











 nova página do texto(beta)
nova página do texto(beta) Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO 
 Permalink
PermalinkIntroducción
El grupo de las diatomeas es uno de los productores primarios más importantes de los océanos y posiblemente contribuye entre el 3040% de la producción primaria total (Nelson et al., 1995; Tréguer et al., 1995). Como tal, las diatomeas son una fuente de carbono exportado a los sedimentos (~55% del carbono total; Sarmiento & Gruber, 2006) desde aguas superficiales, particularmente en latitudes altas y regiones de surgencia (Buesseler, 1998). Así, la producción y el ciclo del ópalo biogénico es un componente importante de la bomba biológica del océano (Ragueneau et al., 2006; Sarmiento & Gruber, 2006) y juega un papel importante en la regulación del intercambio de CO entre el 2002; Sarmiento et al., 2004).
El ácido silícico es un nutriente importante en el océano superficial ya que se requiere para el crecimiento de diatomeas, radiolarios y silicoflagelados. Dado que la producción de sílice biogénico depende del suministro de ácido silícico disuelto a la zona eufótica, el silicio (Si) es un elemento limitante importante. Las diatomeas dominan el ciclo biogeoquímico del Si, e influyen fuertemente en los ciclos biogeoquímicos de otros macronutrientes, como el nitrógeno (N) y fósforo (P) (Reynolds et al., 2006). Globalmente, el ácido silícico es efectivamente atrapado en el Océano del Sur, una implicación importante es que las diatomeas de otros océanos están limitadas por ácido silícico y no por nitrato (Matsumoto et al., 2014).
En condición ideal, las diatomeas saludables necesitan ácido silícico y nitrato biológicamente disponible con una razón 1:1 y este valor puede llegar a ser de aproximadamente 3 en condiciones de limitación por hierro (Brzezinski, 1985). Si la producción superficial está dominada por las diatomeas, lo que no excluye ni descarta la presencia de otros productores primarios (e.g., cocolitofóridos), la relación Si:N de la producción biogénica debe mantenerse por debajo o alrededor de 3. En el Océano del Sur y el Pacífico ecuatorial, la adición de Fe ocasionó una disminución en la razón Si:N de 4:1 a 1:1 (Brzezinski et al., 2003). La causa exacta de este cambio en la razón Si:N sigue en debate (e.g., Matsumoto et al., 2014). Algunos de los factores responsables pueden incluir una reducida silificación de las frústulas de las diatomeas (e.g., Franck et al., 2000), diferencias en condiciones de crecimiento, cambios en la composición de las especies y la variabilidad genética o cambios morfológicos (Marchetti & Cassar, 2009).
Estudios recientes realizados en el Golfo de California, y en el margen de Mazatlán, sugieren que los períodos de surgencia están asociados con una limitación de Fe transitoria, lo que da como resultado una alta producción exportada de sílice biogénico relativo al carbono orgánico (Pichevin et al., 2012, 2014). Una correlación similar entre el enterramiento de ópalo biogénico y la limitación de Fe es evidente en registros sedimentarios en distintas escalas de tiempo (Arellano-Torres et al., 2011; Pichevin et al., 2012, 2014). De esta forma, las condiciones de limitación de ácido silícico que prevalecen en el océano son causadas en gran parte por la deficiencia de hierro lo que conlleva a una restricción indirecta sobre la captación del carbono oceánico (Pichevin et al., 2014).
Se ha demostrado que en el Pacífico nororiental ocurre alternancia en la dominancia del plancton calcáreo a opalino, este último es dominante en períodos de surgencias, durante los cuales la producción de carbonatos es baja (Thunell et al., 1995; Fisler & Hendy, 2008).
En Cuenca Cariaco esta sucesión de grupos estuvo caracterizada por una razón BSi/C que alcanzó sus máximos valores durante la presencia del grupo de las diatomeas, y fue menor bajo condiciones de una columna de agua estratificada, dando paso a la dominancia de grupos calcáreos (Goni et al., 2003). En el margen de Magdalena, la excelente preservación de una capa de cocolitofóridos entre 13000 y 12000 años antes del presente (Van Geen et al., 2003) precede a un máximo en la razón BSi/C entre 12000 y 11000 años antes del presente, confirmando la alternancia en la comunidad planctónica (Sánchez & Carriquiry, 2007). Una compilación global también indica que algunos sitios de enterramiento de sílice biogénico en el océano están caracterizados por una alta razón de sílice biogénico relativo al carbono orgánico, una característica diagnóstico para diatomeas que crecen bajo estrés de Fe (Arellano-Torres et al., 2011; Pichevin et al., 2012, 2014). Por lo tanto, la alternancia de grupos puede ser inferida a partir de cambios en la razón BSi/C y BSi/N (Goni et al., 2003; Sánchez & Carriquiry, 2007; Arellano Torres et al., 2011; Pichevin et al., 2012, 2014).
En este trabajo se presenta evidencia de que el grupo de las diatomeas dominó sobre los grupos calcáreos en el Pacífico nororiental mexicano durante el último milenio, lo cual sustentará la hipótesis de que en el margen de Magdalena, tal como ocurre en el margen de Mazatlán (Arellano-Torres et al., 2011) y Cuenca de Guaymas (Pichevin et al., 2012, 2014), el incremento de sílice biogénico relativo al nitrógeno total estará asociado a una limitación transitoria de Fe, y no por la deficiencia de nitrógeno (Moore et al., 2004) y fósforo (Matsumoto et al., 2014). Asimismo, se propone que el flujo de nutrientes exportados del Golfo de California, a través de su margen occidental hacia el Pacífico nororiental (Bustos-Serrano & Castro, 2006; Alvarez-Borrego, 2012) es un mecanismo que ayudará a la fertilización en el margen de Magdalena.
Materiales y métodos
El núcleo de sedimento del margen de Magdalena (multinúcleo MAGDMC-06; 42 cm, Fig. 1) fue colectado en octubre de 2009 en el crucero "Nitrogen Dynamics" a bordo del R/V "New Horizon" y cubre aproximadamente los últimos mil años. Éste fue muestreado a intervalos de 1 cm para obtener un registro de alta resolución de contenido de nitrógeno total, sílice biogénico y fósforo orgánico. El nitrógeno total se midió en un analizador elemental Costech 4010 en el Laboratorio de Espectrometría de Masas del CICIMAR IPN (LEsMa). El sílice biogénico y el fósforo orgánico fueron determinados por el método espectrofotométrico azul de molibdeno utilizando la técnica de Mortlock & Froelich (1989) y de Aspila (1976), respectivamente.
Figura1 Área de estudio y localización del sitio de colecta del núcleo MAGD-MC06 en el margen suroccidental de la península de Baja California.
El modelo de edad del núcleo MAGD-MC-06 que se utilizó en este estudio es el desarrollado por Van Geen et al. (2003) y Marchitto et al. (2010), y fue construido utilizando mediciones de 14C por AMS en foraminíferos bentónicos (análisis realizados en el National Ocean Sciences Accelerator Mass Spectrometry facility en Woods Hole, Massachusetts). Los datos de radiocarbono fueron convertidos en años calendario usando el software de calibración CALIB 4.3 (Stuvier & Reimer, 1993).
Resultados
El contenido de ópalo biogénico tiene un promedio y desviación estándar de 7 ± 0.6% con un incremento casi constante a lo largo del registro sedimentario del margen de Magdalena. De manera general se distinguen tres períodos de variabilidad en el contenido de ópalo biogénico: (1) En el período de 860 a 570 años, el contenido de ópalo biogénico fue de 6.8 ± 0.2%; (2) Para el período de 550 a 330 años fue de 7.5 ± 0.2%, un período de poca variabilidad en cuanto al ópalo biogénico exportado, período caracterizado por el último enfriamiento global conocido como la Pequeña Edad de Hielo (1550 a 1850 años DC); y (3) En los últimos 330 años, el contenido de ópalo biogénico fue de 8.0 ± 0.3% (Fig. 2a).
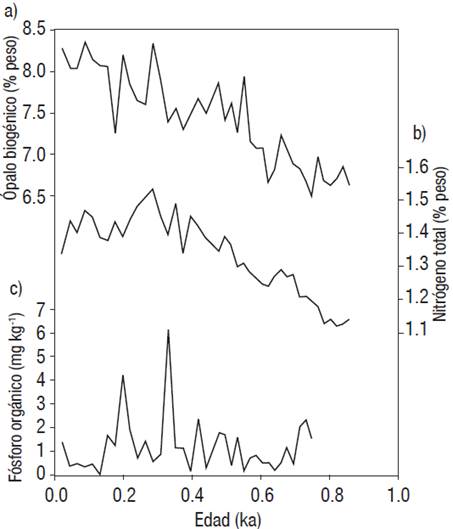
Figuras 2a-c Contenido de (a) Ópalo biogénico, (b) Nitrógeno total (porcentaje en peso) y (c) Fósforo orgánico (mg kg-1) para el núcleo MAGD-MC06 en los últimos 900 años, margen suroccidental de la península de Baja California.
El contenido de nitrógeno total tuvo un promedio y desviación estándar de 1.3 ± 0.1%, y al igual que el ópalo biogénico, tuvo un incremento constante a lo largo del registro para el período de 860 a 290 años, en los últimos 290 años se denota un ligero decremento en el contenido de nitrógeno total (Fig. 2b).
El fósforo orgánico tuvo un promedio y desviación estándar de 1.1± 0.9 mg kg-1 con mínimo de 0.1 y máximo de 4.2 mg kg-1. A lo largo del registro sedimentario, el fósforo orgánico mantuvo casi siempre valores menores a 2 mg kg-1.
Discusión
El ópalo biogénico representa la contribución relativa de organismos (principalmente diatomeas con una menor contribución de radiolarios y silicoflagelados) que exportan ópalo al sedimento. El contenido de ópalo en el sedimento puede ser afectado por la redistribución del sedimento, la preservación diferencial durante el asentamiento y el enterramiento de las partículas, así como la dilución por otros componentes del sedimento (Ragueneau et al., 2000). Estos factores pueden dar lugar a interpretaciones erróneas en el uso del ópalo biogénico como trazador de los cambios en la productividad del plancton silíceo (Arrellano-Torres et al., 2011). Se ha documentado que estos factores no afectan el contenido de ópalo biogénico en registros del margen oriental del Pacífico mexicano y Golfo de California (Sancetta, 1995; Pride et al., 1999; Barron et al., 2005; Barron & Bukry, 2007; Sánchez & Carriquiry, 2007; Arellano-Torres et al., 2011; Barron et al., 2012; Pichevin et al., 2012, 2014).
La tasa de acumulación del sedimento fue constante (Van Geen et al., 2003; Marchitto et al., 2007) y no se observó ninguna variación en el tamaño de grano, a lo largo del núcleo, esto sugiere que no ha existido redistribución del sedimento en el margen de Magdalena. La preservación y dilución no fueron factores que alteraron la exportación de ópalo biogénico, ya que mantiene el mismo comportamiento del nitrógeno total, el cual es más lábil que la matriz mineral de sílice biogénico. Por lo tanto, el contenido de ópalo biogénico y nitrógeno total reflejan variaciones en el suministro de ópalo y nitrógeno de la columna de agua al sedimento en el margen de Magdalena.
Los registros sedimentarios disponibles para el margen de Magdalena se limitan a los últimos 55 mil años (Sánchez & Carriquiry, 2007; Barron et al., 2012; Carriquiry y Sánchez, 2014) y en el Golfo de California a los últimos 40 mil años (Sancetta, 1995; Pride et al., 1999; Barron et al., 2005; Barron & Bukry, 2007; Arellano-Torres et al., 2011; Pichevin et al., 2012; Ricaurte-Villota et al., 2013; Pichevin et al., 2014) donde se ha documentado la variabilidad en el contenido de sílice biogénico, así como en la composición de las especies de diatomeas, radiolarios y silicoflagelados.
La variabilidad en el contenido de ópalo biogénico es de 14 a 2% en el margen oriental del Pacífico mexicano (Golfo de Tehuantepec y margen de Mazatlán, Arellano-Torres et al., 2011; y margen de Magdalena, Sánchez & Carriquiry 2007; Barron et al., 2012), en el Golfo de California es de 50 a 10% en Cuenca de Guaymas (Pichevin et al., 2012, 2014) y de 7 a 2% en Cuenca Alfonso (Ricaurte-Villota et al., 2013).
En el último milenio, los registros de ópalo biogénico mostraron una variabilidad de 9 a 6% en el margen de Magdalena (Fig. 3a, este estudio), de 7 a 4% en Cuenca Alfonso (Fig. 3b, Ricaurte-Villota et al., 2013), de 17 a 13% en Cuenca Pescadero (Fig. 3c, Barron & Bukry, 2007) y de 40 a 20% en Cuenca de Guaymas en el Golfo de California (Fig. 3d, Barron & Bukry, 2007).
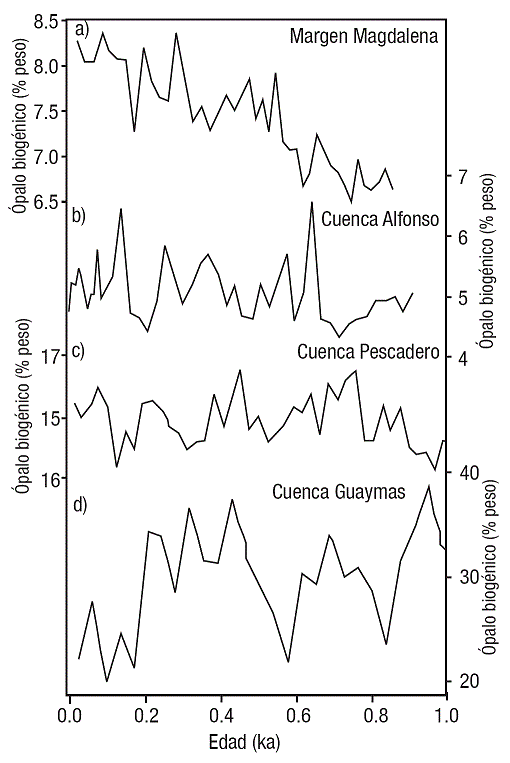
Figuras 3a-d Registros del contenido de ópalo biogénico (porcentaje en peso) para el último milenio en núcleos de sedimento colectados en el Pacífico nororiental mexicano. (a) Núcleo de sedimento MAGD-MC06 en el margen Magdalena (este estudio); (b) Núcleo de sedimento NH01-15MC1 en Cuenca Alfonso, Golfo de California (Ricaurte-Villota et al., 2013); (c) Núcleo de sedimento NH0126 en Cuenca Pescadero, Golfo de California (Barron & Bukry, 2007); y (d) Núcleo de sedimento BAM80 E-17 en Cuenca Guaymas, Golfo de California (Barron & Bukry, 2007).
El ópalo biogénico muestra una clara tendencia de incremento hacia el reciente para el margen de Magdalena, mientras los registros del Golfo de California no presentaron esta misma tendencia y se mantiene el gradiente en el contenido de ópalo biogénico siendo menor en la región sur y mayor hacía la región central del Golfo (Pichevin et al., 2012, 2014). Esto indica que el margen de Magdalena experimentó condiciones en el océano que favorecieron la exportación hacia los sedimentos de ópalo biogénico del grupo de las diatomeas (y contribución de radiolarios y silicoflagelados) sobre el plancton calcáreo. Esta evidencia es soportada por registros sedimentarios colectados a lo largo de la corriente de California. En el margen suroccidental de Baja California se presentó un incremento en las surgencias a partir de 7500 años antes del presente, asociado a la reorganización atmosférica en Norteamérica (Barron et al., 2012; Fig. 2). Así mismo, los trazadores de la productividad exportada (i.e. ópalo biogénico) en Punta Concepción en el margen de California mostraron un incremento continuo para los últimos 1500 años (Hendy et al., 2004). Apoyando la idea de que la productividad exportada fue dominada por el grupo del plancton silíceo, principalmente diatomeas a lo largo de la corriente de California (este estudio, Hendy et al., 2004; Sánchez & Carriquiry, 2007; Barron et al., 2012).
Registros sedimentarios de los últimos 40 mil años a lo largo del margen oriental del Pacífico Tropical han sugerido que el incremento en la exportación de ópalo biogénico (Arellano-Torres et al., 2011) fue producto del suministro de ácido silícico hacia bajas latitudes por el Océano del Sur (Brzezinski et al., 2002; Matsumoto et al., 2002). La disminución en la razón Si:N de 2.8 a 0.9 fue interpretado como un cambio en la dominancia de plancton silíceo hacia algas no silícicas en el margen de Mazatlán (Arellano-Torres et al., 2011) y en la fuente de ácido silícico hacia esta región tropical.
En los últimos 1500 años no existen evidencias documentadas de un suministro de ácido silícico del Océano del Sur hacia otros océanos (ya que éste queda "secuestrado" en esta región, Matsumoto et al., 2014). Sin embargo, la razón Si:N de 2.8±0.1 en el margen de Magdalena (Fig. 4), así como el incremento de ópalo biogénico reportado por Hendy et al. (2004) y Barron et al. (2012) a lo largo de la corriente de California, sugiere que otros mecanismos de fertilización del océano fueron los responsables de mantener el incremento en la exportación de ópalo biogénico en el área de estudio.
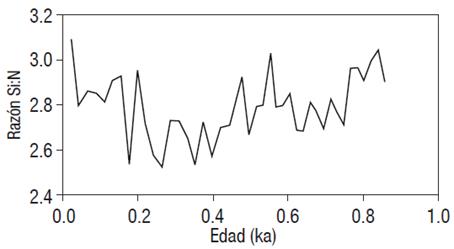
Figura 4 Razón Si:N (molar) para el núcleo MAGD-MC06 en los últimos 900 años, margen suroccidental de la península de Baja California.
La concentración del ácido silícico y nitratos (Van Geen et al., 2003), y la razón Si(OH)4 :NO3- en el margen de Magdalena es mostrada en las Figs. 5a-b. Las concentraciones de ambos nutrientes son similares a los promedios anuales reportados para la entrada del Golfo de California (Bustos-Serrano y Castro, 2006) y cuenca de Guaymas (Pichevin et al., 2012). Al igual que en cuenca de Guaymas, la razón silicatos:nitratos es >1 en aguas subsuperficiales y estos valores son mayores a la razón Si:N para diatomeas saludables creciendo bajo condiciones no limitantes de nutrientes (Brzezinski, 1985). Dos mecanismos han sido propuestos para explicar esta alta razón Si:N para el Golfo de California y en este trabajo se propone la hipótesis de un tercer mecanismo: (1) La remoción de nitrato en el agua subsuperficial e intermedia por desnitrificación (Altabet et al., 1999; Pride et al., 1999; Deustch et al., 2014); (2) La acumulación de ácido silícico en el Golfo de California a través del ciclo del ópalo biogénico y la liberación de Si del detritus en hundimiento (Pichevin et al., 2012, 2014); y (3) La exportación de nutrientes (ácido silícico, Pichevin et al., 2012, 2014, y nitratos, Álvarez-Borrego, 2012) del Golfo de California hacia el Océano Pacífico y su posible dispersión y contribución a la fertilización del margen suroccidental de Baja California Sur.
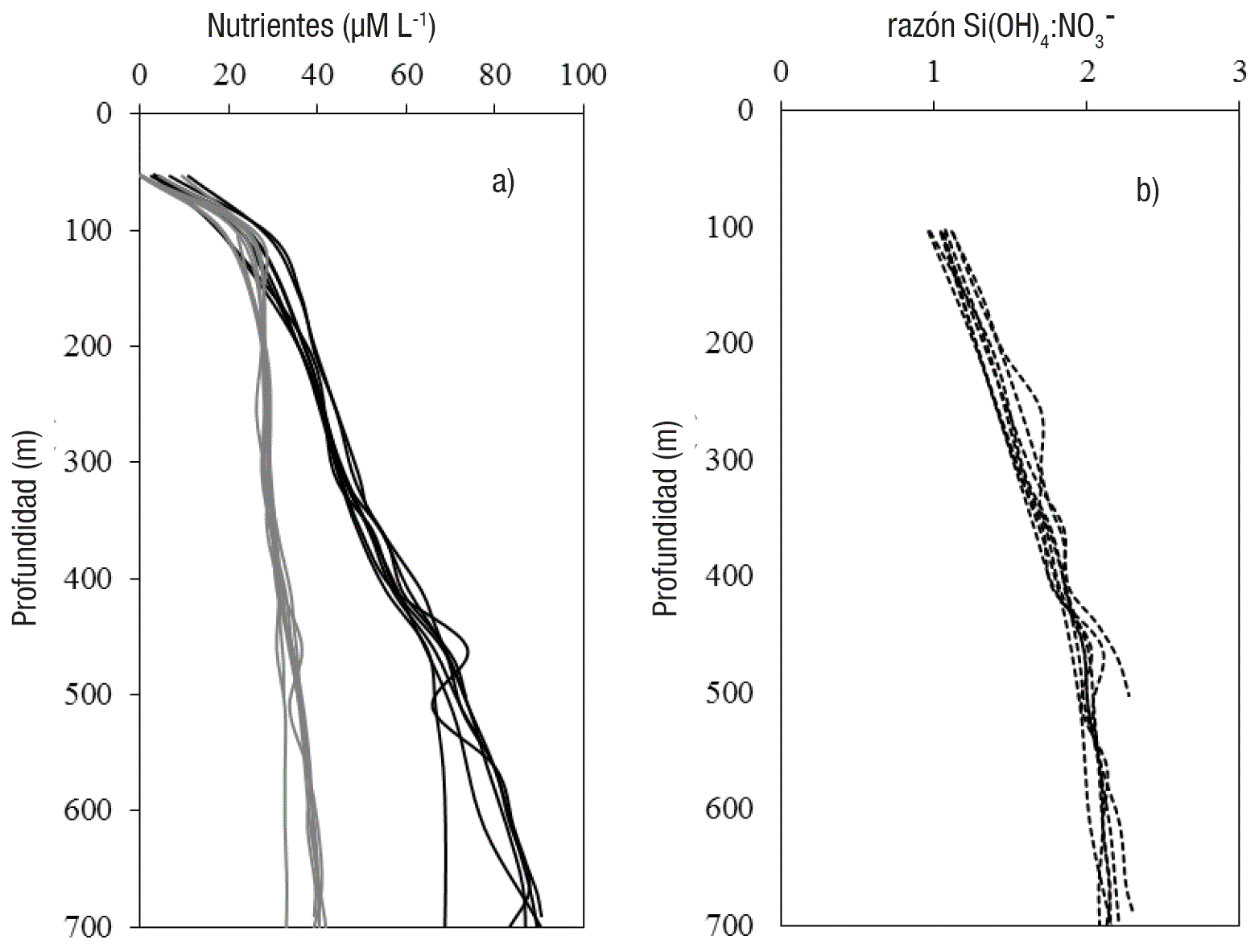
Figuras 5 a-b. a) 1 Perfiles de ácido silícico (líneas continuas negras) y nitratos (líneas continuas grises). b) Perfil de la razón ácido silícico:nitratos (molar) con la profundidad de la columna de agua, colectados en octubre-noviembre de 1999, durante la campaña oceanográfica OMX01-Melville99 en el margen suroccidental de la península de Baja California.
La desnitrificación en el Pacífico nororiental mexicano es intensa y debajo de la nutriclina la concentración de los nitratos es similar en el margen de Magdalena (Van Geen et al., 2003), la boca del Golfo de California (Bustos-Serrano & Castro, 2006) y cuenca de Guaymas (Pichevin et al., 2012). Por lo tanto, la desnitrificación local, como primer mecanismo, no puede explicar la razón Si(OH)4 :NO3- presente en el margen de Magdalena y el Golfo de California. El segundo mecanismo, la alta concentración de ácido silícico, debido a la regeneración local (disolución de ópalo) explica la alta razón Si(OH)4 :NO3- en aguas subsuperficiales e intermedias en el Golfo de California (Pichevin et al., 2012). El tercer mecanismo sugerido es la exportación de nutrientes del Golfo hacia el margen suroccidental de Baja California Sur y está sustentado en la dinámica de la boca del Golfo (i.e., Lavín & Marinone, 2003; Mascarenhas et al., 2004). La interacción de las corrientes del Golfo de California, de California y costera Mexicana generan remolinos ciclónicos y anticiclónicos que se mueven en dirección oeste y noroeste (Cepeda-Morales et al., 2009; Pantoja et al., 2012) y este mecanismo puede ser responsable de que se exporten nutrientes del Golfo de California hacia el Océano Pacífico (Álvarez-Borrego, 2012) y sean llevados hacia el margen suroccidental de Baja California Sur, como se ha sugerido para un registro sedimentario de baja resolución en el margen de Magdalena (Barron et al., 2012). Finalmente, la razón Si:N de 2.8±0.1 sugiere el crecimiento de diatomeas saludables para el último milenio, considerando, que un valor aproximado de 3 es diagnóstico de limitación por hierro y no de nutrientes, como nitratos y fosfatos (Brzezinski, 1985; Arellano-Torres et al., 2011; Pichevin et al., 2012, 2014).

