños, tal es el caso del género Baetodes (Ephemeroptera) y varias subfamilias de Chironomidae (Diptera). Estos resultados son similares a otros estudios en las quebradas andinas colombianas, que emplearon paquetes de hojarasca (e.g., Chará et al., 2007; Meza & Rubio, 2010) y donde se argumenta la importancia de los fragmentadores y depredadores, que a pesar de ser escasos, poseen un tamaño considerable y, por lo tanto, un mayor aporte en términos de biomasa.
Caracterización de las quebradas y su posible impacto. En términos generales, Chuscales registró una escasa población de organismos indicadores de buena calidad de agua, sinónimo de una contaminación moderada en el ecosistema. Según García y Rosas (2010), la desaparición de familias intolerantes a la contaminación, indica que han ocurrido cambios ecológicos negativos en las zonas de muestreo y esta disminución de la biodiversidad puede estar directamente relacionada con actividades agrícolas, asentamientos humanos, cambios en el paisaje y alteraciones en los valores de los parámetros fisicoquímicos. Sin embargo, precisar una correlación significativa entre el impacto de la ganadería y los atributos estructurales y funcionales estuvo fuera del alcance de esta investigación.
La poca diversidad en Chuscales puede estar asociada con la pérdida del bosque ribereño, que se relaciona, a su vez, con una disminución en el aporte de materia orgánica al cauce principal, lo que tiene por consecuencia, la pérdida inevitable de los hábitats de los macroinvertebrados. Este tipo de presiones antropogénicas producen cambios que perjudican los atributos ecológicos de esta comunidad: composición, riqueza y abundancia o densidad relativa de individuos (Cao et al., 1997).
Para futuras investigaciones, se recomienda la inclusión de microorganismos (hongos y bacterias), se considere el efecto de la textura y la composición química de las hojas y, se aíslen el efecto del tamaño de la malla y el enriquecimiento alóctono de nutrientes, resultante de las actividades ganaderas, particularmente en secciones de la quebrada con reducción drástica de la vegetación ribereña.

