











 nueva página del texto (beta)
nueva página del texto (beta) Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroducción
El municipio de Zinapécuaro se caracteriza por diversas actividades económicas entre las que sobresalen la elaboración de pan, la alfarería, las prácticas agrícolas, frutícolas, madereras y, en menor escala, la ganadera (Zubieta-Rojas, 2010). Sin embargo, las actividades poco planeadas que se realizan en cuencas como la del río Zinapécuaro, han tenido como consecuencia un fuerte deterioro de los ecosistemas, la pérdida de servicios ambientales y un incremento en la vulnerabilidad de cuencas (Díaz, 2013; Mestre, 2014). Como ya reiteradamente se ha comentado para otros cuerpos de agua en México, de este río tampoco se tiene información acerca de la calidad del agua a lo largo de su recorrido y, por consiguiente, se desconoce de manera específica el tipo de contaminantes presentes.
Actualmente diferentes grupos taxonómicos son considerados como indicadores de calidad del agua (Gamboa et al., 2008; Hanson et al., 2010), en particular, las diatomeas, que han sido empleadas para valorar las condiciones ecológicas cambiantes de los sistemas lóticos (Tang et al., 2006; Salomoni et al., 2006; Zampella, 2007), puesto que sus poblaciones responden a las etapas de sucesión de la vegetación original, a diferentes usos del suelo, así como a las actividades industriales, agrícolas y urbanas que incrementan el uso de nutrientes y sedimentos en las cuencas, situación que genera alteración en las condiciones ambientales de los ríos con efectos negativos en sus componentes bióticos (Ndiritu et al., 2006). Los muestreos son fáciles de realizar y sus requerimientos ecológicos son mejor comprendidos que los de otros componentes de la biota acuática (Martínez de Fabricius et al., 2003; Díaz & Maidana, 2005; Walker & Pan, 2006; Weilhoefer & Pan, 2006; López & Siqueiros, 2011).
En este trabajo se analizó la riqueza, composición, abundancia y distribución de la comunidad de diatomeas epilíticas en cuatro temporadas de muestreo y su relación con algunas variables ambientales en la cabecera del río Zinapécuaro.
Materiales y métodos
La cuenca Alta del río Zinapécuaro tiene una altitud que oscila entre 2,512 y 2,433 m.s.n.m., con coordenadas 19° 50' - 19° 51' N; 100° 43' - 100° 46' O. Su clima es templado con lluvias en verano, con una precipitación promedio anual de 622.5 mm y temperaturas que fluctúan entre 3 y 34 °C (Correa, 2003). La vegetación primaria es bosque de oyamel, bosque de pino y bosque mixto de pino y encino (Correa, 2003), ecosistemas que se están erradicando para dar paso al establecimiento de huertas de aguacate.
Se seleccionaron cinco sitios de muestreo, considerando las diferentes actividades humanas que se desarrollan en la región, y en función del uso comunitario que se hace del recurso agua. Dos son afluentes del río Zinapécuaro, el manantial de La Yerbabuena (Yman), que proporciona agua potable a la ranchería, se usa como potrero y tiene sembradíos en los alrededores; y el río Yerbabuena (Yrío), empleado como abrevadero, además de verter las aguas residuales del poblado y de un aserradero. El tercer sitio es un aljibe recolector de manantiales que surte de agua potable al poblado La Yerbabuena (Elaj). Tanto el poblado La Galera (Lgal) como el balneario Las Orquídeas (Lorq) forman parte del cauce principal de este cuerpo de agua; el primero se utiliza como abrevadero y desagüe, en tanto que el segundo atrae al turismo, ya que se encuentra en una zona de bosque de pino con numerosos manantiales que se incorporan al río y tiene instalado un banco de materiales que vierte desechos de caolín directamente a la corriente (Tabla 1, Fig. 1).
Tabla 1: Localización de los sitios de estudio en la cabecera del río Zinapécuaro con anotación de actividades humanas realizadas en la zona.

*Nota: Af-afluente; P-potrero; S-sembradíos; Eud-extracción de agua para uso doméstico; Ab-abrevadero para aves de corral y ganado; As-aserradero; Ar-vertido de aguas residuales; Ap-almacenamiento de agua potable; Ha-huertas de aguacate; Tl-turismo local; Bm-banco de materiales.

Figura 1: Localización de los sitios de estudio en la cabecera del río Zinapécuaro, Michoacán, México. 1) Manantial La Yerbabuena (Yman). 2) El río Yerbabuena (Yrío). 3) El Aljibe (Elaj). 4) La Galera (Lgal). 5) Las Orquídeas (Lorq).
Se efectuaron cuatro recolectas, una en poslluvias en noviembre de 2012 y tres en 2013: estiaje (abril), lluvias (junio) y poslluvias (noviembre). In situ, se midieron el oxígeno disuelto, nitritos, nitrógeno amoniacal, alcalinidad total y dureza total con un equipo HACH FF-1A. La profundidad se registró con una sonda graduada. Con un potenciómetro Conductronic PC-18 se determinaron los parámetros de temperatura, pH y sólidos disueltos totales (SDT) (Tabla 2).
Tabla 2: Variables fisicoquímicas de los sitios de muestreo en la cabecera del río Zinapécuaro en el periodo 2012 y 2013.

Z=Profundidad. T=Temperatura. O.D.=Oxígeno Disuelto. SDT=Solidos disueltos totales. N-NO2=Nitritos. N-NH4=Amonio. Alc= Alcalinidad a la fenoftaleína. Dt.=Dureza total. T=Flujo turbulento. L=Flujo laminar. s/f=Sin flujo.
Las muestras de diatomes epilíticas se tomaron de seis rocas seleccionadas de forma arbitraria, en zonas con flujo rápido, para cada sitio y temporada. Se empleó el método de Iserentant et al., 1999, que consiste en cepillar la superficie superior de las rocas (aproximadamente 5 cm2) y fijar las muestras en formol al 4%. Las valvas de las diatomeas se limpiaron con peróxido de hidrógeno (40%) para eliminar la materia orgánica y con ácido clorhídrico (37%) para disolver el carbonato de calcio. Se hicieron dos preparaciones permanentes por sitio y temporada mediante el montaje de los frústulos limpios en resina sintética (Merck IR=1.5). Las valvas se identificaron con auxilio de un microscopio óptico Axiostar plus ®bajo una magnificación de 100x, según los criterios de Krammer, 1997; Krammer y Lange-Bertalot, 1997a, b; Krammer y Lange-Bertalot, 2004a, b; Lange-Bertalot, 2001; Metzeltin y Lange-Bertalot, 1998, 2002; Metzeltin et al., 2005.
A partir de las muestras ya limpias se tomaron submuestras de 200 µl con las cuales se contabilizaron 400 valvas por sitio y por temporada (Kelly et al., 1998) y se obtuvo la abundancia relativa de los taxones. Con los datos de abundancia absoluta se estimó la riqueza (S); la diversidad se calculó con el índice de Shannon-Wiener; la similitud entre sitios y temporadas se evaluó a través de un análisis jerárquico de agrupación con el método de liga promedio y el índice de similitud de Bray-Curtis. Los valores de diversidad obtenidos con el índice Shannon-Wiener se compararon con la prueba de t modificada por Hutcheson (Zar, 2010). La riqueza esperada se estimó empleando el modelo no paramétrico de acumulación de especies Chao 2 (Magurran, 2004) y los estimadores paramétricos de Jacknife 1 (JK 1) y Bootstrap (Boot). De igual forma, se aplicó un Análisis de Varianza de Similitudes (ANOSIM) para determinar diferencias estadísticamente significativas en cuanto a la composición de las comunidades registradas por sitio y temporada, además de un Análisis de Correspondencia Canónica (CANOCO) para identificar cuáles de las variables fisicoquímicas medidas tuvieron mayor efecto sobre la distribución de las especies. La rarefacción se efectuó para evaluar la relación entre el esfuerzo de muestreo y el número de especies encontrado, así como para clarificar el efecto de las condiciones de los sitios y temporadas independientes al tamaño de muestra, para los que se emplearon las abundancias absolutas registradas por sitio y temporada. Los análisis se llevaron a cabo con los programas BioDiversity (McAleece et al., 1997) y PAST v. 2.17 (Hammer et al., 2001).
Resultados
Caracterización ambiental. Con base en las características fisicoquímicas, el río Zinapécuaro se puede considerar como un cuerpo de agua subtropical (14.5 - 22.3 °C). Presenta aguas de circumneutrales a alcalinas (pH 7.0 - 8.2); con una concentración de sólidos disueltos que se vio incrementada en La Galera durante el periodo de estiaje (767 ppm) y lluvias (623 ppm) de 2013. Las concentraciones de oxígeno disuelto tuvieron condiciones de mediana oxigenación (4.0 mg/L) en el estiaje y hasta muy elevada en la época de lluvias (10.0 mg/L). En relación con la profundidad, el valor mínimo fue para poslluvias (2012) (5 cm), salvo en El Aljibe que prácticamente estaba vacío y sólo se presentaron pequeñas charcas. El amonio tuvo la concentración máxima en poslluvias (2013) (0.9 mg/L) y los nitritos aumentaron sus valores en lluvias y poslluvias (2013) (0.1 y 0.2 mg/L respectivamente). En cuanto a la alcalinidad el valor máximo se presentó en lluvias (2013) (153.9 mg/L); la dureza total compartió el valor máximo tanto en poslluvias (2012) como en lluvias (2013) (153 mg/L) (Tabla 2).
Riqueza por sitio y temporada. Como resultado del muestreo en los cinco sitios y las cuatro recolectas se registraron 70 especies, seis variedades y una forma de diatomeas epilíticas. Estos taxones representan una familia, dos géneros y dos especies de la clase Coscinodiscophyceae; una familia, tres géneros, seis especies y dos variedades de la clase Fragilariophyceae y 15 familias, 25 géneros, 62 especies, cuatro variedades y una forma correspondieron a la clase Bacillariophyceae. En la Tabla 3 se enlistan las especies y sus autoridades por clase taxonómica y orden alfabético.
Tabla 3: Listado taxonómico por clases y orden alfabético, de las diatomeas epilíticas recolectadas en la cabecera del río Zinapécuaro.

Las especies que se encontraron en todos los sitios de muestreo y en las cuatro temporadas fueron: Achnanthidium exiguum, A. minutissimum, Encyonema silesiacum, Eolimna subminuscula, Frustulia latita, F. neomundana, Gomphonema lagenula, Navicula rhynchocephala, Nitzschia clausii, N. palea, Planothidium frequentissimum, P. lanceolatum y Rhopalodia brebisonii.
Entre las especies que tuvieron baja abundancia relativa en un sitio y en una temporada se encontraron Amphora ovalis, Caloneis spec. cf. bacillum, Cocconeis pseudothumensis, Diploneis pseudovalis, Eunotia bilunaris, E. cataractarum Fragilaria capuccina aff. gracilis, Frustulia krammeri, Gomphonema clavatum, Navicula clementis, Nitzschia umbonata, Pinnularia gibba, Placoneis undulata, Stephanodiscus niagarae y Ulnaria goulardii. En tanto, aquéllas con baja abundancia en un sitio fueron Achnanthes inflata, Eunotia formica, E. quaternaria y Fragilaria capucina var. vaucheriae (Tabla 4).
Tabla 4: Abundancia relativa de las diatomeas epilíticas en la cabecera del río Zinapécuaro durante el periodo de estudio (2012-2013).

Como se puede apreciar en la Tabla 5, las estimaciones de riqueza obtenidas con los diferentes modelos utilizados, indicaron que para toda el área y las temporadas muestreadas se esperaría registrar entre 78 y 87 especies, por lo que el estudio conjunta entre el 80.5% y el 89.7% de lo estimado. Por temporadas, se aprecia que las 55 especies registradas durante el estiaje representaron la época más rica; pero también fue la que menor representación de riqueza obtuvo con un máximo de 83.3 % de la riqueza específica esperada, de acuerdo con los diferentes estimadores utilizados. En la temporada de lluvias se registró entre el 79% y el 90.7% de la riqueza esperada, que constituyen los valores más altos y menos variables de representación de la riqueza esperada (Tabla 5).
Tabla 5: Estimaciones de riqueza específica y proporción representada por la riqueza observada por temporada de muestreo. La proporción que representa la riqueza registrada con respecto al estimado se muestra en porcentaje dentro del paréntesis.

En general, las diferencias entre las estimaciones de riqueza y la riqueza registrada fueron menores del 5% con Bootstrap, en la época de poslluvias (2012) a 40% aproximadamente en el estiaje (2013), de acuerdo con el estimador Chao 2.
La Galera fue el sitio con menor riqueza y también el mejor representado en riqueza con 91.7 al 100% de lo esperado; mientras que los sitios del manantial y río La Yerbabuena comprendieron registros de 75 a 86% de la riqueza esperada (Tabla 6).
Tabla 6: Estimaciones de riqueza por sitio muestreado. Entre paréntesis se muestra el porcentaje que representa la riqueza observada con respecto a la estimación.

* En El Aljibe sólo se tuvo un muestreo por lo que no es posible estimar la riqueza esperada
La técnica de rarefacción permitió apreciar que en general el esfuerzo de muestreo utilizado fue el adecuado para recoger la riqueza específica de diatomeas epilíticas en los sitios y temporadas estudiados, excepto en el manantial y el río Yerbabuena durante la época de lluvias (2013) y el manantial de la Yerbabuena en el estiaje (2013), cuyas curvas mostraron aún tendencia creciente, lo que sugirió que se requeriría un número mayor de submuestras para alcanzar una buena representación de su riqueza (Figuras 2, 3, 4 y 5). También se puede apreciar que las diferencias de riqueza encontradas entre los sitios no es un efecto del número de individuos capturados, sino de las características y condiciones de los sitios, los cuales presentan diferencias importantes en los parámetros fisicoquímicos medidos.

Figura 2: Curva de rarefacción para diatomeas de la cabecera del río Zinapécuaro durante la temporada de poslluvias (2012).

Figura 3: Curva de rarefacción para diatomeas de la cabecera del río Zinapécuaro durante la temporada de estiaje (2013).

Figura 4: Curva de rarefacción para diatomeas de la cabecera del río Zinapécuaro durante la temporada de lluvias (2013).

Figura 5: Curva de rarefacción para diatomeas de la cabecera del río Zinapécuaro durante la temporada de poslluvias (2013).
Diversidad por sitios/temporadas. La Tabla 7 muestra los valores de diversidad medida con el índice de Shannon (H’) para cada sitio y temporada. El valor más alto, según la temporada, se presentó en época de lluvias (2013) en Las Orquídeas (2.724) sin diferencia significativa con El Aljibe en la época de estiaje, lo mismo ocurrió en ambos sitios durante poslluvias en 2013. Los valores más bajos para las cuatro épocas muestreadas se presentaron en La Galera, esto sugiere un efecto importante de las condiciones de este sitio en la diversidad de diatomeas. La misma situación se apreció con los cinco valores más elevados de diversidad, los cuales correspondieron a tres temporadas diferentes y tan sólo a tres sitios: Las Orquídeas (contó con tres de estos valores), El aljibe y el río Yerbabuena.
Tabla 7: Diversidad estimada con el Índice de Shannon-Wiener por sitios/ temporadas. En la columna de la derecha se indican las diferencias estadísticas entre sitios/temporadas. Letras diferentes representan valores de diversidad estadísticamente diferentes (p < 0.05). Valores marcados en negritas representan el efecto de las condiciones de los sitios en la diversidad de diatomeas.

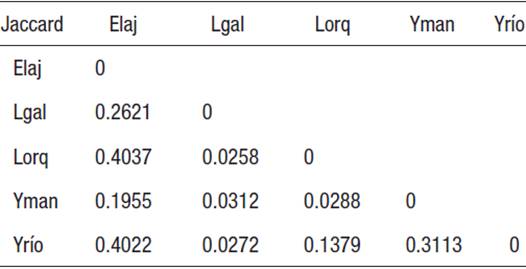
Similitud entre sitios y temporadas. La prueba estadística ANOSIM de una vía para temporadas, no mostró diferencia estadística significativa en similitud, producto de la época de muestreo (p>0.05); mientras que para evaluar el efecto de los sitios, el análisis con el índice binario de semejanza de Jaccard, mostró diferencia de riqueza entre La Galera con respecto a tres de las otras localidades, excepto con El Aljibe. Al aplicar el análisis cuantitativo con el índice de Bray-Curtis, se recuperaron las diferencias de La Galera anteriormente mencionadas, además de la diferencia entre el manantial de La Yerbabuena y Las Orquídeas (Tabla 8).
Tabla 8: Resultados de ANOSIM de una vía para la comparación de sitios/ temporadas. Áreas sombreadas representan diferencias significativas (p<0.05).
La Figura 6 muestra un gráfico de agrupación con el índice de Bray-Curtis en el que se aprecia un grupo formado por tres de los cuatro muestreos del sitio La Galera, que refleja las diferencias mencionadas.

Figura 6: Agrupación de los sitios de muestreo por sitios/temporadas obtenido con el Índice de Bray-Curtis, método de liga promedio. La flecha indica el grupo formado por tres de las recolectas en La Galera.
El Análisis de Correspondencia Canónico mostró más de 70% de la variación explicada en los dos primeros ejes de ordenación, los valores de correlación canónica obtenidos (Tabla 9) indicaron que la distribución de las especies estudiadas varió de acuerdo con la profundidad, nitritos, amonio, pH y dureza total.
Tabla 9: Valores de correlación canónica obtenidos para los diferentes parámetros fisicoquímicos medidos en los diferentes sitios y épocas. El eje 1 está relacionado positivamente con la profundidad y negativamente con el pH y la dureza total. El eje 2 está correlacionado positivamente con los nitritos y el amonio.

El ACC permitió separar tres grupos principales de especies como se observa en la Figura 7. El primer grupo de especies, formado por Navicula antonii, N. germainii, N. recens, Nitzschia filiformis, N. inconspicua, N. gracilis, N. palea, N. umbonata y Stephanocyclus meneghiniana, se encuentra en sitios con menor profundidad, elevados valores de dureza total y medios alcalinos. En el lado superior derecho del gráfico se aprecia otro grupo de especies, entre las que se ubican a Craticula accomoda, Diploneis pseudovalis, Encyonema silesiacum, Eunotia quaternaria, E. veneris, Fragilaria capuccina var. vaucheriae, Frustulia latita, Gomphonema pseudoaugur, Lemnicola hungarica, Navicula cryptocephala, Rhoicosphenia abbreviata y Rhopalodia brebissonii, por mencionar algunas y que se le asocia a zonas de mayor profundidad, mayores concentraciones de nitritos y amonio y bajas concentraciones de dureza total y medios circumneutrales. Un tercer grupo de especies, integrado por Cocconeis pseudothumensis, Cymbella naviculiformis, Eolimna subminuscula, Eunotia cataractarum, E. bilunaris, Frustulia krammeri, Geissleria ignota, Luticola mutica, Navicula wildii, Planothidium frequentissimum, entre otras, se vincula con elevados valores de profundidad, bajas concentraciones de nitritos y amonio. El resto de las especies se encuentran en valores intermedios de concentraciones de nitritos y amonio, en diferentes profundidades, medios circumneutrales y bajas concentraciones de dureza total.

Figura 7: Ordenamiento obtenido con el Análisis de Correspondencia Canónico (ACC) que muestra la distribución de las especies en función de los sitios/temporadas y los parámetros fisicoquímicos medidos. Variación explicada en los dos primeros ejes de ordenación > 70%. Las flechas indican la dirección de la variación (menor a mayor valor medido) de los parámetros más importantes en los dos ejes principales. Acrónimos de las especies como en la Tabla 2.
Discusión
Se ha considerado que la distribución de las comunidades de diatomeas bentónicas es el resultado de las interacciones espaciales y temporales de los factores fisicoquímicos, más que de los factores geológicos o climáticos (Potapova & Charles, 2002); por ello, la química del agua ha sido considerada como el elemento crucial para la composición de las especies (Soininen et al. 2004).
En el caso de la cabecera del río Zinapécuaro se observaron cambios en los valores de los parámetros fisicoquímicos así como variaciones de la comunidad de diatomeas en las temporadas de estudio. Los sitios manifestaron poca contaminación y una incipiente perturbación, resultado del manejo antrópico que se da al río en esta zona (Tabla 1). Esto explica la presencia de especies como Eolimna subminuscula, Encyonema silesiacum, Gomphonema lagenula, Nitzschia clausii, N. palea, o Navicula veneta, que por sus afinidades ecológicas indican cierto grado de eutrofización (Krammer & Lange Bertalot, 1997a, b; Krammer & Lange Bertalot, 2004a, b; Segura-García, 2011) (Tabla 4).
Especies como Achnanthidium minutissimum, Planothidium frequentissimum y Rhoicosphenia abbreviata, sensibles a la contaminación y reportadas para aguas oligotróficas a mesotróficas (Cox, 1996; Krammer & Lange Bertalot, 1997a, b; Krammer & Lange Bertalot, 2004a, b; Prygiel & Coste, 2000; Segura-García, 2011) también exhiben las mismas condiciones de oligotrofia a mesotrofia en el área de estudio (Tabla 4).
Aunque la temperatura del agua varía con el gradiente altitudinal y con la radiación solar (Pedraza-Garzón, 2011), la temperatura no tuvo relación aparente con la oxigenación, por lo que se observaron temperaturas templadas durante el muestreo y éste no fue un factor determinante en la distribución de las especies.
El oxígeno disuelto se mantuvo con elevadas concentraciones en el manantial y el río Yerbabuena y Las Orquídeas en todas las temporadas (6.0 - 8.0 mg/L) debido a que en esos sitios el flujo de agua es turbulento, tienen pendiente pronunciada, con zonas de rápidos, con fondo pedregoso que favorecen la solubilidad de este gas (Penalta-Rodríguez, 2007). Sin embargo, la baja riqueza de especies observada en El aljibe y La Galera durante el estiaje (2013) se vio relacionada con las bajas concentraciones de oxígeno (4.0 mg/L, para ambos casos). En el primer sitio el flujo de agua era imperceptible, lo que inhibió la solubilidad del oxígeno y en el segundo fueron evidentes los drenajes de las viviendas que se vaciaron al río, además de observarse un florecimiento de Microcystis aeruginosapor lo que la actividad bacteriana abatió las concentraciones de oxígeno, consecuencia del proceso de eutrofización (De León, 2002).
El área de estudio se ve influenciada por las actividades que se desarrollan en el campo geotérmico de Los Azufres, localizado en la parte más alta de la cuenca (2,800 m.s.n.m.). Entre los problemas de alto impacto provenientes de esas labores se encuentran la generación de vapor, la mezcla de agua-vapor y la producción de salmuera, la cual es reinyectada por gravedad al subsuelo (Valdéz et al., 2013). Específicamente la infiltración de la salmuera al manto freático influyó en el aumento de valores de pH que oscilaron de circumneutrales a alcalinos durante la investigación (7.0 - 8.2). Por otro lado, los compuestos derivados de la salmuera también repercuten en el aumento de la concentración de sólidos disueltos totales en las aguas freáticas de la cuenca, muy notorio en La Galera durante 2013 (183 - 767 mg/L), aunque en poslluvias un tramo del río haya estado seco debido a la extracción de agua para el poblado. En Las Orquídeas, en el mismo año (119 mg/L), la elevada concentración se debió a que el banco de materiales ahí establecido aumentó el vertido de residuos hacia el cauce.
En el caso de la alcalinidad los máximos valores se observaron en temporada de lluvias (2013) para dos sitios; La Galera (153.9 mg/L) y Las Orquídeas (119.7 mg/L). Con respecto a las mayores concentraciones de dureza total, se midieron en La Galera durante poslluvias (2012) (153.9 mg/L) y estiaje y lluvias (2013) (171 y 153.9 mg/L, respectivamente), consecuencia del transporte del material río abajo; estas variables se encuentran en relación directa con el comportamiento del pH (Wetzel, 2001). Especies como Amphora ovalis registrada en Las Orquídeas en estiaje 2013 y las especies de Nitzschia, así lo indicaron durante toda la temporada de estudio (Van Dam et al., 1994).
En cuanto a los valores de nitritos y amonio, se encontraron bajas concentraciones de estas variables como resultado de aguas bien oxigenadas (Wetzel, 2001). Sin embargo, estos compuestos fueron determinantes para la presencia de especies como Craticula accomoda, Encyonema silesiacum, Eunotia veneris, E. quaternaria, Frustulia latita, Fragilaria capuccina var. vaucheriae, Gomphonema pseudoagur, Lemnicola hungarica, Navicula angusta, N. cryptocephala, por mencionar algunas, las cuales toleran diversas concentraciones de estos elementos (Krammer & Lange Bertalot, 1997a, b; Krammer & Lange Bertalot, 2004a, b; Segura-García, 2011).
Salvo en el caso del amonio, la máxima concentración se midió en Las Orquídeas durante poslluvias (2013) (1.3 mg/L), lo que indicó que en el sitio hay contaminación de origen industrial (banco de extracción de materiales), por escorrentía de zonas de cultivo, huertas de aguacate y aguas residuales, es decir, elevada eutrofización. Especies como Gomphonema lagenula, G. pseudoaugur, G. pumilum, Navicula symmetrica, Nitzschia clausii, N. palea, Pinnularia interrupta, Planothidium lanceolatum, Rhopalodia brebisonii y Surirella minuta, registradas en el sitio, han sido reportadas para otros países en ríos próximos a terrenos agrícolas cuyas escorrentías contienen alta concentración de fosfatos y nitratos, así como para aguas eutróficas (Cox, 1996; Dere et al., 2006; Germain, 1981; Köster & Hübener, 2001; Krammer & Lange-Bertalot, 2004a, b; 2007a, b; Lange-Bertalot, 2001; Leland & Porter, 2000; Martínez de Fabricius et al. 2003; Ndiritu et al. 2006; Novelo et al. 2007; Novelo, 1998; Szczepocka & Szulc, 2006; Zampella et al. 2007). Lo anterior, es un escenario semejante que prevalece en Las Orquídeas.
En el sitio también se registraron especies como Achnanthidium exiguum, A. minutissimum, Planothidium frequentissimum o Rhoicosphenia abbreviata que son sensibles a la contaminación y reportadas para aguas oligotróficas a mesotróficas (Cox, 1996; Krammer & Lange Bertalot, 1997 a, b; Krammer & Lange Bertalot, 2004a, b; Prygiel & Coste, 2000; Segura-García, 2011), diatomeas provenientes por deriva, desde los manantiales que afloran en la zona y que se incorporan en este tramo del río.
Las diferencias de riqueza específica entre el manantial de la Yerbabuena durante el estiaje (2013) (Fig. 3) y el manantial y río Yerbabuena en la época de lluvias (2013) (Fig. 4) se debió especialmente al cambio de temporada, ya que durante el estiaje la profundidad del río fue menor, por lo que la turbulencia no fue tan intensa. En la temporada de lluvias aumentó el caudal y puesto que es un canal estrecho se generaron remolinos y corrientes circulares, lo que permitió una fuerza hidráulica importante para el transporte de partículas en suspensión (Reid & Wood, 1976). Por tanto, se propició el aumento en el número de especies para esta temporada.
El valor de diversidad de Shannon (H`) sin diferencias significativas para Las Orquídeas y El aljibe en tres temporadas (Tabla 7) fue debido a que en ambas localidades se incorporan manantiales al sistema, lo que generó condiciones similares; además de compartir especies que se consideran indicadoras de aguas oligotróficas como Achnanthidium minutissimum, Planothidium frequentissimum y Rhoicosphenia abbreviata (Cox, 1996; Krammer & Lange Bertalot, 1997a, b; Krammer & Lange Bertalot, 2004a, b; Prygiel & Coste, 2000; Segura-García, 2011). La presencia de Encyonema silesiacum, Eolimna subminuscula, Gomphonema lagenula, especies indicadoras de contaminación (Krammer & Lange Bertalot, 1997a, b; Krammer & Lange Bertalot, 2004a, b; Segura-García, 2011), en ambos sitios revelan condiciones eutróficas, ya que El Aljibe se ve impactado por los agroquímicos de huertas de aguacate y sembradíos cercanos y Las Orquídeas recibe los residuos generados por el banco de materiales establecido en el lugar.
La agrupación entre el río la Yerbabuena (lluvias y poslluvias, 2013) y Las Orquídeas (poslluvias, 2013), obtenida en el análisis por temporadas y sitios, indicaría que su afinidad es consecuencia de las condiciones del río en esta zona, es decir, aguas con elevada oxigenación, con tendencia a la alcalinidad. En ambos sitios se registró a Plantohidium frequentissimum como indicadora de medios oligotróficos; en tanto, Navicula rhynchocephala, Nitzschia palea y Planothidium lanceolatum señalan la incorporación de materia orgánica al sistema (Cox, 1996; Krammer & Lange Bertalot, 1997a, b; Krammer & Lange Bertalot, 2004a, b; Segura-García, 2011). En cuanto a La Galera, el río cruza en medio del poblado y los drenajes de las viviendas se vierten directamente al cauce sin ningún tipo de tratamiento. Las especies que estuvieron presentes en esta localidad son consideradas como tolerantes a niveles críticos de contaminación, indicadoras de ambientes hipereutróficos, bajas concentraciones de oxígeno disuelto y aumento de sólidos disueltos (Van Dam et al., 1994; Cox, 1996; Lange-Bertalot, 2001; Krammer & Lange-Bertalot, 2004b; Segura-García, 2011) (Fig. 6). Entre las especies que destacaron por su abundancia se encuentran Navicula germainii, N. recens, Nitzchia clausii, N. inconspicua y N. palea (Tabla 4).
De los grupos formados a partir del ACC, el perteneciente al del río Yerbabuena incluyó especies adaptadas a elevadas concentraciones de materia orgánica; el de La Galera es un buen indicador de ambientes perturbados por actividad humana (escorrentías de aguas residuales y potreros) (Lange-Bertalot, 2001; Krammer & Lange-Bertalot, 2004b; Metzeltin & Lange-Bertalot, 1998; Segura-García, 2011); mientras que el último grupo tolera bien la desecación y se encuentran en aguas oligotróficas a mesotróficas (Van Dam et al., 1994; Lange-Bertalot, 2001; Krammer & Lange-Bertalot, 2004b; Metzeltin & Lange-Bertalot, 1998; Segura-García, 2011; Segura-García et al., 2012).
Las especies de diatomeas encontradas en el presente estudio reflejaron las condiciones ambientales de cada sitio y temporada en las que fueron recolectadas, puesto que se identificaron especies indicadoras de aguas oligotróficas, ambiente inherente al nacimiento de un río, y especies que revelan contaminación proveniente de actividad urbana, industrial y agrícola.
Los resultados del presente estudio dieron a conocer que los géneros de diatomeas epilíticas mayormente representados fueron Navicula con 11 especies, Nitzschia y Gomphonema con 7 y Pinnularia con 6, todas pertenecientes al orden Pennales.
Fue evidente que las actividades humanas en la cabecera del río Zinapécuaro están modificando las características fisicoquímicas del agua, así como las condiciones ambientales en las que se encuentran los organismos. Especies de diatomeas previamente reportadas como indicadoras de sitios eutrofizados para otras regiones de México y del mundo fueron encontradas en este río como un efecto del impacto antrópico.
El análisis de las comunidades de diatomeas epilíticas de la cabecera del río Zinapécuaro ha demostrado ser muy útil para la evaluación de la variación espaciotemporal asociada a ella, sin embargo, es necesario continuar con muestreos estacionales que permitan caracterizar las condiciones ecológicas del cuerpo de agua, la utilidad de la composición, abundancia y distribución de las especies de diatomeas como bioindicadores de éste y otros sistemas acuáticos.

