











 nova página do texto(beta)
nova página do texto(beta) Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO 
 Permalink
PermalinkIntroducción
El fitoplancton marino lleva a cabo diversas funciones ecológicas que le confieren una importancia básica en el ecosistema pelágico de todo el mundo. Con el advenimiento de varias nuevas tecnologías, los protocolos de investigación de esta importante comunidad se han sistematizado y la información que se aporta cotidianamente incluye la distribución espacial tridimensional, las distintas contribuciones de las fracciones de talla (pico, nano y microplanton) y de las diferentes entidades taxonómicas (e.g. diatomeas, dinoflagelados, cocolitofóridos y otros) y datos de metagenómica, para conocer más sobre su biodiversidad y ecología.
El análisis de los pigmentos fotosintéticos del fitoplancton constituye actualmente una herramienta muy útil para reconocer grupos taxonómicos, así como sus distribuciones espaciales (geográficas y verticales) y las contribuciones a la biomasa total. El principio de este método es la existencia de pigmentos diagnósticos de ciertos grupos taxonómicos, aunque también hay pigmentos que son compartidos por dos o más grupos taxonómicos (Jeffrey & Vesk, 1997). Los análisis se efectúan rutinaria, rápida y eficientemente por medio de HPLC (Cromatografía Líquida de Alta Precisión, por sus siglas en inglés) (Vidussi et al., 1996; Zapata et al., 2000).
En ciertas épocas y varias porciones del Golfo de Tehuantepec se han estudiado la composición, abundancia y distribución de ciertos grupos taxonómicos del fitoplancton (Hernández-Becerril, 1987, 1988, 1990; Hernández-Becerril et al., 1993, 2013; Meave & Hernández-Becerril, 1998; Aké-Castillo et al., 1999; Hernández-Becerril & BravoSierra, 2001; Maciel-Baltazar & Hernández-Becerril, 2013), incluyendo el hallazgo de nuevos registros y especies (Bravo-Sierra &HernándezBecerril, 2003; Hernández-Becerril & Bravo-Sierra, 2004a, 2004b; Hernández-Becerril & Díaz-Almeyda, 2006; Gárate-Lizárraga, 2008; Hernández-Becerril & Barón-Campis, 2008), así como la revisión del fitoplancton tóxico y nocivo de la zona (Hernández-Becerril et al., 2007), y también se han medido la biomasa (básicamente como clorofila a) y producción primaria fitoplanctónica (Robles-Jarero & Lara-Lara, 1993; Lara-Lara & Bazán-Guzmán, 2005).
Se conoce que durante la época de vientos "Tehuanos" (invierno y principio de primavera) y la consecuente mezcla turbulenta producida por éstos (Färber-Lorda et al., 2004; Liang et al., 2009), el fitoplancton del Golfo de Tehuantepec actúa como una "bomba de Carbono" (Robles-Jarero & Lara-Lara, 1993; Färber-Lorda et al., 2004), y que las comunidades fitoplanctónicas que aparecen durante tales escenarios son dominadas en diversidad y biomasa principalmente por especies de diatomeas, quienes responden eficientemente a una alta turbulencia, mientras que en condiciones de relativa estabilidad, los dinoflagelados aparecen como el grupo más diverso y quizás más abundante (Meave & Hernández-Becerril, 1998). Aparentemente también varían las contribuciones de las diferentes fracciones de talla del fitoplancton a la biomasa total y productividad primaria, en distintas épocas climáticas (Lara-Lara & Bazán-Guzmán, 2005).
Sin embargo se sabe menos de las comunidades fitoplanctónicas de verano, cuando la concentración superficial de clorofila a es más baja, por ejemplo en junio (Lluch-Cota et al., 1997), y se suponen condiciones más tropicales, con mayor estratificación y decremento de concentraciones de nutrientes. En este trabajo se dan a conocer la composición y distribución de cinco pigmentos fotosintéticos del fitoplancton del Golfo de Tehuantepec en condiciones de verano, y una interpretación y discusión de las diferentes fases estacionales o temporales de esta interesante área de estudio desde la perspectiva del fitoplancton.
Material y métodos
Área de estudio. El Golfo de Tehuantepec se caracteriza principalmente por la presencia de dos épocas bien definidas: (1) durante invierno y principios de primavera se presentan vientos intensos del Norte, con velocidades de hasta 25 m s-1, siendo éste el mecanismo de generación de surgencia, y (2) durante el verano, con vientos del Este, generalmente de menor intensidad y donde se presenta estratificación térmica y lluviosa (Roden, 1961; Liang et al., 2009). Esta área muestra fluctuaciones estacionales e interanuales (Wyrtki, 1974) que determinan la incidencia de la Corriente Costera del Norte (invierno y primavera), la corriente de Costa Rica (verano y otoño) (Monreal & Salas, 1998), la corriente Norecuatorial y la corriente Surecuatorial (Molina-Cruz & Martínez-López, 1994). Sin embargo, se ha observado una clara influencia de procesos costeros en la época de fuertes vientos durante la cual se presentan fenómenos de surgencias eólicas y mezcla turbulenta, debido al trasporte de Ekman se genera una pendiente en la termoclina con mayor profundidad en la parte Oeste, mientras que en el lado Este, ésta se encuentra más somera (Lavín et al. 1992). En la época de surgencias, la termoclina se eleva hasta 10 m generando una franja de baja temperatura que llega a presentar 200 km de anchura y 500 km de longitud con temperaturas superficiales de 6-10° C por debajo de la temperatura promedio que es de 24-26° C (Roden 1961; McCreary et al. 1989; Monreal-Gómez & Salas-de-León 1998). La concentración de nutrientes (nitratos) aumenta en la zona fertilizada por la mezcla turbulenta y se extiende por el borde de la plataforma continental hasta su cruce con el Cañón del Río Suchiate (Machain-Castillo et al., 2008).
Por otro lado, durante el verano se forma una alberca de agua caliente con una temperatura media superficial superior a los 28° C, que se considera una consecuencia de los patrones de circulación, ya que las corrientes en el Pacífico mexicano son muy lentas, por lo que las masas de agua son expuestas durante largo tiempo a la insolación, resultando en elevadas temperaturas superficiales (Trasviña et al., 1999). Bajo estas condiciones, la concentración de nutrientes es consistente con el patrón de circulación mostrando un flujo hacia el Oeste y la ausencia de fertilización por efecto del viento, aunque en verano, la presencia de un giro ciclónico en el Este del golfo contribuye a la fertilización (Machain-Castillo et al., 2008).
Obtención de datos y muestras. Se obtuvieron datos ambientales, básicamente datos provenientes de CTD: temperatura, salinidad y oxígeno disuelto, y muestras de botella Niskin (10 L) para pigmentos fotosintéticos en 9 estaciones, distribuidas tres en cada zona: Oeste (profunda), Centro y Este (más somera) (Tabla 1, Fig. 1), durante el crucero oceanográfico "Tehua 2003", efectuado a bordo del B/O "El Puma", en junio, 2003. Las estaciones de la zona Este tuvieron profundidades menores a 100 m (Tabla 1).
Tabla 1 Profundidad de las estaciones en el Golfo de Tehuantepec, de la capa de mezcla (posición de las termoclinas principales) en cada estación y de los picos de máxima concentración de pigmentos fotosintéticos, todas en metros (m). Dv Cl a, Fuc y Zea, corresponden a Divinil-clorofila a, Fucoxantina y Zeaxantina, respectivamente.
Figura 1 Mapa del Golfo de Tehuantepec, mostrando la localización de las estaciones de muestreo (1-9), en las zonas Oeste (1-3), Centro (4-6) y Este (7-9).
Se tomaron muestras para el análisis de pigmentos a 5 niveles en profundidad (superficie, 20, 30, 40 y 70 ó 100 m, según la profundidad de cada estación) para cada estación, y se filtraron 2 L de agua por cada muestra, a través de filtros GF/F Whatman de microfibra de vidrio de 25 mm de diámetro, con ayuda de una bomba de vacío. Las mues tras se congelaron (de -10 a -15° C) en un tanque de Nitrógeno líquido y se transportaron al laboratorio para su análisis.
Análisis de muestras (HPLC). Para la extracción de pigmentos se siguió el método de Vidussi et al. (1996), que es sensible a la detección de Divinil-clorofila a. A cada filtro se agregaron 3 mL de metanol y se sonicaron (utilizando un Sonicador Sonifier 250) 6 ciclos de 30:30 s, por muestra; 24 h después se obtuvo el extracto, centrifugando durante 5 min a 1500 rpm. De cada extracto se inyectaron 400 μL al sistema HPLC (Autosampler: Waters 717 plus, Controller: Waters 600, Degassit: 6324 y Photodiode Array Detector: Waters 996), en el cual se utilizó una precolumna y columna C8 (Shandon), de 3 μm de tamaño de partícula Hypersil® MOS de 10 cm, 4.6 mm (diámetro interno) (Vidussi et al., 1996). La elusión se hizo fluir a razón de 1 mL min-1 usando un gradiente binario lineal entre el solvente A (Metanol al 0.5N y Acetato de Amonio en solución acuosa, en una proporción de 70:30%) y el solvente B (Metanol 100%), los cuales se programaron mediante un procedimiento de adhesión (minutos; % solvente A, % solvente B): (0; 75, 25), (1; 50, 50), (15; 0, 100), (18.5; 0, 100), (19; 75, 25). La identificación de pigmentos se realizó por la comparación de los tiempos de retención, en los cuales se presentaba un pico determinado de pigmentos estándares, que sirven como calibradores, con los tiempos y picos obtenidos en las muestras. Los estándares provinieron de DHI Lab Products (Hoersholm, Dinamarca). Se hicieron los cálculos de las concentraciones, considerando las áreas de los picos y los volúmenes filtrado e inyectado. Los pigmentos considerados en este trabajo fueron Fucoxantina (que se considera representativo de las diatomeas), Prasinoxantina (pigmento mayoritario de las Prasinophyceae o algas verdes primitivas), Violaxantina (pigmento presente en algas verdes y Eustigmatophyceae), Zeaxantina (considerado un pigmento indicador de las Cianobacterias, particularmente de Synechococcus, organismo picoplanctonte) y Divinil-clorofila a (pigmento diagnóstico de Prochlorococcus, un género de muy amplia distribución y altas densidades celulares, perteneciente al picoplancton) (Jeffrey & Vesk, 1997).
Resultados
Condiciones hidrográficas. La temperatura superficial presentó valores cercanos a 30° C en todas las estaciones. Los perfiles verticales indicaron condiciones de alta estratificación de la columna de agua en todas las estaciones, con capas de mezcla bien definidas y termoclinas entre 20 y 35 m de profundidad (Tabla 1, Figs. 2, 3, 4). En tanto la salinidad varió entre 33.7 y 34.86, generalmente incrementándose con la profundidad (Figs. 2, 3, 4), aunque en las estaciones 2, 3 y 5 hubo un súbito incremento (la mayor diferencia fue de 34.2 hasta 34.7 en la estación 2) en la capa subsuperficial (entre 10 y 30 m).
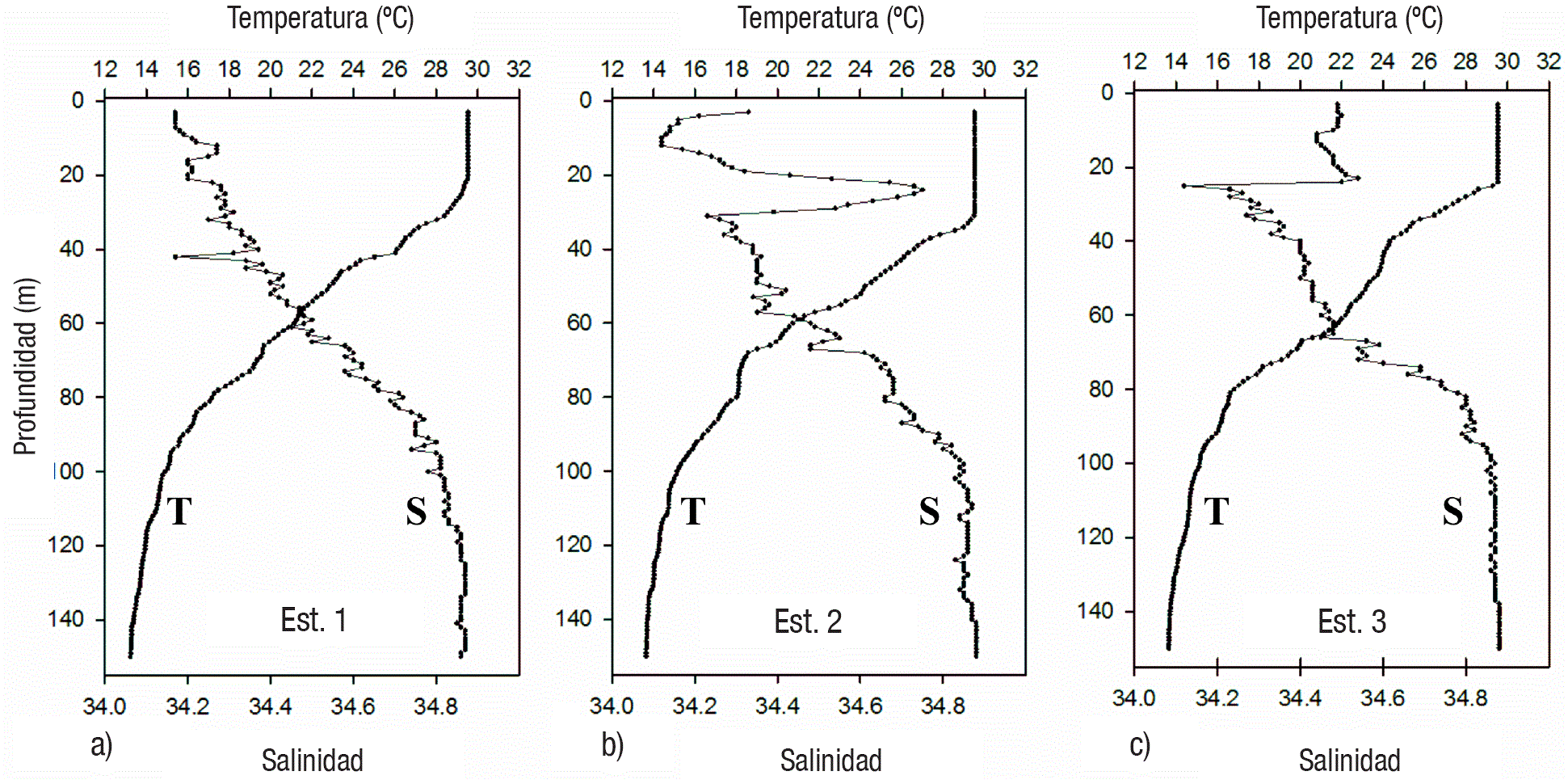
Figuras 2a-c Perfiles verticales de temperatura y salinidad en las estaciones de la zona Oeste (1-3) del Golfo de Tehuantepec.
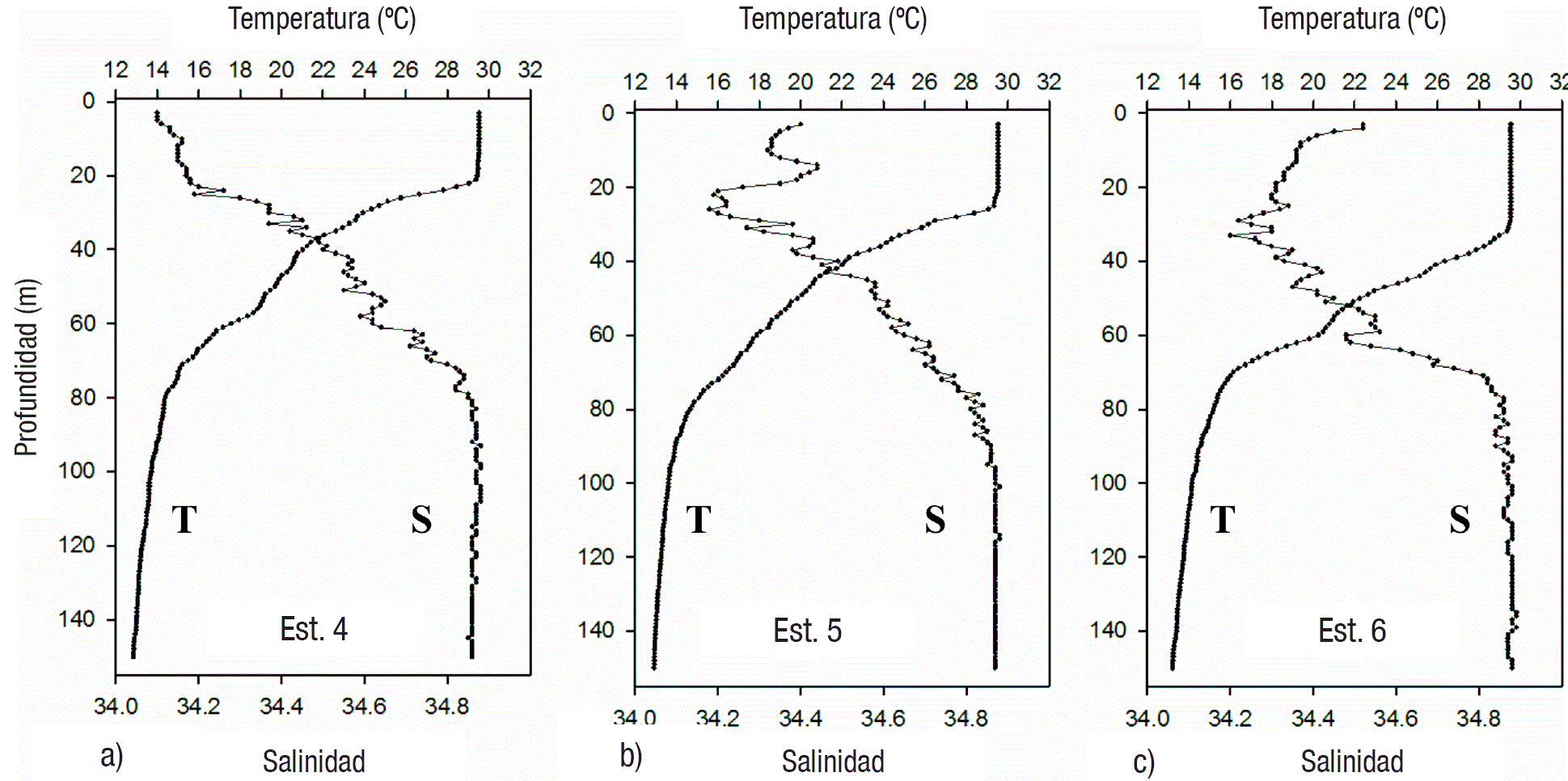
Figuras 3a-c Perfiles verticales de temperatura y salinidad en las estaciones de la Centro (4-6) del Golfo de Tehuantepec.
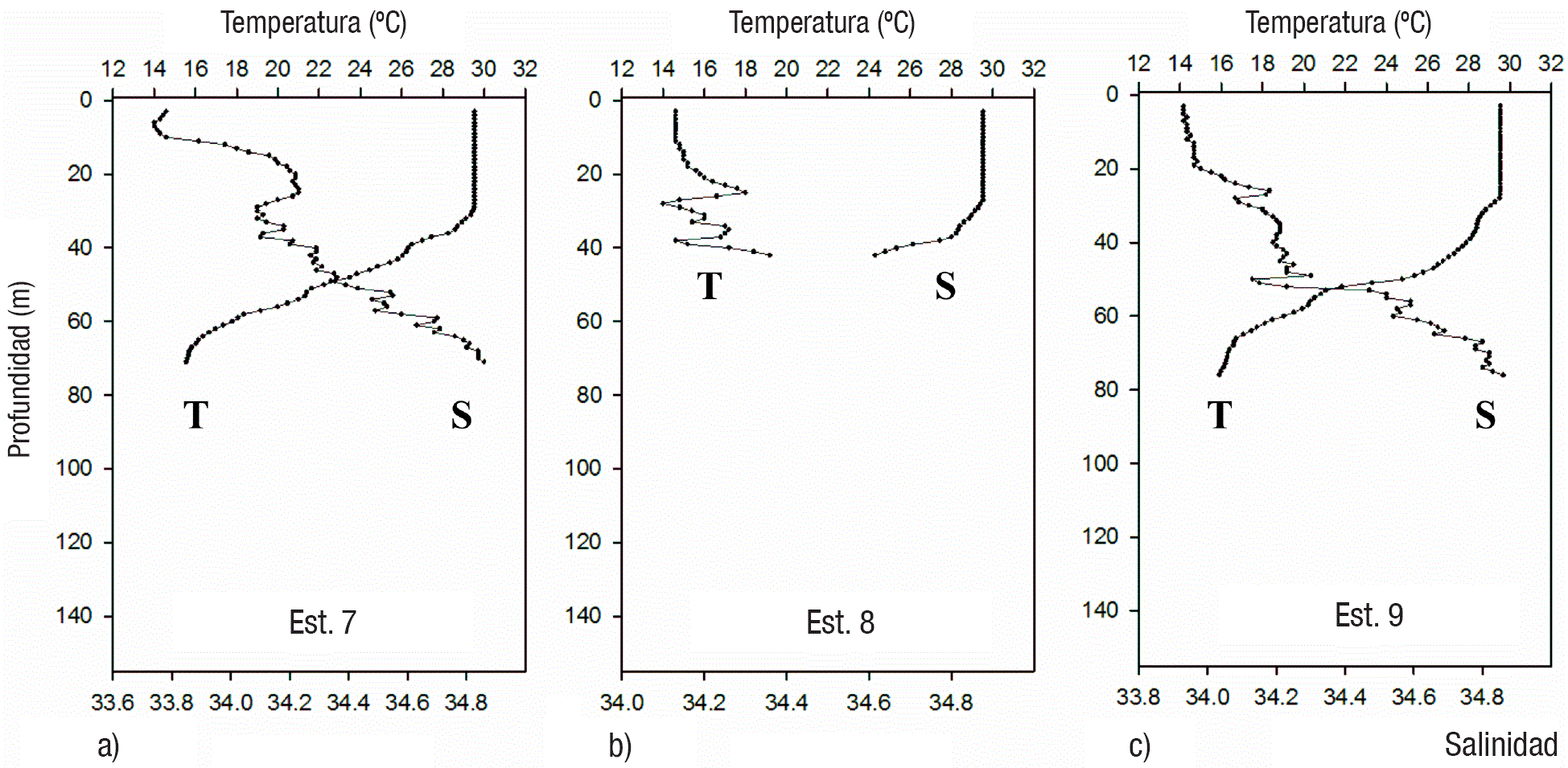
Figuras 4a-c Perfiles verticales de temperatura y salinidad en las estaciones de la zona Este (7-9) del Golfo de Tehuantepec.
Pigmentos fotosintéticos. Los valores de los pigmentos fotosintéticos en las estaciones del Golfo de Tehuantepec variaron considerablemente (hasta en cuatro órdenes de magnitud): Fucoxantina de 0.001 a 3.9 μg L-1 (1-3 900 ng L-1), Prasinoxantina de 0.002 a 0.9 μg L-1 (2-900 ng L-1), Violaxantina de 0.003 a 0.2 μg L-1 (3-200 ng L-1), Zeaxantina de 0.009 a 0.374 μg L-1 (9-374 ng L-1) y Divinil-clorofila a de 0.01 a 1.6 μg L-1 (1-1 600 ng L-1) (Tabla 2, Figs. 5, 6, 7). Todos los pigmentos estuvieron presentes en todas las muestras analizadas, aunque en algunos casos las concentraciones fueron muy bajas (Tabla 2, Figs. 5, 6, 7).
Tabla 2 Concentraciones de los pigmentos fotosintéticos a diferentes profundidades en las estaciones del Golfo de Tehuantepec. Todas las con centraciones están en μg L-1.
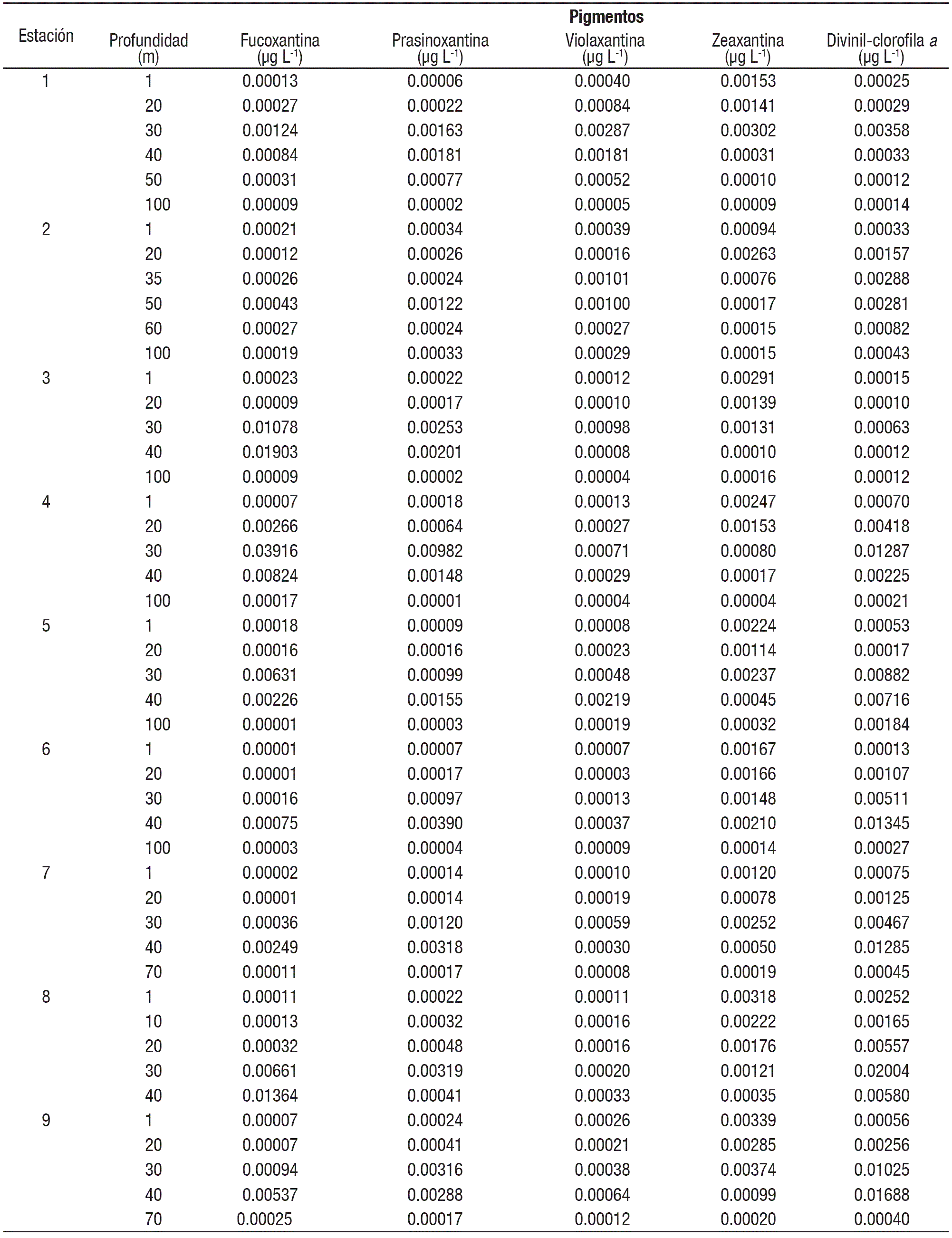
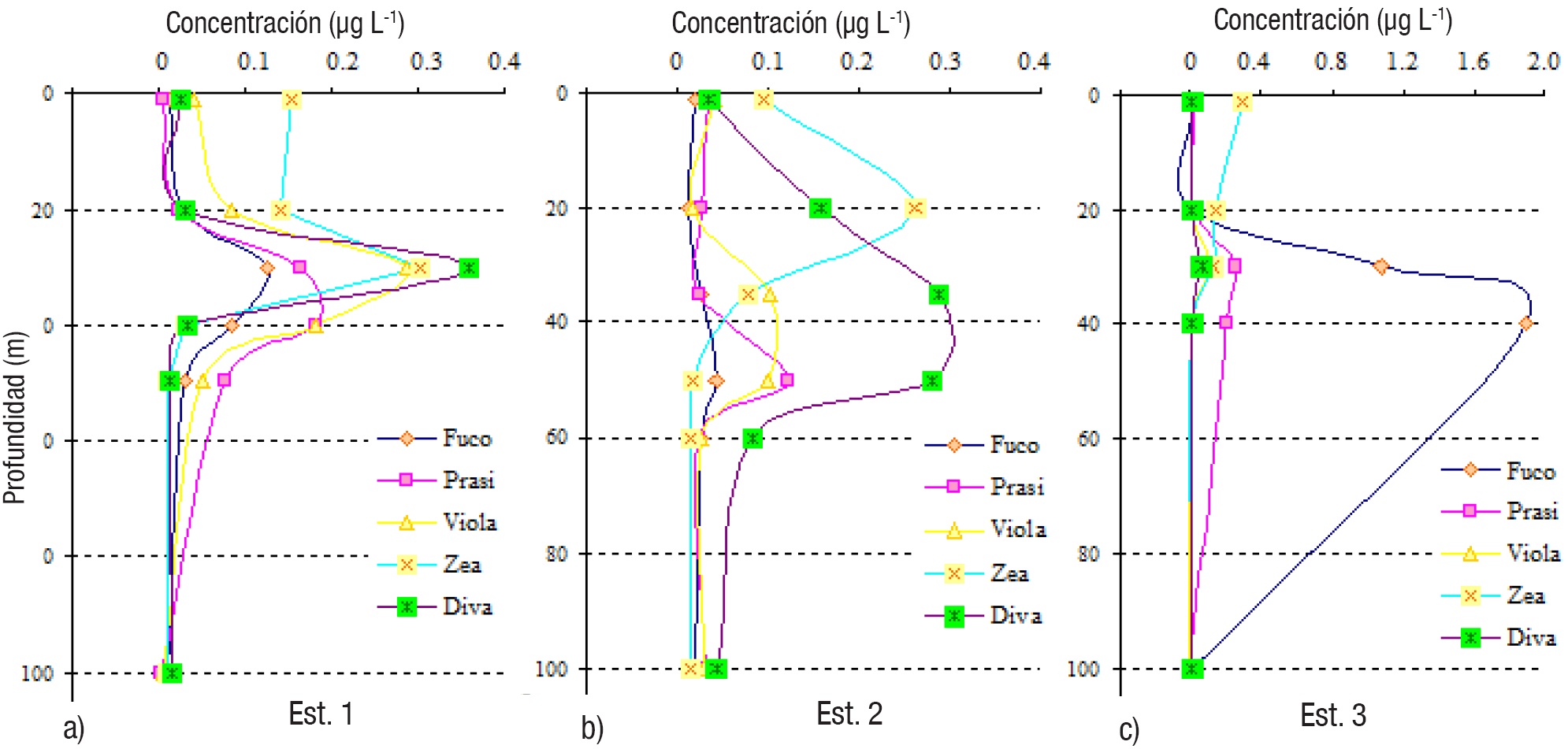
Figuras 5a-c Perfiles verticales de los pigmentos fotosintéticos del fitoplancton en las estaciones de la zona Oeste (1-3) del Golfo de Tehuantepec.
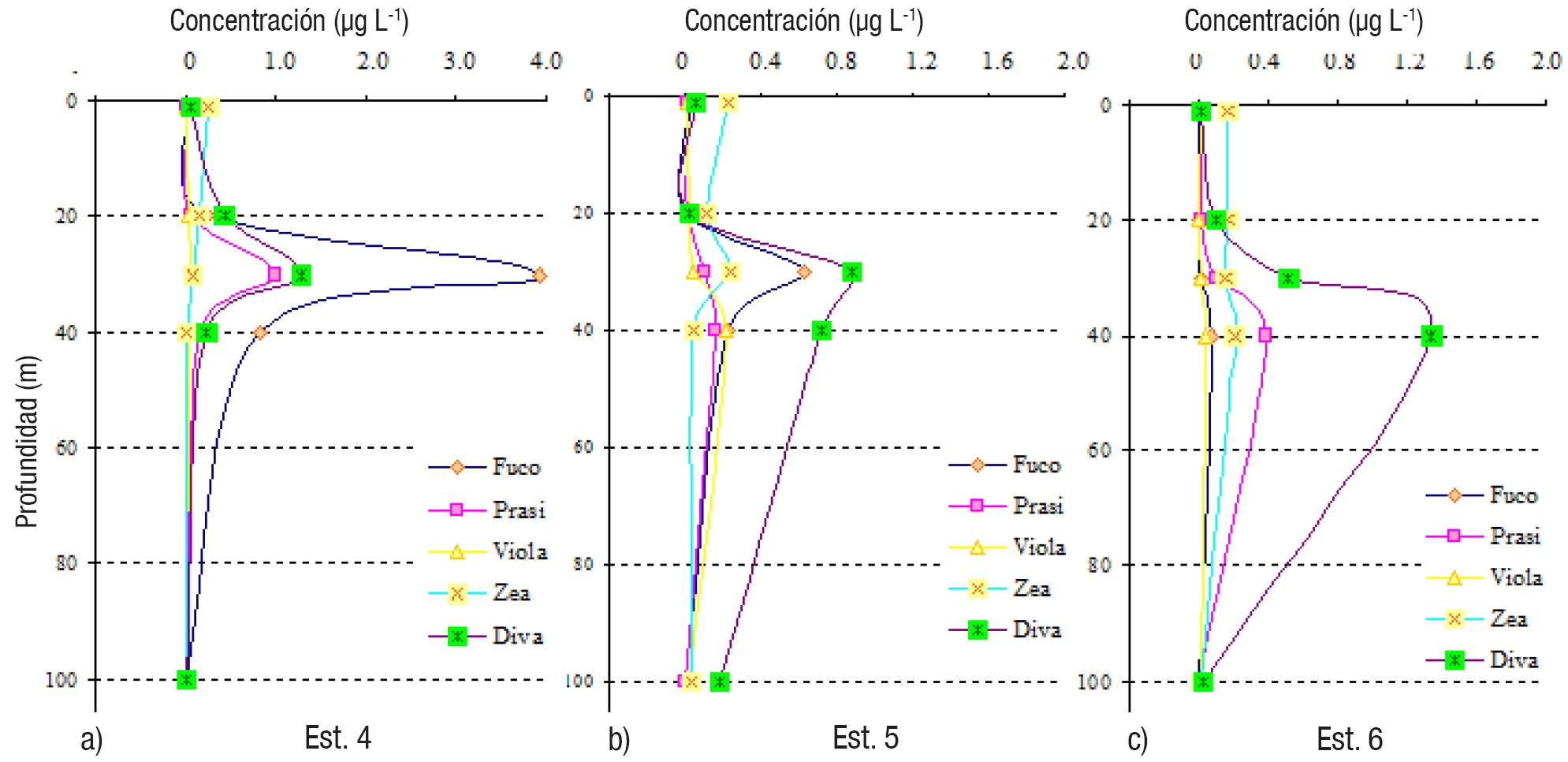
Figuras 6a-c Perfiles verticales de los pigmentos fotosintéticos del fitoplancton en las estaciones de la zona Centro (4-6) del Golfo de Tehuantepec.
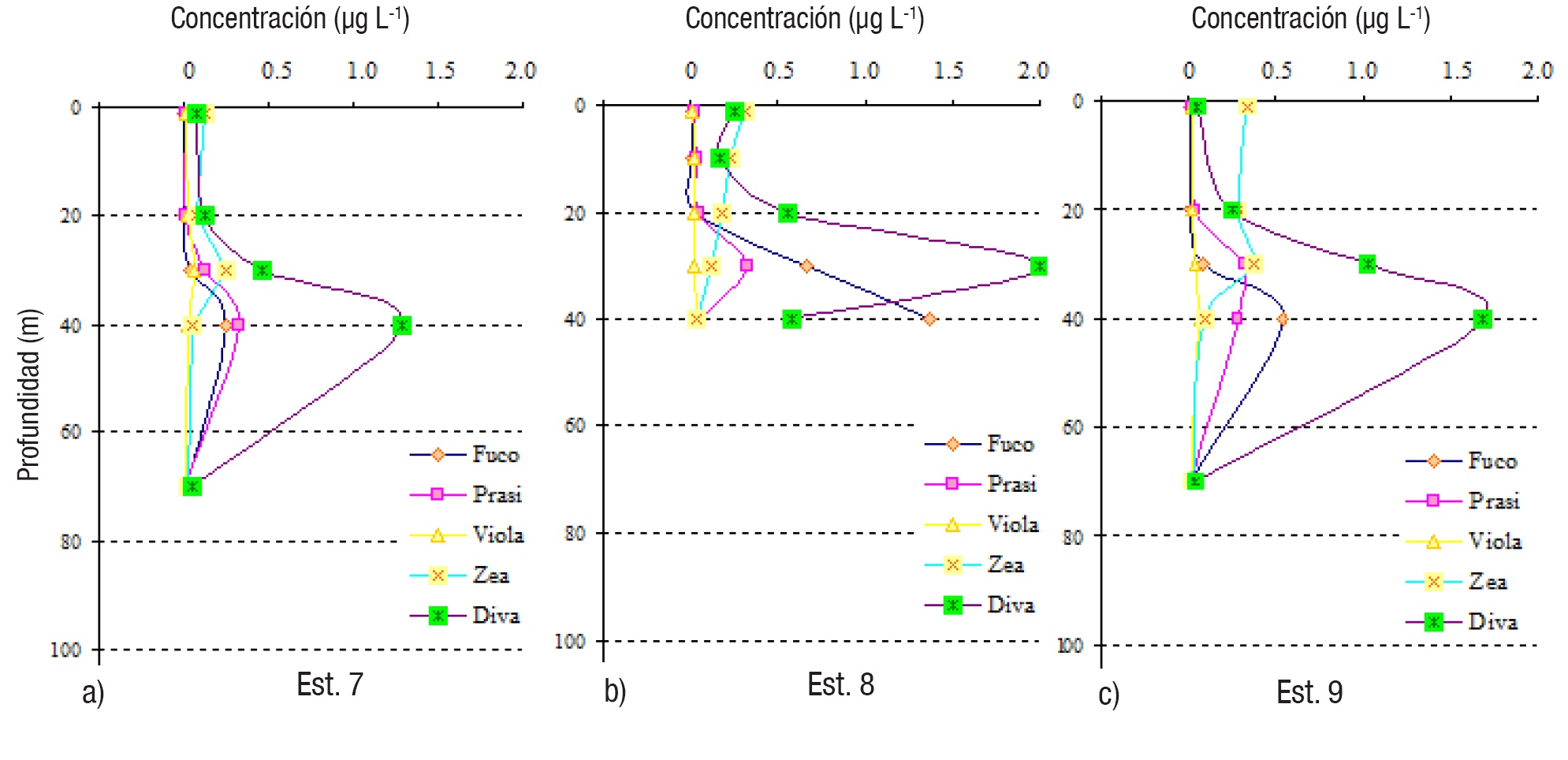
Figuras 7a-c Perfiles verticales de los pigmentos fotosintéticos del fitoplancton en las estaciones de la zona Este (7-9) del Golfo de Tehuantepec.
La mayoría de las estaciones mostraron predominancia de Divinilclorofila a, incluyendo estaciones poco profundas (7-9, Fig. 7) y más alejadas de la costa (1 y 7, Figs. 5, 7), donde sobresalen los picos subsuperfiales (entre 30 y 40 m) de dicho pigmento (Figs. 5, 6, 7); en la estación 8, a 30 m, se localizó la concentración más elevada de este pigmento (2.004 μg L-1) (Tabla 2). Las más altas concentraciones (con valores máximos mayores de 1.0 μg L-1) se detectaron en la zona Este (estaciones 7-9, menores de 100 m de profundidad) y la estación 6, del Centro (Tabla 2, Figs. 6-7).
En solo dos estaciones, 3 y 4 (Tabla 2, Figs. 5-6), la Fucoxantina fue el pigmento más representativo, notable principalmente en las picos máximos subsuperficiales (30 y 40 m), con una concentración máxima de 3.916 μg L-1 (estación 4, a 30 m) (Tabla 2, Fig. 6), indicando altas densidades de diatomeas. Las concentraciones de Zeaxantina fueron bajas en general, con un máximo de 0.374 μg L-1 en la estación 9, a 30 m (Tabla 2, Fig. 7).
La distribución vertical de todos los pigmentos tuvo un patrón general de picos de máximas concentraciones en subsuperficie (entre 30 y 40 m) (Tabla 2, Figs. 5, 6, 7); en la estación 2, la Zeaxantina tuvo un pico máximo más superficial (20 m) y la Prasinoxantina a 50 m (Tabla 2, Fig. 5), mientras que los otros pigmentos lo tuvieron entre 30 y 40 m (Figs. 5, 6, 7). A excepción de la estación 8, los patrones de distribución vertical de Fucoxantina y Divinil-clorofila a, fueron muy similares (Figs. 5, 6, 7). El pigmento Zeaxantina tuvo mayores concentraciones en superficie y subsuperficie en casi todas las estaciones (1-3, 5, 6-9) (Figs. 5, 6, 7), y disminuyó con la profundidad en las estaciones 3, 4 y 8 (Figs. 5, 7). No se notaron picos más profundos de Divinil-clorofila a, como en otras zonas oceánicas del Pacífico mexicano.
Discusión
Hidrografía y pigmentos fotosintéticos del fitoplancton. En las nueve estaciones de las tres zonas consideradas en este estudio (Oeste, Centro y Este) se observó una estratificación típica de verano, con relativa alta estabilidad de la columna de agua, y una capa superficial de espesor entre 20 y 40 m, que contiene agua cálida con temperatura cercana a 30° C y termoclinas principales bien identificados con su parte superior a esas profundidades (Tabla 1, Figs. 2, 3, 4). Los picos de pigmentos se localizaron debajo de la profundidad de cada termoclina, aunque hubo un pico de Zeaxantina a 20 m en la estación, 2, donde la termoclina está alrededor de los 33 m (Tabla 2, Figs. 2, 5), y el pico de Divinil-clorofila a coincidió con la profundidad de la termoclina en la estación 2 (Tabla 1); algunos picos de pigmentos (Divinil-clorofila a y Prasinoxantina) casi coincidieron con la profundidad de la termoclina en las estación 8 (Tablas 1 y 2, Figs. 4, 7).
Además del aumento de salinidad con la profundidad, el otro rasgo sobresaliente es el incremento súbito en la capa subsuperficial en las estaciones 2, 3 y 5, que puede explicarse como una intrusión de agua subsuperficial más salina, y no pareció afectar la estratificación térmica, pero se puede asociar con un pico de Zeaxantina (0.263 μg L-1) en la estación 2, a 20 m (Tabla 2, Figs. 2, 5).
Los valores encontrados de pigmentos fotosintéticos son comparables con los encontrados por Almazán-Becerril et al. (2010), para las costas occidentales de Baja California, donde aparecieron valores máximos de Fucoxantina de 0.95 μg L-1 y de Zeaxantina de 0.29 μg L-1, aunque en este trabajo la Fucoxantina llegó a ser cuatro veces más alta en su concentración máxima (3.9 μg L-1), una orden de magnitud más mayor que los detectados en Baja California; las concentraciones de Zeaxantina fueron muy similares a las de ese trabajo (un valor máximo de 0.374 μg L-1 en este estudio). Estos niveles también han sido encontrados en otras regiones del mundo, como ciertas áreas del Océano Atlántico (Barlow et al., 2002).
Dadas las elevadas concentraciones del pigmento Divinil-clorofila a (mayores a 1.0 μg L-1 en la zona Este del Golfo de Tehuantepec y otras estaciones, y un máximo de 1.6 μg L-1), fue evidente la predominancia de Prochlorococcus, el organismo fotosintético más pequeño (picoplanctonte) y abundante del planeta, y de amplia distribución mundial (Partensky et al., 1999; Casey et al., 2007), pero mejor representado en ambientes oceánicos, oligotróficos y tropicales o subtropicales (Campbell et al., 1997; Goericke et al., 2000; Veldhuis et al., 2005; Choi et al., 2011). La Fucoxantina, pigmento representativo de las diatomeas, fue considerable en dos estaciones (3 y 4), donde se mostraron picos máximos entre 30 y 40 m (Tabla 2, Figs. 5-6).
Es posible considerar que, aunque no se muestran datos de clorofila a, la distribución vertical de los pigmentos siga un patrón similar al de la clorofila a, que en otros trabajos ha mostrado picos máximos de concentración entre 10 y 18 m, para época de invierno (Robles-Jarero & Lara-Lara, 1993; Lara-Lara & Bazán-Guzmán, 2005). Estos valores máximos de clorofila a coincidieron con los picos de producción primaria medida (Robles-Jarero & Lara-Lara, 1993; Lara-Lara y Bazán-Guzmán, 2005).
La distribución vertical de los pigmentos mostró una estructura de capas ("layered structure", Barlow et al., 2002) en las estaciones 7 a 9, de la zona Este (Fig. 7), donde se localizarían distintas poblaciones de fitoplancton, sobre todo de formas picoplanctónicas, Prochlorococcus y Synechococcus, pero fue más notable esta condición en la estación 2 (Tablas 1 y 2, Fig. 5), con alternancia de picos de Zeaxantina (20 m) y Divinil-clorofila a (35 m). La distribución vertical de los pigmentos marcadores de Prochlorococcus y Synechococcus encontrada en este trabajo es similar a la mostrada en aguas oligotróficas tropicales por Veldhuis et al. (2005, Figs. 1, 5).
Condiciones ambientales y comunidades fitoplanctónicas. Se puede esperar que varíen las contribuciones de las diferentes fracciones de talla del fitoplancton a la biomasa total y productividad primaria, en distintas épocas climáticas (Lara-Lara y Bazán-Guzmán, 2005), por lo que se puede plantear la posibilidad de encontrar distintas estaciones climáticas o fases temporales del Golfo de Tehuantepec, sobre la base de condiciones oceanográficas e hidrográficas (e.g. Machain-Castillo et al., 2008) y los pigmentos fotosintéticos (que indican la predominancia de diferentes grupos fitoplanctónicos). Algunas de estas fases podrían corresponder solo a pulsos temporales de poca duración (días o semanas), moduladas sobre todo por la intensidad de vientos y profundidad de la termoclina (o capa de mezcla), o de más largo plazo por la influencia de tormentas tropicales (Lluch-Cota et al., 1997).
Una fase climática es en invierno y principio de primavera, con valores de nutrientes (nitratos) relativamente altos (mayores de 5 μM) (Machain-Castillo et al., 2008), y donde ya son conocidas las elevadas densidades de diatomeas, usualmente productoras de cadenas o colonias, o con formas solitarias de tallas mayores a 20 μm, que forman parte del microplancton y que predominan en condiciones de mezcla turbulenta y surgencias, que son aparentes en invierno y principio de primavera (Meave & Hernández-Becerril 1998), precisamente cuando soplan los vientos del norte sobre la parte central del Golfo de Tehuantepec.
Sin embargo también existe información que difiere un poco, ya que en invierno (febrero) Lara-Lara y Bazán-Guzmán (2005) destacaron la contribución del nanoplancton a la biomasa (como clorofila a) y a la productividad primaria, con entre 60 y 100%, y 70 y 100%, respectivamente, y la del picoplancton con un 50% del total de la clorofila a, con una participación importante del microplancton en la porción oriental del Golfo de Tehuantepec, y en primavera (abril, 2000) Bravo-Sierra y Hernández-Becerril (2003) encontraron poblaciones densas de especies nanoplanctónicas (diatomeas pequeñas, cocolitofóridos y Parmales) en varios puntos del Golfo de Tehuantepec.
Otra fase climática es el verano (junio), cuando la concentración de clorofila a superficial es más baja (Lluch-Belda et al., 1997), los valores de nitratos son relativamente bajos (menores a 1 μM) (Machain-Castillo et al., 2008) y hay ausencia de surgencias. Además en este trabajo mostramos valores altos de temperatura superficial y la columna de agua bien estratificada (Figs. 2, 3, 4). En este escenario, la fracción del picoplancton, representada por los pigmentos diagnósticos Divinil-clorofila a para Prochlorococcus y Zeaxantina para Synechococcus, parece predominante (a excepción de un par de estaciones, 3 que es costera y 4, un poco más oceánica), tanto en superficie (Zeaxantina), como en los picos máximos de subsuperficie (Divinil-clorofila a) (Figs. 5, 6, 7). El picoplancton aparece también como un grupo funcional distinto, sobre todo por su talla y capacidad de adaptación a condiciones bajas de luz (Anabalón et al., 2014).
Han sido encontradas diferentes estructuras de fitoplancton en varias regiones del mundo, donde los eventos de surgencia fuertes son sucedidos por surgencias más débiles o franca relajación, lo que se refleja en la composición, abundancia, fracciones de talla y grupos funcionales (autótrofos y heterótrofos) del fitoplancton (Anabalón et al., 2014).
Se espera que para estudios futuros, se pueda agregar a la información de pigmentos fotosintéticos, aquella derivada de la estructura de la comunidad (composición y abundancia), sobre todo con el empleo de citometría de flujo para análisis de picoplancton y fluorescencia natural in situ, además de complementar con estudios de biología molecular. La descripción de las comunidades debería conducir a estudios detallados de los mecanismos de producción y su exportación a los niveles tróficos secundarios.

